
Applications of Bacillus subtilis Protein Display for Medicine, Catalysis, Environmental Remediation, and Protein Engineering

Abstract
:1. Introduction
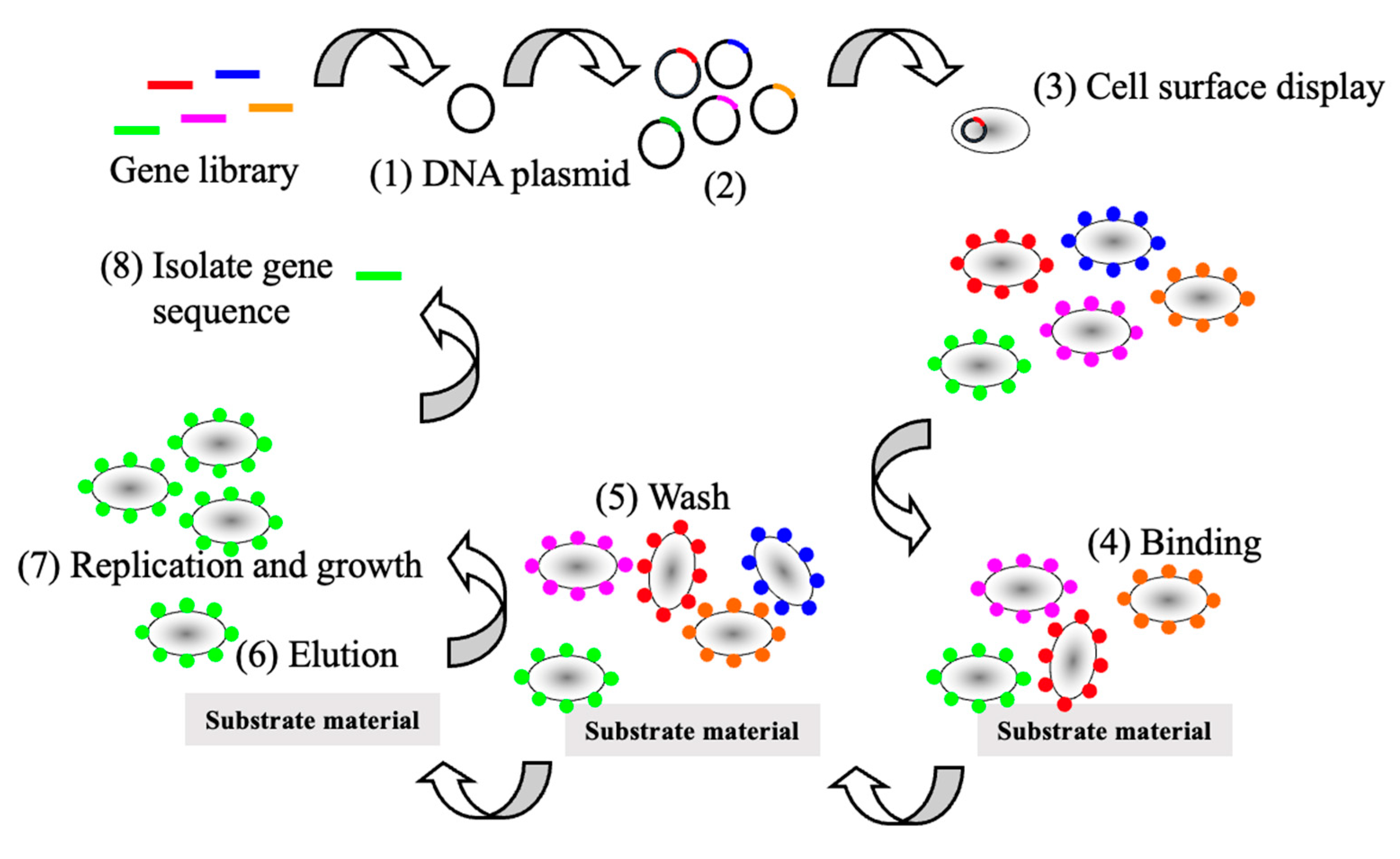
1.1. Protein Display
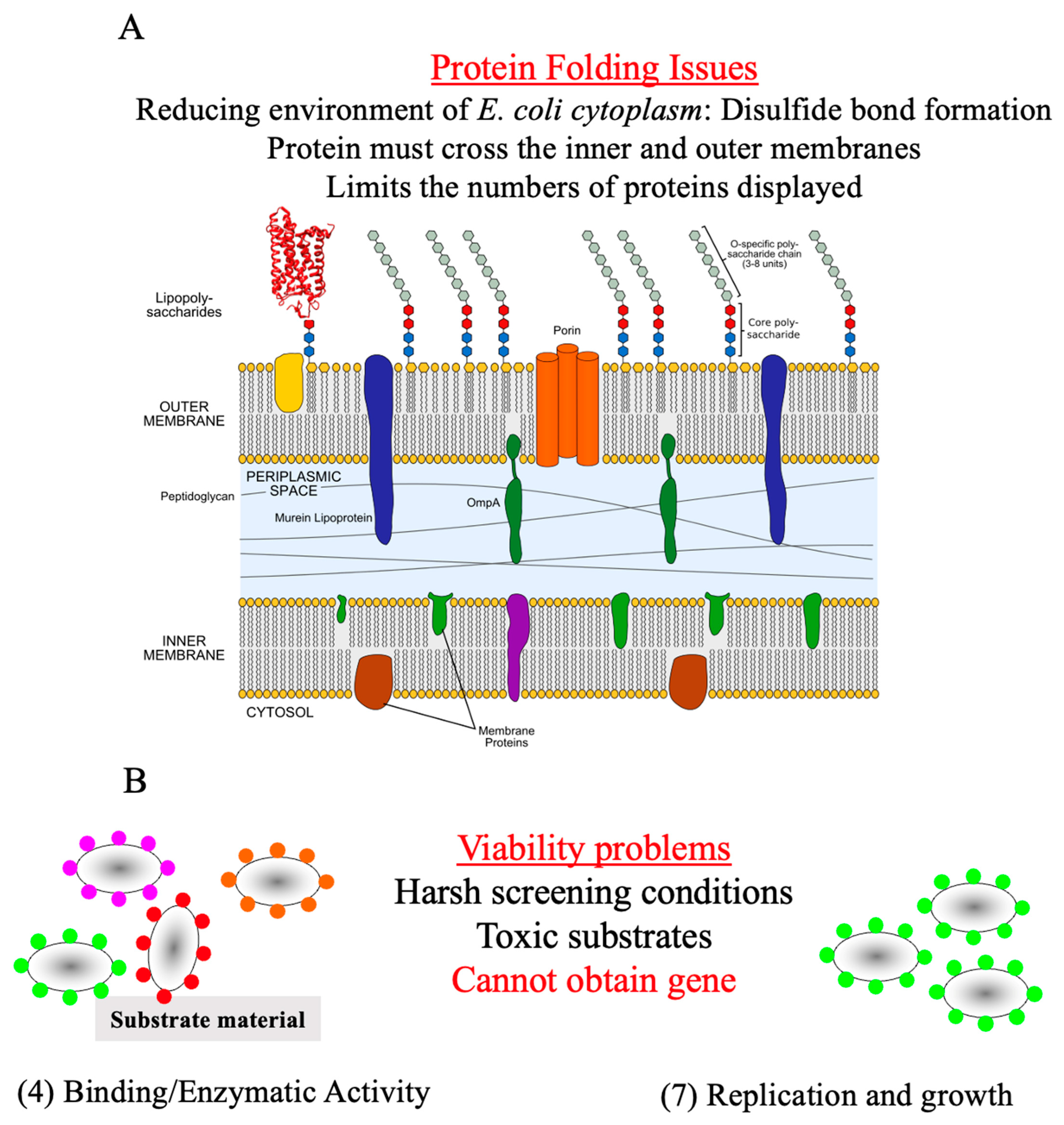
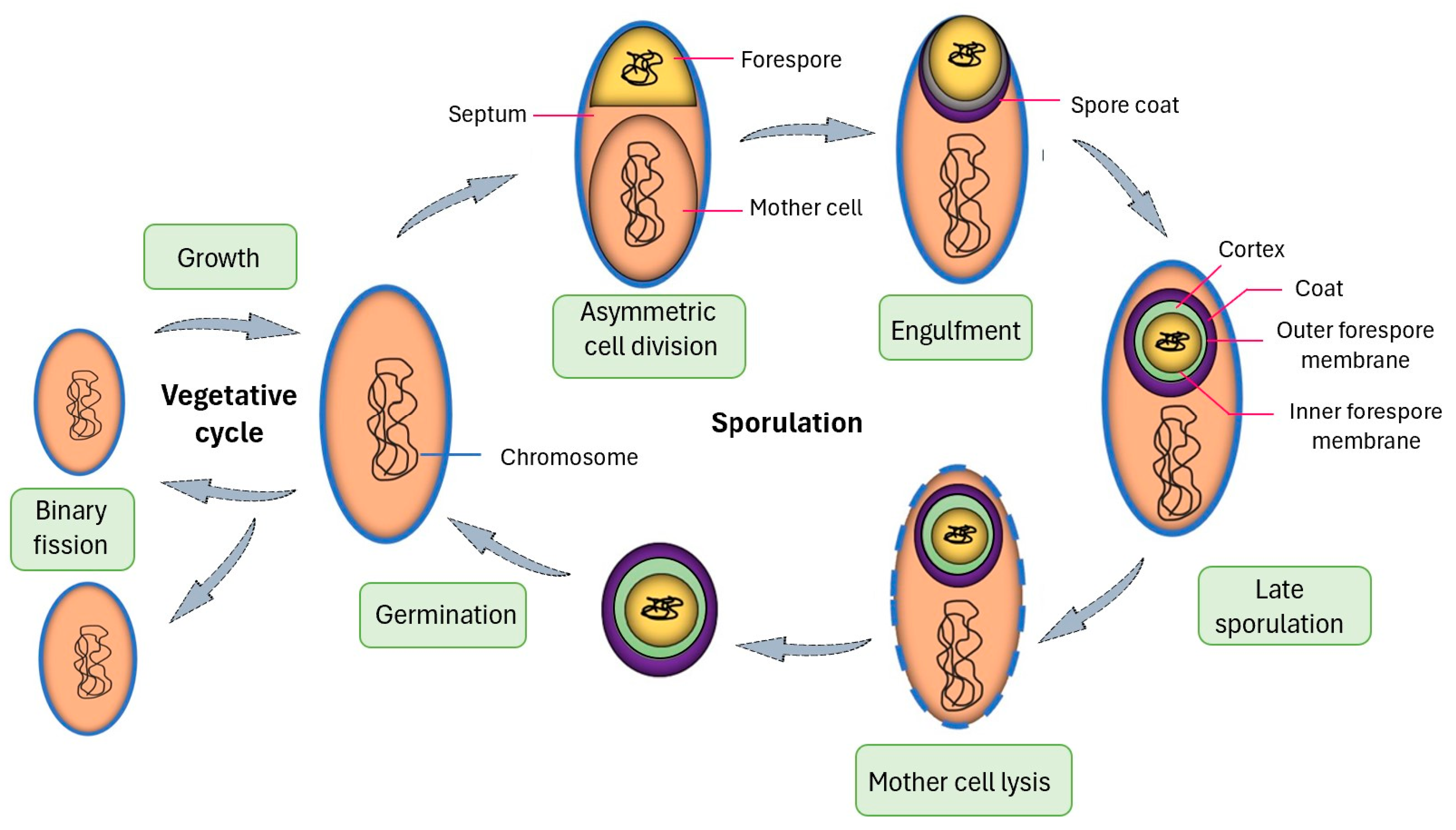
1.2. Advantages of Spore Display
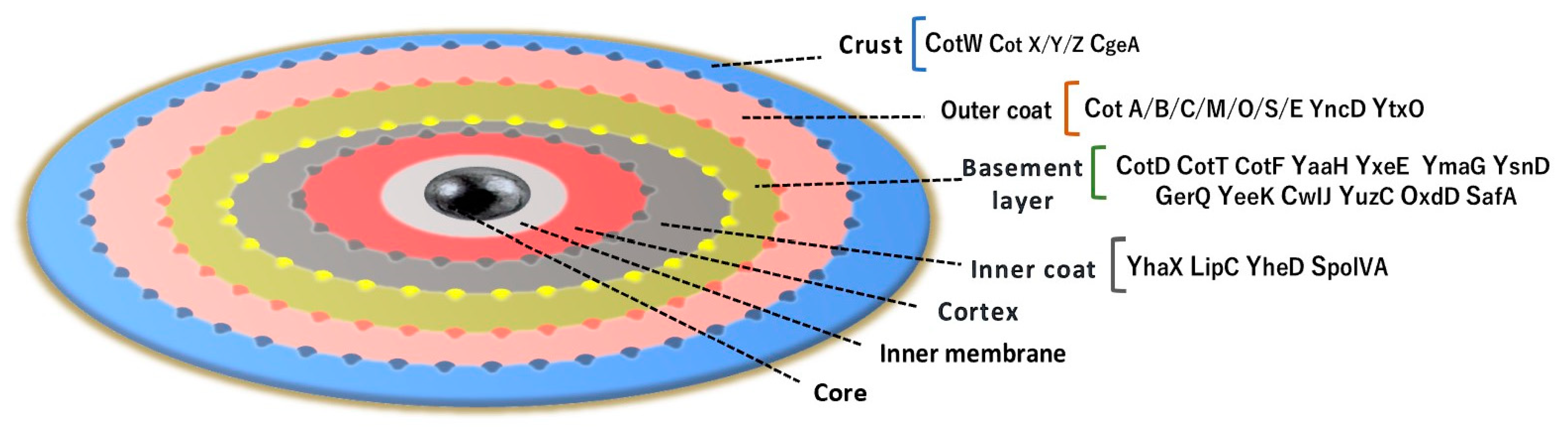
1.3. B. subtilis Anchor Proteins for Protein Display
2. Applications
2.1. Vaccine Development/Drug Delivery
2.2. Biocatalysis
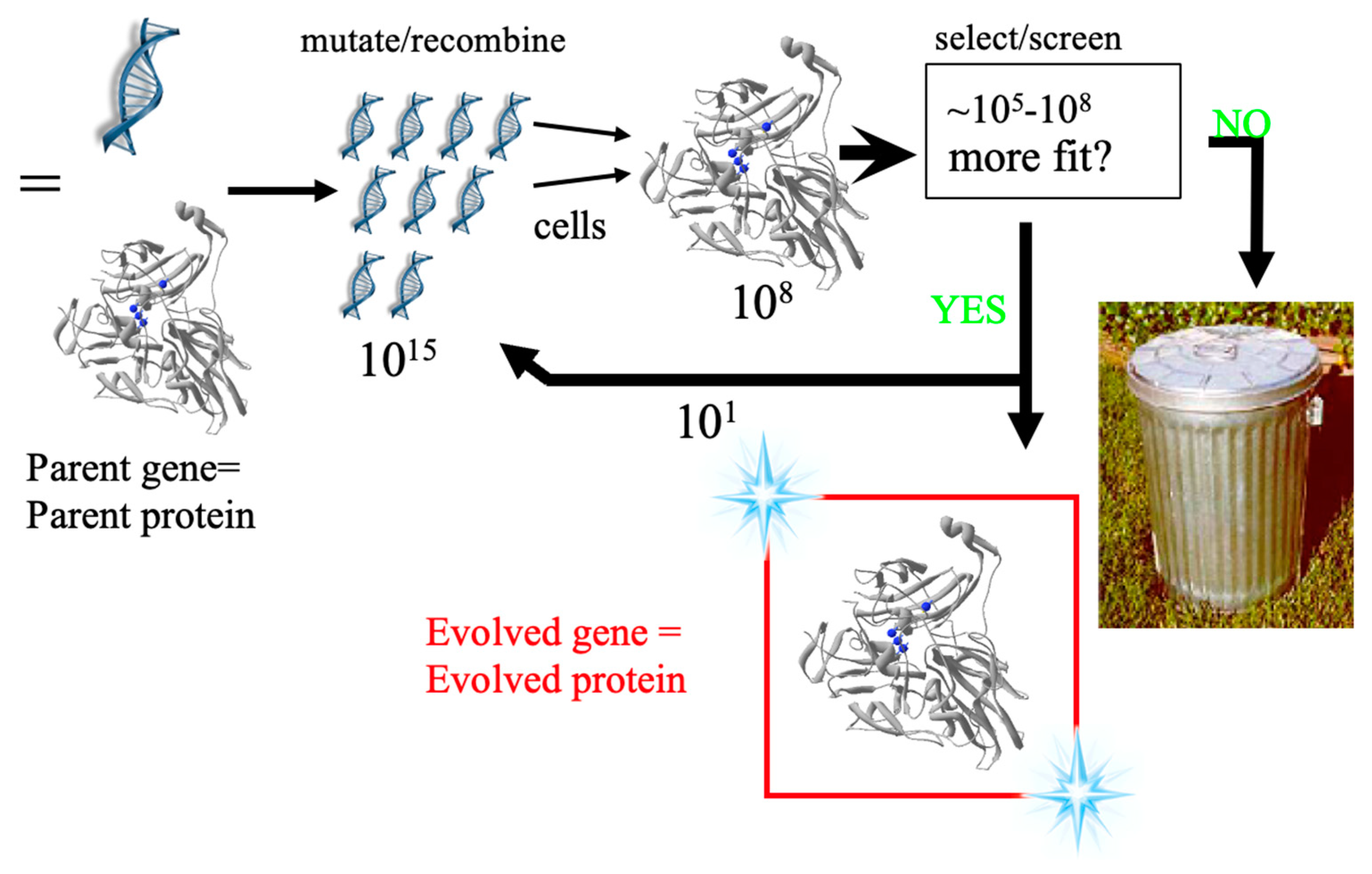
2.3. Protein Engineering and Optimization
2.4. Environmental Applications
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Arnold, F.H. Advances in Protein Chemistry. Evolutionary Protein Design; Academic Press: New York, NY, USA, 2001; Volume 55, p. 438. [Google Scholar]
- Amstutz, P.; Forrer, P.; Zahnd, C.; Pluckthun, A. In vitro display technologies: Novel developments and applications. Curr. Opin. Biotechnol. 2001, 12, 400–405. [Google Scholar] [CrossRef]
- Forrer, P.; Jung, S.; Pluckthun, A. Beyond binding: Using phage display to select for structure, folding and enzymatic activity in proteins. Curr. Opin. Struct. Biol. 1999, 9, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.; Schmoldt, H.U.; Adams, T.M.; Wilhelm, S.; Kolmar, H. Ultra-high-throughput screening based on cell-surface display and fluorescence-activated cell sorting for the identification of novel biocatalysts. Curr. Opin. Biotechnol. 2004, 15, 323–329. [Google Scholar] [CrossRef]
- Lee, S.Y.; Choi, J.H.; Xu, Z. Microbial cell-surface display. Trends Biotechnol. 2003, 21, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Wittrup, K.D. Protein engineering by cell-surface display. Curr. Opin. Biotechnol. 2001, 12, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Bessette, P.H.; Aslund, F.; Beckwith, J.; Georgiou, G. Efficient folding of proteins with multiple disulfide bonds in the Escherichia coli cytoplasm. Proc. Natl. Acad. Sci. USA 1999, 96, 13703–13708. [Google Scholar] [CrossRef]
- Driks, A. Bacillus subtilis spore coat. Microbiol. Mol. Biol. Rev. 1999, 63, 1–20. [Google Scholar] [CrossRef]
- Stragier, P.; Losick, R. Molecular genetics of sporulation in Bacillus subtilis. Annu. Rev. Genet. 1996, 30, 297-41. [Google Scholar] [CrossRef]
- Nicholson, W.L.; Munakata, N.; Horneck, G.; Melosh, H.J.; Setlow, P. Resistance of Bacillus endospores to extreme terrestrial and extraterrestrial environments. Microbiol. Mol. Biol. Rev. 2000, 64, 548–572. [Google Scholar] [CrossRef]
- Kennedy, M.J.; Reader, S.L.; Swierczynski, L.M. Preservation records of micro-organisms: Evidence of the tenacity of life. Microbiology (Reading) 1994, 140 Pt 10, 2513–2529. [Google Scholar] [CrossRef]
- Fessner, W.-D. (Ed.) Biocatalysis: From Discovery to Applications; Topics in Current Chemistry; Springer-Verlag: Berlin/Heidelberg, Germany, 2000; Volume 200, p. 254. [Google Scholar]
- Isticato, R.; Cangiano, G.; Tran, H.T.; Ciabattini, A.; Medaglini, D.; Oggioni, M.R.; De Felice, M.; Pozzi, G.; Ricca, E. Surface display of recombinant proteins on Bacillus subtilis spores. J. Bacteriol. 2001, 183, 6294–6301. [Google Scholar] [CrossRef] [PubMed]
- Isticato, R. Bacterial Spore-Based Delivery System: 20 Years of a Versatile Approach for Innovative Vaccines. Biomolecules 2023, 13, 947. [Google Scholar] [CrossRef] [PubMed]
- Duc, L.H.; Hong, H.A.; Fairweather, N.; Ricca, E.; Cutting, S.M. Bacterial spores as vaccine vehicles. Infect. Immun. 2003, 71, 2810–2818. [Google Scholar] [CrossRef] [PubMed]
- Vetráková, A.; Chovanová, R.K.; Rechtoríková, R.; Krajčíková, D.; Barák, I. Bacillus subtilis spores displaying RBD domain of SARS-CoV-2 spike protein. Comput. Struct. Biotechnol. J. 2023, 21, 1550–1556. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.C.L.; Li, P.; Tsang, M.S.M.; Sung, J.C.C.; Kwong, K.W.Y.; Zheng, T.; Hon, S.S.M.; Lau, C.P.; Cheng, W.; Chen, F.; et al. Creating a Vaccine-like Supplement against Respiratory Infection Using Recombinant Bacillus subtilis Spores Expressing SARS-CoV-2 Spike Protein with Natural Products. Molecules 2023, 28, 4996. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.S.; Guluarte, J.O.; Abbott, D.W.; Inglis, G.D.; Guan, L.L.; Alexander, T.W. Development of a spore-based mucosal vaccine against the bovine respiratory pathogen Mannheimia haemolytica. Sci. Rep. 2023, 13, 12981. [Google Scholar] [CrossRef]
- Yin, L.; Meng, Z.; Zhang, Y.; Hu, K.; Chen, W.; Han, K.; Wu, B.Y.; You, R.; Li, C.H.; Jin, Y.; et al. Bacillus spore-based oral carriers loading curcumin for the therapy of colon cancer. J. Control. Release 2018, 271, 31–44. [Google Scholar] [CrossRef]
- Wang, H.; Yang, R.; Hua, X.; Zhao, W.; Zhang, W. Functional display of active β-galactosidase on Bacillus subtilis spores using crust proteins as carriers. Food Sci. Biotechnol. 2015, 24, 1755–1759. [Google Scholar] [CrossRef]
- Seok, J.K.; Jung, H.C.; Pan, J.G. Transgalactosylation in a water-solvent biphasic reaction system with β-galactosidase displayed on the surfaces of Bacillus subtilis spores. Appl. Environ. Microbiol. 2007, 73, 2251–2256. [Google Scholar]
- Tavassoli, S.; Hinc, K.; Iwanicki, A.; Obuchowski, M.; Ahmadian, G. Investigation of spore coat display of Bacillus subtilis β-galactosidase for developing of whole cell biocatalyst. Arch. Microbiol. 2013, 195, 197–202. [Google Scholar] [CrossRef]
- He, W.; Jiang, B.; Mu, W.; Zhang, T. Production of d-Allulose with d-Psicose 3-Epimerase Expressed and Displayed on the Surface of Bacillus subtilis Spores. J. Agric. Food Chem. 2016, 64, 7201–7207. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Tian, R.; Ni, Z.; Zhang, Q.; Zhang, T.; Chen, Z.; Chen, K.; Yang, S. Surface display of the thermophilic lipase Tm1350 on the spore of Bacillus subtilis by the CotB anchor protein. Extremophiles 2015, 19, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, T.; Jia, J.; Vastermark, A.; Tian, R.; Ni, Z.; Chen, Z.; Chen, K.; Yang, S. Expression and display of a novel thermostable esterase from Clostridium thermocellum on the surface of Bacillus subtilis using the CotB anchor protein. J. Ind. Microbiol. Biotechnol. 2015, 42, 1439–1448. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Feng, F.; Chen, L.; Yao, Q.; Chen, K. Surface display of Acetobacter pasteurianus AdhA on Bacillus subtilis spores to enhance ethanol tolerance for liquor industrial potential. Eur. Food Res. Technol. 2014, 238, 285–293. [Google Scholar] [CrossRef]
- Gao, C.; Xu, X.; Zhang, X.; Che, B.; Ma, C.; Qiu, J.; Tao, F.; Xu, P. Chemoenzymatic synthesis of N-acetyl-D-neuraminic acid from N-acetyl-D-glucosamine by using the spore surface-displayed N-acetyl-D-neuraminic acid aldolase. Appl. Environ. Microbiol. 2011, 77, 7080–7083. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, Z.; Ni, Z.; Tian, R.; Zhang, T.; Jia, J.; Chen, K.; Yang, S. Display of Thermotoga maritima MSB8 nitrilase on the spore surface of Bacillus subtilis using out coat protein CotG as the fusion partner. J. Mol. Catal. B Enzym. 2016, 123, 73–80. [Google Scholar] [CrossRef]
- Gupta, N.; Lee, F.S.; Farinas, E.T. Laboratory evolution of laccase for substrate specificity. J. Mol. Catal. B Enzym. 2010, 62, 230–234. [Google Scholar] [CrossRef]
- Gupta, N.; Farinas, E.T. Directed evolution of CotA laccase for increased substrate specificity using Bacillus subtilis spores. Protein Eng. Des. Sel. 2010, 23, 679–682. [Google Scholar] [CrossRef]
- Gupta, N.; Farinas, E.T. Narrowing laccase substrate specificity using active site saturation mutagenesis. Comb. Chem. High Throughput Screen. 2009, 12, 269–274. [Google Scholar] [CrossRef]
- Jia, H.; Lee, F.S.; Farinas, E.T. Bacillus subtilis spore display of laccase for evolution under extreme conditions of high concentrations of organic solvent. ACS Combi. Sci. 2014, 16, 665–669. [Google Scholar] [CrossRef]
- Sheng, S.; Jia, H.; Topiol, S.; Farinas, E.T. Engineering CotA Laccase for Acidic pH Stability Using Bacillus subtilis Spore Display. J. Microbiol. Biotechnol. 2017, 27, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Sheng, S.; Farinas, E.T. Laccase and Its Mutant Displayed on the Bacillus subtilis Spore Coat for Oxidation of Phenolic Compounds in Organic Solvents. Catalysts 2021, 11, 606. [Google Scholar] [CrossRef]
- Hosseini-Abari, A.; Kim, B.G.; Lee, S.H.; Emtiazi, G.; Kim, W.; Kim, J.H. Surface display of bacterial tyrosinase on spores of Bacillus subtilis using CotE as an anchor protein. J. Basic. Microbiol. 2016, 56, 1331–1337. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Song, T.; Jiang, H.; Pei, C.; Huang, Q.; Xi, H. Bacillus subtilis Spore Surface Display of Haloalkane Dehalogenase DhaA. Curr. Microbiol. 2019, 76, 1161–1167. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Wang, J.; Zhang, Z.; Shi, S.; Li, D.; Shen, W.; Shen, E.; Zhou, J. Catalytic transformation of HODAs using an efficient meta-cleavage product hydrolase-spore surface display system. J. Mol. Catal. B Enzym. 2014, 102, 204–210. [Google Scholar] [CrossRef]
- Hinc, K.; Ghandili, S.; Karbalaee, G.; Shali, A.; Noghabi, K.A.; Ricca, E.; Ahmadian, G. Efficient binding of nickel ions to recombinant Bacillus subtilis spores. Res. Microbiol. 2010, 161, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Li, S.; Camilleri, E.; Korza, G.; Yankova, M.; King, S.M.; Setlow, P. Accumulation and release of rare earth ions by spores of Bacillus species and the location of these ions in spores. Appl. Environ. Microbiol. 2019, 85, e00956-19. [Google Scholar] [CrossRef]
- Valenzuela-García, L.I.; Alarcón-Herrera, M.T.; Ayala-García, V.M.; Barraza-Salas, M.; Salas-Pacheco, J.M.; Díaz-Valles, J.F.; Pedraza-Reyes, M. Design of a Whole-Cell Biosensor Based on Bacillus subtilis Spores and the Green Fluorescent Protein to Monitor Arsenic. Microbiol. Spectr. 2023, 11, e00432-23. [Google Scholar] [CrossRef]
- McKenney, P.T.; Driks, A.; Eichenberger, P. The Bacillus subtilis endospore: Assembly and functions of the multilayered coat. Nat. Rev. Microbiol. 2013, 11, 33–44. [Google Scholar] [CrossRef]
- Kunst, F.; Ogasawara, N.; Moszer, I.; Albertini, A.M.; Alloni, G.; Azevedo, V.; Bertero, M.G.; Bessieres, P.; Bolotin, A.; Borchert, S.; et al. The complete genome sequence of the gram-positive bacterium Bacillus subtilis. Nature 1997, 390, 249–256. [Google Scholar] [CrossRef]
- Moszer, I. The complete genome of Bacillus subtilis: From sequence annotation to data management and analysis. FEBS Lett. 1998, 430, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Cutting, S.M. Bacillus probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.A.; Huang, J.M.; Khaneja, R.; Hiep, L.V.; Urdaci, M.C.; Cutting, S.M. The safety of Bacillus subtilis and Bacillus indicus as food probiotics. J. Appl. Microbiol. 2008, 105, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Wan, Q.; Krajcikova, D.; Tang, J.; Tzokov, S.B.; Barak, I.; Bullough, P.A. Diverse supramolecular structures formed by self-assembling proteins of the Bacillus subtilis spore coat. Mol. Microbiol. 2015, 97, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Bartels, J.; Lopez Castellanos, S.; Radeck, J.; Mascher, T. Sporobeads: The Utilization of the Bacillus subtilis Endospore Crust as a Protein Display Platform. ACS Synth. Biol. 2018, 7, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Zilhao, R.; Serrano, M.; Isticato, R.; Ricca, E.; Moran, C.P., Jr.; Henriques, A.O. Interactions among CotB, CotG, and CotH during assembly of the Bacillus subtilis spore coat. J. Bacteriol. 2004, 186, 1110–1119. [Google Scholar] [CrossRef] [PubMed]
- Isticato, R.; Esposito, G.; Zilhao, R.; Nolasco, S.; Cangiano, G.; De Felice, M.; Henriques, A.O.; Ricca, E. Assembly of multiple CotC forms into the Bacillus subtilis spore coat. J. Bacteriol. 2004, 186, 1129–1135. [Google Scholar] [CrossRef]
- Donovan, W.; Zheng, L.; Sandman, K.; Losick, R. Genes encoding spore coat polypeptides from Bacillus subtilis. J. Mol. Biol. 1987, 196, 1–10. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, C.S.; Kim, B.G. Spore-displayed streptavidin: A live diagnostic tool in biotechnology. Biochem. Biophys. Res. Commun. 2005, 331, 210–214. [Google Scholar] [CrossRef]
- Sacco, M.; Ricca, E.; Losick, R.; Cutting, S. An additional GerE-controlled gene encoding an abundant spore coat protein from Bacillus subtilis. J. Bacteriol. 1995, 177, 372–377. [Google Scholar] [CrossRef]
- Kim, J.; Schumann, W. Display of proteins on Bacillus subtilis endospores. Cell. Mol. Life Sci. 2009, 66, 3127–3136. [Google Scholar] [CrossRef] [PubMed]
- Ciabattini, A.; Parigi, R.; Isticato, R.; Oggioni, M.R.; Pozzi, G. Oral priming of mice by recombinant spores of Bacillus subtilis. Vaccine 2004, 22, 4139–4143. [Google Scholar] [CrossRef] [PubMed]
- Mauriello, E.M.; Duc, L.H.; Isticato, R.; Cangiano, G.; Hong, H.A.; De Felice, M.; Ricca, E.; Cutting, S.M. Display of heterologous antigens on the Bacillus subtilis spore coat using CotC as a fusion partner. Vaccine 2004, 22, 1177–1187. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.D.; Fulton, R.W.; Lehenbauer, T.W.; Step, D.L.; Confer, A.W. The epidemiology of bovine respiratory disease: What is the evidence for predisposing factors? Can. Vet. J. 2010, 51, 1095–1102. [Google Scholar] [PubMed]
- Li, W.; Li, J.; Dai, X.; Liu, M.; Khalique, A.; Wang, Z.; Zeng, Y.; Zhang, D.; Ni, X.; Zeng, D.; et al. Surface Display of porcine circovirus type 2 antigen protein cap on the spores of Bacillus subtilis 168: An effective mucosal vaccine candidate. Front. Immunol. 2022, 13, 1007202. [Google Scholar] [CrossRef]
- Lê, K.A.; Robin, F.; Roger, O. Sugar replacers: From technological challenges to consequences on health. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 310–315. [Google Scholar] [CrossRef]
- Hossain, A.; Yamaguchi, F.; Matsuo, T.; Tsukamoto, I.; Toyoda, Y.; Ogawa, M.; Nagata, Y.; Tokuda, M. Rare sugar d-allulose: Potential role and therapeutic monitoring in maintaining obesity and type 2 diabetes mellitus. Pharmacol. Ther. 2015, 155, 49–59. [Google Scholar] [CrossRef]
- Gaonkar, S.K.; Alvares, J.J.; Furtado, I.J. Recent advances in the production, properties and applications of haloextremozymes protease and lipase from haloarchaea. World J. Microbiol. Biotechnol. 2023, 39, 322. [Google Scholar] [CrossRef]
- Peng, Y.; Su, A.; Huang, W.; Lan, S.; Yang, T.; Tan, Q. Research progress on microbial thermophilic lipase. Food. Ferment. Ind. 2021, 47, 289–294. [Google Scholar]
- Maldonado, R.R.; Lopes, D.B.; Aguiar-Oliveira, E.; Kamimura, E.S.; Macedo, G.A. A review on Geotrichum lipases: Production, purification, immobilization and applications. Chem. Biochem. Eng. Q. 2016, 30, 439–454. [Google Scholar] [CrossRef]
- Brígida, A.I.S.; Amaral, P.F.F.; Coelho, M.A.Z.; Gonçalves, L.R.B. Lipase from Yarrowia lipolytica: Production, characterization and application as an industrial biocatalyst. J. Mol. Catal. B Enzym. 2014, 101, 148–158. [Google Scholar] [CrossRef]
- Pace, H.C.; Brenner, C. The nitrilase superfamily: Classification, structure and function. Genome Biol. 2001, 2, reviews0001.1–reviews0001.9. [Google Scholar] [CrossRef] [PubMed]
- Solomon, E.I.; Sundaram, U.M.; Machonkin, T.E. Multicopper oxidases and oxygenases. Chem. Rev. 1996, 96, 2563–2605. [Google Scholar] [CrossRef] [PubMed]
- Mayolo-Deloisa, K.; González-González, M.; Rito-Palomares, M. Laccases in Food Industry: Bioprocessing, Potential Industrial and Biotechnological Applications. Front. Bioeng. Biotechnol. 2020, 8, 00222. [Google Scholar] [CrossRef] [PubMed]
- Khatami, S.H.; Vakili, O.; Movahedpour, A.; Ghesmati, Z.; Ghasemi, H.; Taheri-Anganeh, M. Laccase: Various types and applications. Biotechnol. Appl. Biochem. 2022, 69, 2658–2672. [Google Scholar] [CrossRef]
- Hamid, R.; Khan, M.A.; Ahmad, M.; Ahmad, M.M.; Abdin, M.Z.; Musarrat, J.; Javed, S. Chitinases: An update. J. Pharm. Bioallied Sci. 2013, 5, 21–29. [Google Scholar]
- Rostami, A.; Hinc, K.; Goshadrou, F.; Shali, A.; Bayat, M.; Hassanzadeh, M.; Amanlou, M.; Eslahi, N.; Ahmadian, G. Display of B. pumilus chitinase on the surface of B. subtilis spore as a potential biopesticide. Pestic. Biochem. Physiol. 2017, 140, 17–23. [Google Scholar] [CrossRef]
- Hsieh, H.Y.; Lin, C.H.; Hsu, S.Y.; Stewart, G.C. A Bacillus spore-based display system for bioremediation of atrazine. Appl. Environ. Microbiol. 2020, 86, e01230-20. [Google Scholar] [CrossRef]
- Qu, Y.; Shi, S.; Ma, Q.; Kong, C.; Zhou, H.; Zhang, X.; Zhou, J. Multistep conversion of para-Substituted Phenols by Phenol Hydroxylase and 2,3-Dihydroxybiphenyl 1,2-Dioxygenase. Appl. Biochem. Biotechnol. 2013, 169, 2064–2075. [Google Scholar] [CrossRef]
- Seah, S.Y.K.; Labbé, G.; Nerdinger, S.; Johnson, M.R.; Snieckus, V.; Eltis, L.D. Identification of a serine hydrolase as a key determinant in the microbial degradation of polychlorinated biphenyls. J. Biol. Chem. 2000, 275, 15701–15708. [Google Scholar] [CrossRef]
- Haber, L.T.; Bates, H.K.; Allen, B.C.; Vincent, M.J.; Oller, A.R. Derivation of an oral toxicity reference value for nickel. Regul. Toxicol. Pharmacol. 2017, 87, S1–S18. [Google Scholar] [CrossRef] [PubMed]
- Tchounwou, P.B.; Yedjou, C.G.; Udensi, U.K.; Pacurari, M.; Stevens, J.J.; Patlolla, A.K.; Noubissi, F.; Kumar, S. State of the science review of the health effects of inorganic arsenic: Perspectives for future research. Environ. Toxicol. 2019, 34, 188–202. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Application | Anchor | Fusion | Function | References |
|---|---|---|---|---|
| Vaccine; adjuvant; drug delivery | CotB | C-terminal tetanus fragment (TTFC) | Intranasal dosage produced a mucosal (IgA) and systemic (IgG) reaction in murine mod | [13,14,15] |
| CotZ, CotY | SARS-CoV-2 Receptor binding domain | Oral vaccine against SARS-CoV-2 virus | [16,17] | |
| Spore | Neutralizing epitopes from Mannheimia haemolytica | Adjuvant for bovine respiratory disease | [18] | |
| Spore | Covalent linkage of curcumin and folate | Drug delivery for colon cancer | [19] | |
| Biocatalysis | CotG | β-galactosidase | Transgalactosylation in biphasic reaction mixtures | [20,21] |
| CotC, CotY, CotZ | β-galactosidase | β-gal reaction; evaluate as potential enzymatic anchors | [22] | |
| CotZ | D-psicose 3-epimerase | Allulose synthesis | [23] | |
| CotB | Lipase | Demonstrates advantage of enzyme displayed enzyme comparted to non-immobilized biocatalysis | [24] | |
| CotB | Esterase | Demonstrates advantage of enzyme displayed enzyme comparted to non-immobilized biocatalysis | [25] | |
| CotC | Alcohol dehydrogenase | Ethanol tolerance for flavor production in liquor | [26] | |
| CotG | N-acetyl-D-neuraminic acid aldolase | Synthesis of N-acetyl-D-neuraminic from N-acetyl-D-glucosamine | [27] | |
| CotG | Nitrilase | Hydrolysis of nitriles to ammonia and carboxylic acid | [28] | |
| CotC | Genomic substitution of wild-type CotC (laccase) | [29,30,31,32,33,34] | ||
| Environmental applications | CotE | Tyrosinase | Phenol polluted environments | [35] |
| CotG | Haloalkane dehalogenase | Degradation of sulfur mustard | [36] | |
| CotG | Meta-cleavage product (MCP) hydrolase (MfphA and BphD) | 2-hydroxy-6-oxohexa-2,4-dienoic acids transformation | [37] | |
| CotB | His18 | Nickel binding | [38] | |
| Spore | Not applicable | Rare earth element binding | [39] | |
| Spore | Not applicable | Arsenic sensor | [40] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmoodi, A.; Farinas, E.T. Applications of Bacillus subtilis Protein Display for Medicine, Catalysis, Environmental Remediation, and Protein Engineering. Microorganisms 2024, 12, 97. https://doi.org/10.3390/microorganisms12010097
Mahmoodi A, Farinas ET. Applications of Bacillus subtilis Protein Display for Medicine, Catalysis, Environmental Remediation, and Protein Engineering. Microorganisms. 2024; 12(1):97. https://doi.org/10.3390/microorganisms12010097
Chicago/Turabian StyleMahmoodi, Asieh, and Edgardo T. Farinas. 2024. "Applications of Bacillus subtilis Protein Display for Medicine, Catalysis, Environmental Remediation, and Protein Engineering" Microorganisms 12, no. 1: 97. https://doi.org/10.3390/microorganisms12010097