
RETRACTED: Drug Resistance and Molecular Characteristics of Carbapenem-Resistant OXA-48-Producing Klebsiella pneumoniae Strains in Hainan, China
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection and Strains Identification
2.2. Susceptibility Testing of Agents
2.3. Bacterial Conjugation
2.4. Microbiological Characteristics
2.5. Stability of blaOXA-48-Encoding Plasmid in CR-hvKP Strains
2.6. Virulence Assessment
2.7. Whole-Genome Sequencing (WGS) Analysis
2.8. Statistical Analysis
3. Result
3.1. Clinical Features of Patients Infected with OXA-48-Producing CR-hvKP Strains
3.2. Bacterial Conjugation Experiment
3.3. Plasmid Stability
3.4. Antibiotic Sensitivity of OXA-48-Producing hvKP Strains
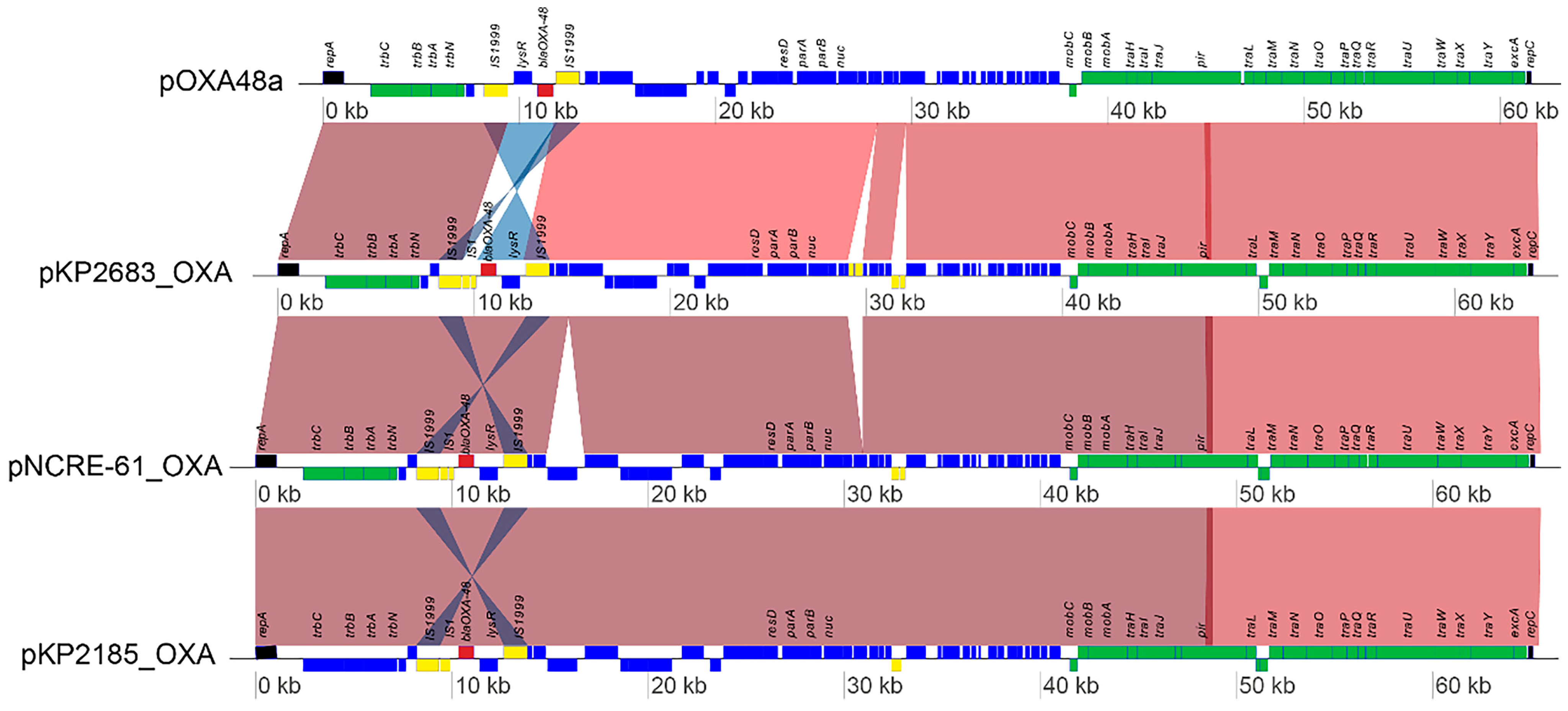
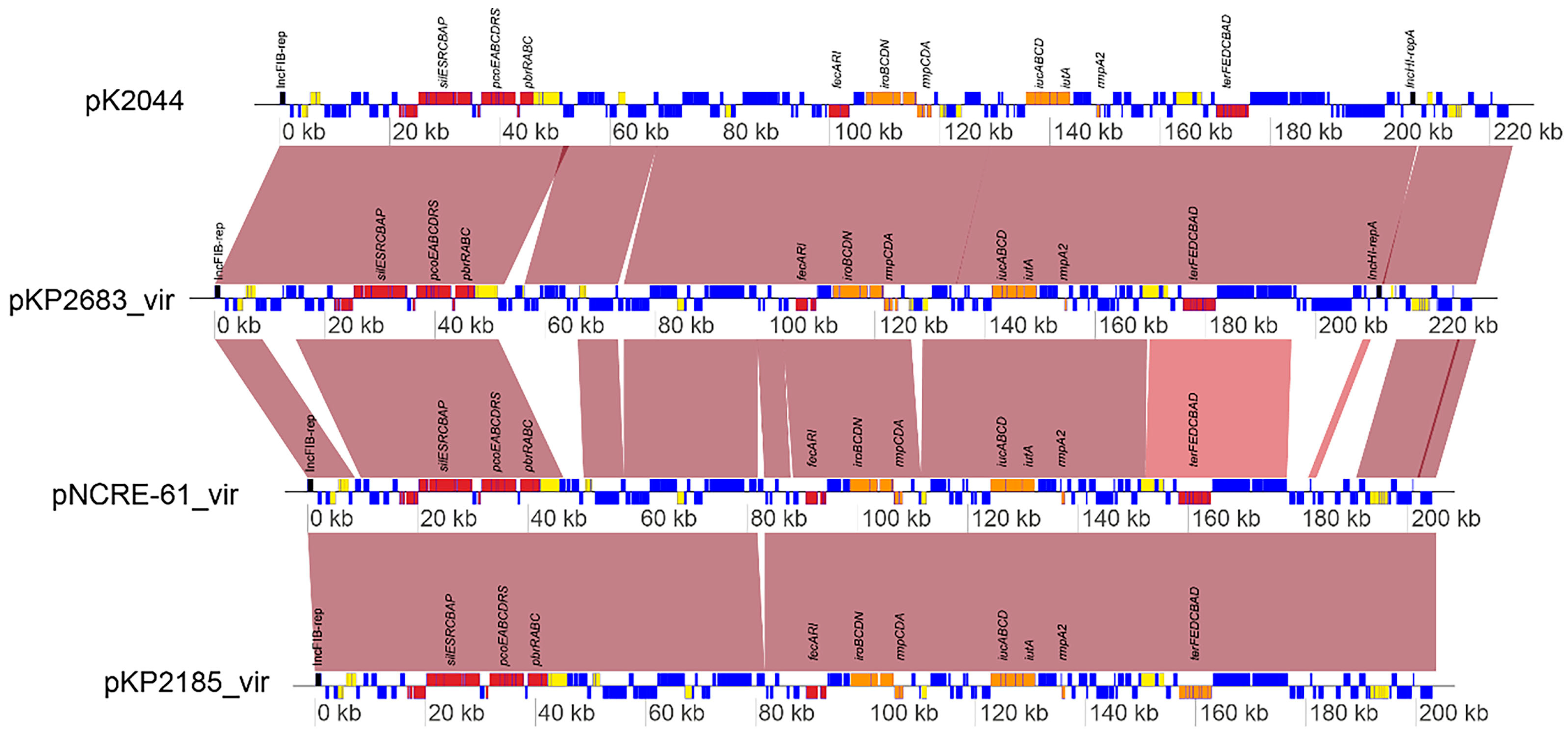
3.5. Microbiological Characteristics, In Vivo Virulence Assessment, and Genomic Analysis of OXA-48-Producing hvKP Strains
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2019 Antimicrobial Resistance Collaborators. Global mortality associated with 33 bacterial pathogens in 2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2022, 400, 2221–2248. [Google Scholar] [CrossRef]
- Loconsole, D.; Accogli, M.; De Robertis, A.L.; Capozzi, L.; Bianco, A.; Morea, A.; Mallamaci, R.; Quarto, M.; Parisi, A.; Chironna, M. Emerging high-risk ST101 and ST307 carbapenem-resistant Klebsiella pneumoniae clones from bloodstream infections in Southern Italy. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 24. [Google Scholar] [CrossRef]
- Ali, M.R.; Yang, Y.; Dai, Y.; Lu, H.; He, Z.; Li, Y.; Sun, B. Prevalence of multidrug-resistant hypervirulent Klebsiella pneumoniae without defined hypervirulent biomarkers in Anhui, China: A new dimension of hypervirulence. Front. Microbiol. 2023, 14, 1247091. [Google Scholar] [CrossRef]
- Russo, T.A.; Olson, R.; Fang, C.T.; Stoesser, N.; Miller, M.; MacDonald, U.; Hutson, A.; Barker, J.H.; La Hoz, R.M.; Johnson, J.R. Identification of Biomarkers for Differentiation of Hypervirulent Klebsiella pneumoniae from Classical K. pneumoniae. J. Clin. Microbiol. 2018, 56, e00776-18. [Google Scholar] [CrossRef]
- Yang, X.; Dong, N.; Chan, E.W.; Zhang, R.; Chen, S. Carbapenem Resistance-Encoding and Virulence-Encoding Conjugative Plasmids in Klebsiella pneumoniae. Trends Microbiol. 2021, 29, 65–83. [Google Scholar] [CrossRef]
- Han, Y.L.; Wen, X.H.; Zhao, W.; Cao, X.S.; Wen, J.X.; Wang, J.R.; Hu, Z.D.; Zheng, W.Q. Epidemiological characteristics and molecular evolution mechanisms of carbapenem-resistant hypervirulent Klebsiella pneumoniae. Front. Microbiol. 2022, 13, 1003783. [Google Scholar] [CrossRef]
- Lan, P.; Jiang, Y.; Zhou, J.; Yu, Y. A global perspective on the convergence of hypervirulence and carbapenem resistance in Klebsiella pneumoniae. J. Glob. Antimicrob. Resist. 2021, 25, 26–34. [Google Scholar] [CrossRef]
- Jin, L.; Wang, R.; Gao, H.; Wang, Q.; Wang, H. Identification of a Novel Hybrid Plasmid Encoding KPC-2 and Virulence Factors in Klebsiella pneumoniae Sequence Type 11. Antimicrob. Agents Chemother. 2021, 65, 02435-20. [Google Scholar] [CrossRef]
- Huang, Y.H.; Chou, S.H.; Liang, S.W.; Ni, C.E.; Lin, Y.T.; Huang, Y.W.; Yang, T.C. Emergence of an XDR and carbapenemase-producing hypervirulent Klebsiella pneumoniae strain in Taiwan. J. Antimicrob. Chemother. 2018, 73, 2039–2046. [Google Scholar] [CrossRef]
- Turton, J.; Davies, F.; Turton, J.; Perry, C.; Payne, Z.; Pike, R. Hybrid resistance and Virulence plasmids in ‘high-risk’ clones of Klebsiella pneumoniae, including those carrying blaNDM-5. Microorganisms 2019, 7, 326. [Google Scholar] [CrossRef]
- Tian, C.; Xing, M.; Zhao, Y.; Fan, X.; Bai, Y.; Fu, L.; Wang, S. Whole genome sequencing of OXA-232-producing wzi93-KL112-O1 carbapenem-resistant Klebsiella pneumoniae in human bloodstream infection co-harboring chromosomal ISEcp1-based blaCTX-M-15 and one rmpA2-associated virulence plasmid. Front. Cell Infect. Microbiol. 2022, 12, 984479. [Google Scholar] [CrossRef]
- Hallal Ferreira Raro, O.; Nordmann, P.; Dominguez Pino, M.; Findlay, J.; Poirel, L. Emergence of Carbapenemase-Producing Hypervirulent Klebsiella pneumoniae in Switzerland. Antimicrob. Agents Chemother. 2023, 67, e0142422. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- Yu, F.; Lv, J.; Niu, S.; Du, H.; Tang, Y.W.; Pitout, J.D.D.; Bonomo, R.A.; Kreiswirth, B.N.; Chen, L. Multiplex PCR analysis for the rapid detection of Klebsiella Pneumoniae carbapenem-resistant (ST258, 11) and hypervirulent (ST23, 65, 86, 375) strains. J. Clin. Microbiol. 2018, 56, e00731-18. [Google Scholar] [CrossRef]
- Yang, X.; Sun, Q.; Li, J.; Jiang, Y.; Li, Y.; Lin, J.; Chen, K.; Chan, E.W.; Zhang, R.; Chen, S. Molecular epidemiology of carbapenem-resistant hypervirulent Klebsiella pneumoniae in China. Emerg. Microbes. Infect. 2022, 11, 841–849. [Google Scholar] [CrossRef]
- Hu, D.; Chen, W.; Zhang, Q.; Li, M.; Yang, Z.; Wang, Y.; Huang, Y.; Li, G.; Tian, D.; Fu, P.; et al. Prevalence of Carbapenem-Resistant Hypervirulent Klebsiella pneumoniae and Hypervirulent Carbapenem-Resistant Klebsiella pneumoniae in China Determined via Mouse Lethality Tests. Front. Cell Infect. Microbiol. 2022, 12, 882210. [Google Scholar] [CrossRef]
- Lin, Y.T.; Cheng, Y.H.; Juan, C.H.; Wu, P.F.; Huang, Y.W.; Chou, S.H.; Yang, T.C.; Wang, F.D. High mortality among patients infected with hypervirulent antimicrobial-resistant capsular type K1 Klebsiella pneumoniae strains in Taiwan. Int. J. Antimicrob. Agents. 2018, 52, 251–257. [Google Scholar] [CrossRef]
- Hendrickx, A.P.A.; Landman, F.; de Haan, A.; Witteveen, S.; van Santen-Verheuvel, M.G.; Schouls, L.M. The Dutch Cpe Surveillance Study Group. blaOXA-48-like genome architecture among carbapenemase-producing Escherichia coli and Klebsiella pneumoniae in the Netherlands. Microb. Genom. 2021, 7, 000512. [Google Scholar]
- Becker, L.; Kaase, M.; Pfeifer, Y.; Fuchs, S.; Reuss, A.; von Laer, A.; Sin, M.A.; Korte-Berwanger, M.; Gatermann, S.; Werner, G. Genome-based analysis of Carbapenemase-producing Klebsiella pneumoniae isolates from German hospital patients, 2008–2014. Antimicrob. Resist. Infect. Control. 2018, 7, 62. [Google Scholar] [CrossRef]
- Simner, P.J.; Antar, A.A.R.; Hao, S.; Gurtowski, J.; Tamma, P.D.; Rock, C.; Opene, B.N.A.; Tekle, T.; Carroll, K.C.; Schatz, M.C.; et al. Antibiotic pressure on the acquisition and loss of antibiotic resistance genes in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2018, 73, 1796–1803. [Google Scholar] [CrossRef]
- Pulingam, T.; Parumasivam, T.; Gazzali, A.M.; Sulaiman, A.M.; Chee, J.Y.; Lakshmanan, M.; Chin, C.F.; Sudesh, K. Antimicrobial resistance: Prevalence, economic burden, mechanisms of resistance and strategies to overcome. Eur. J. Pharm. Sci. 2022, 170, 106103. [Google Scholar] [CrossRef]
- Bulman, Z.P.; Tan, X.; Chu, T.Y.; Huang, Y.; Rana, A.P.; Singh, N.; Flowers, S.A.; Kyono, Y.; Kreiswirth, B.N.; Chen, L. Ceftazidime-avibactam based combinations against carbapenemase producing Klebsiella pneumoniae harboring hypervirulence plasmids. Comput. Struct. Biotechnol. J. 2022, 20, 3946–3954. [Google Scholar] [CrossRef]
- Cheng, Y.H.; Huang, T.W.; Juan, C.H.; Tseng, Y.Y.; Chen, T.W.; Yang, T.C.; Lin, Y.T. Tigecycline-non-susceptible hypervirulent Klebsiella pneumoniae strains in Taiwan. J. Antimicrob. Chemother. 2020, 75, 309–317. [Google Scholar] [CrossRef]
- McDermott, P.F.; Tyson, G.H.; Kabera, C.; Chen, Y.; Li, C.; Folster, J.P.; Ayers, S.L.; Lam, C.; Tate, H.P.; Zhao, S. Whole-Genome Sequencing for Detecting Antimicrobial Resistance in Nontyphoidal Salmonella. Antimicrob. Agents Chemother. 2016, 60, 5515–5520. [Google Scholar] [CrossRef]
- Yang, J.; Long, H.; Hu, Y.; Feng, Y.; McNally, A.; Zong, Z. Klebsiella oxytoca Complex: Update on Taxonomy, Antimicrobial Resistance, and Virulence. Clin. Microbiol. Rev. 2022, 35, e0000621. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Lam, M.M.C.; Wick, R.R.; Watts, S.C.; Cerdeira, L.T.; Wyres, K.L.; Holt, K.E. A genomic surveillance framework and genotyping tool for Klebsiella pneumoniae and its related species complex. Nat. Commun. 2021, 12, 4188. [Google Scholar] [CrossRef]
- Lam, M.M.C.; Wick, R.R.; Judd, L.M.; Holt, K.E.; Wyres, K.L. Kaptive 2.0: Updated capsule and lipopolysaccharide locus typing for the Klebsiella pneumoniae species complex. Microb. Genom. 2022, 8, 000800. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Case | NCRE61 | KP2185 | KP2683-1 |
|---|---|---|---|
| Invasive procedures and devices at the time of culture | endotracheal intubation and a mechanical ventilator | tracheostomy urinary catheter, central venous catheter, | central venous catheter |
| Diagnosis Location at time of culture | respiratory medicine, ordinary ward | general surgery ordinary ward | bacteremia intensive care unit |
| 28-day mortality or survivor | survivor | survivor | survivor |
| Prior antibiotic exposure within 1 month | imipenem | piperacillin | piperacillin, meropenem, |
| Definitive antimicrobial therapy | tigecycline and piperacillin | meropenem | meropenem and tigecycline |
| Co-morbidity | chronic obstructive pulmonary disease | diabetes cerebral infarction | hypertension diabetes |
| Length of stay after infection: days | 64 days | 55 days | 73 days |
| Community- or hospital-acquired infection | hospital-acquired infection | hospital-acquired infection | hospital-acquired infection |
| Age, years | 68 | 71 | 62 |
| Gender | male | male | male |
| Strain | Conjugational Efficiency | Minimal Inhibitory Concentration (μg/mL) | ||
|---|---|---|---|---|
| Ertapenem | Imipenem | Meropenem | ||
| NCRE61 transconjugant | 8.2 × 10−4 | 0.5 | 2 | 0.38 |
| KP2683-1 transconjugant | 5 × 10−5 | 3 | 4 | 0.5 |
| KP2185 transconjugant | 3.5 × 10−4 | 0.38 | 3 | 0.38 |
| E. coli J53 | - | 0.003 | 0.19 | 0.016 |
| MIC (mg/L) a | |||
|---|---|---|---|
| Antibiotics | NCRE61 | KP2185 | KP2683-1 |
| Tigecycline | 1.0 | 0.5 | 0.75 |
| Imipenem | 3 | 2 | 2 |
| Ertapenem | 4 | 12 | 8 |
| Colistin | 4 | 2 | 1 |
| Ceftazidime/avibactam | 1.0 | 1.0 | 0.25 |
| Piperacillin/Tazobactam | ≥128 | ≥128 | ≤128 |
| Cefepime | ≥64 | ≥64 | ≤1 |
| Ceftriaxone | ≥64 | ≥64 | ≤1 |
| Cefuroxime | ≥64 | ≥64 | 4 |
| Cefazolin | ≥64 | ≥64 | 16 |
| Ceftazidime | 16 | ≥64 | ≤1 |
| Ciprofloxacin | ≥4 | ≥4 | ≤0.25 |
| Levofloxacin | ≥8 | ≥8 | ≤0.12 |
| Gentamicin | ≥16 | ≥16 | ≤1 |
| Amikacin | ≥64 | ≥64 | ≤2 |
| Strain No | NCRE61 | KP2185 | KP2683-1 |
|---|---|---|---|
| Virulence factors associated with hypervirulent strains | ΔrmpA, rmpD, rmpC, rmpA2, iroBCDN, iucABCD-iutA, ybtSXQPAUTE, irp1, irp2 | ΔrmpA, rmpD, rmpC, rmpA2, iroBCDN, iucABCD-iutA | rmpADC, ΔrmpA2, iroBCDN, iucABCD-iutA, ybtSXQPAUTE, irp1, irp2 |
| β-lactamase resistance gene | OXA-48, DHA-1, CTX-M-14, TEM-1, SHV-11 | OXA-48, DHA-1, CTX-M-14, TEM-1, SHV-11 | OXA-48, SHV-11 |
| LD50 | 2.4 × 106 | 5.1 × 106 | 2.1 × 103 |
| Capsular type | K64 | K64 | K1 |
| ST type | ST11 | ST11 | ST23 |
| Date of isolation | 22 March 2022 | 2 December 2022 | 28 January 2023 |
| Specimen | Bronchoalveolar lavage fluid | Liver abscess | Blood |
| Strain No | Antimicrobial Resistant Genes | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| β-Lactamases | Aminoglycoside | Fluoroquinolones | Chloramphenicol | Tetracycline | Macrolides | Rifampin | Sulfonamides | Trimethoprim | |
| NCRE61 | blaOXA-48, blaSHV-11, blaTEM-1, blaDHA-1, blaCTX-M-14 | strB, strA, rmtB, aac3-IId, aadA16, aadA2, aph(3″)-Ib, aph(6)-Id, aac(6′)-Ib-cr5 | qnrB4 | floR2 | tetA, tetG | - | arr3 | sul1, sul2 | dfrA27, dfrA12 |
| KP2683-1 | blaOXA-48, blaSHV-11 | - | - | - | - | - | - | - | - |
| KP2185 | blaOXA-48, blaSHV-11, blaTEM-1, blaDHA-1, blaCTX-M-14 | strB, strA, rmtB, aac3-IId, aadA2, aph(3″)-Ib, aph(6)-Id | GyrA-83I; ParC-80I, qnrB4 | floR2 | tetG | erm42 | - | sul1, sul2 | dfrA12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, M.; Liu, L.; Liu, B.; Zhou, X.; Li, Q. RETRACTED: Drug Resistance and Molecular Characteristics of Carbapenem-Resistant OXA-48-Producing Klebsiella pneumoniae Strains in Hainan, China. Microorganisms 2024, 12, 49. https://doi.org/10.3390/microorganisms12010049
Ye M, Liu L, Liu B, Zhou X, Li Q. RETRACTED: Drug Resistance and Molecular Characteristics of Carbapenem-Resistant OXA-48-Producing Klebsiella pneumoniae Strains in Hainan, China. Microorganisms. 2024; 12(1):49. https://doi.org/10.3390/microorganisms12010049
Chicago/Turabian StyleYe, Min, Lei Liu, Bin Liu, Xiangdong Zhou, and Qi Li. 2024. "RETRACTED: Drug Resistance and Molecular Characteristics of Carbapenem-Resistant OXA-48-Producing Klebsiella pneumoniae Strains in Hainan, China" Microorganisms 12, no. 1: 49. https://doi.org/10.3390/microorganisms12010049