
Enhanced Expression of Alcohol Dehydrogenase I in Pichia pastoris Reduces the Content of Acetaldehyde in Wines
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal and Bacterial Strains, Plasmids, Reagents, and Medium
2.2. Construction of Plasmids
2.3. Construction and Cultivation of Recombinant Strains of Pichia pastoris
2.4. Extraction of Intracellular Proteins
2.5. Detection of the Enzymatic Activity of ADH1
2.6. Purification of ADH1
2.7. Detection of the Content of Acetaldehyde in Wine
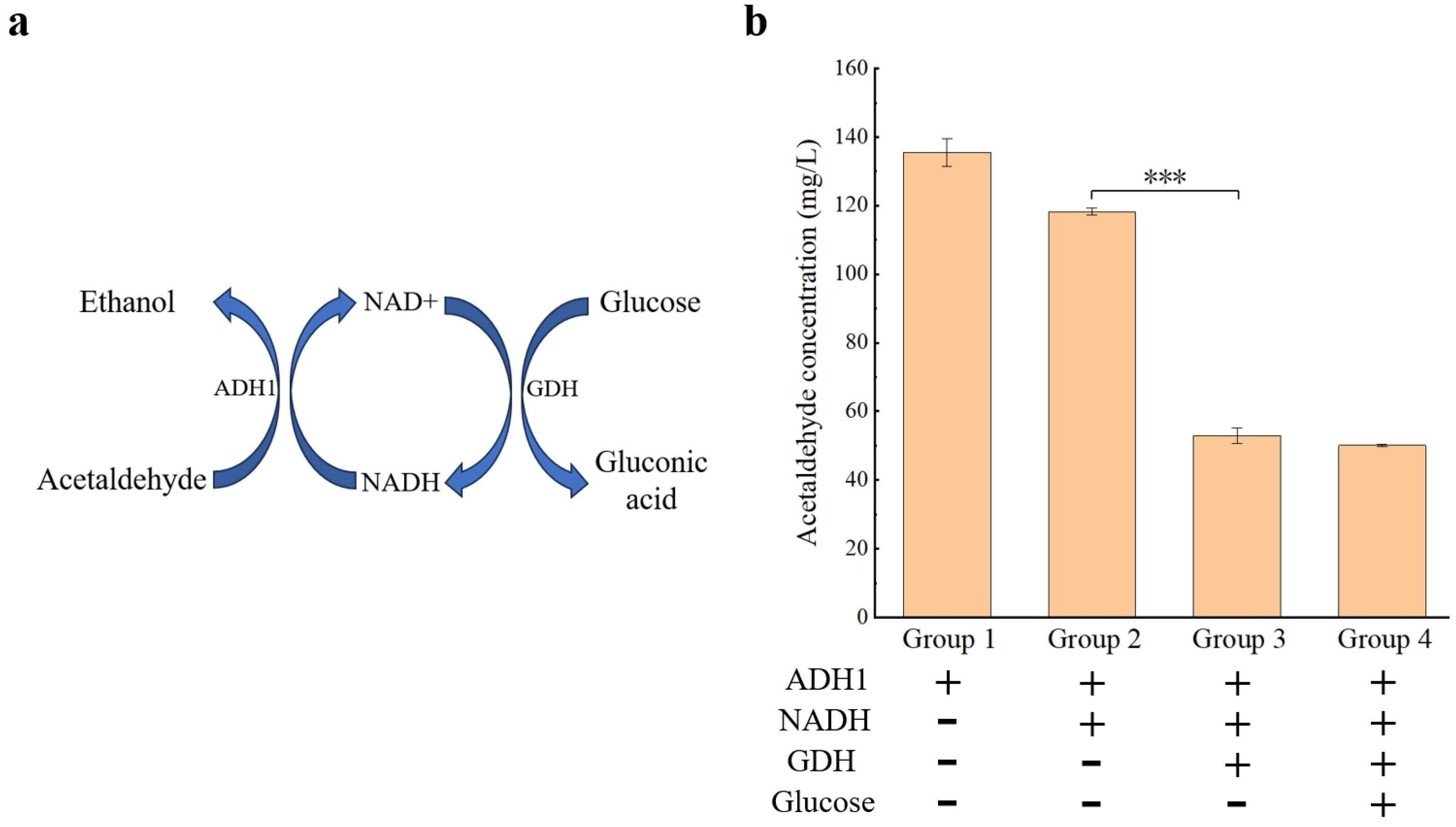
2.8. Application of ADH1 in Reducing the Content of Acetaldehyde in Wine
2.9. Statistical Analysis
3. Results
3.1. Construction of Expression Vectors
3.2. Expression of ADH1 in Pichia pastoris GS115
3.3. Purification of ADH1
3.4. Reduced Content of Acetaldehyde in Wine by ADH1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, S.Q.; Pilone, G.J. An overview of formation and roles of acetaldehyde in winemaking with emphasis on microbiological implications. Int. J. Food Sci. Technol. 2000, 35, 49–61. [Google Scholar] [CrossRef]
- Ochando, T.; Mouret, J.R.; Humbert-Goffard, A.; Aguera, E.; Sablayrolles, J.M.; Farines, V. Comprehensive study of the dynamic interaction between SO2 and acetaldehyde during alcoholic fermentation. Food Res. Int. 2020, 136, 109607. [Google Scholar] [CrossRef] [PubMed]
- Rousseva, M.; Kontoudakis, N.; Schmidtke, L.M.; Scollary, G.R.; Clark, A.C. Impact of wine production on the fractionation of copper and iron in Chardonnay wine: Implications for oxygen consumption. Food Chem. 2016, 203, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Danilewicz, J.C. Mechanism of Autoxidation of Polyphenols and Participation of Sulfite in Wine: Key Role of Iron. Am. J. Enol. Vitic. 2011, 62, 319–328. [Google Scholar] [CrossRef]
- Zea, L.; Serratosa, M.P.; Merida, J.; Moyano, L. Acetaldehyde as Key Compound for the Authenticity of Sherry Wines: A Study Covering 5 Decades. Compr. Rev. Food Sci. Food Saf. 2015, 14, 681–693. [Google Scholar] [CrossRef]
- Cucciniello, R.; Forino, M.; Picariello, L.; Coppola, F.; Moio, L.; Gambuti, A. How acetaldehyde reacts with low molecular weight phenolics in white and red wines. Eur. Food Res. Technol. 2021, 247, 2935–2944. [Google Scholar] [CrossRef]
- Sheridan, M.K.; Elias, R.J. Reaction of Acetaldehyde with Wine Flavonoids in the Presence of Sulfur Dioxide. J. Agric. Food Chem. 2016, 64, 8615–8624. [Google Scholar] [CrossRef]
- Garcia, L.; Perrin, C.; Nolleau, V.; Godet, T.; Farines, V.; Garcia, F.; Caille, S.; Saucier, C. Impact of Acetaldehyde Addition on the Sensory Perception of Syrah Red Wines. Foods 2022, 11, 1693. [Google Scholar] [CrossRef]
- Sheridan, M.K.; Elias, R.J. Exogenous acetaldehyde as a tool for modulating wine color and astringency during fermentation. Food Chem. 2015, 177, 17–22. [Google Scholar] [CrossRef]
- Secretan, B.; Straif, K.; Baan, R.; Grosse, Y.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Freeman, C.; Galichet, L.; et al. A review of human carcinogens-Part E: Tobacco, areca nut, alcohol, coal smoke, and salted fish. Lancet Oncol. 2009, 10, 1033–1034. [Google Scholar] [CrossRef]
- Lachenmeier, D.W.; Monakhova, Y.B. Short-term salivary acetaldehyde increase due to direct exposure to alcoholic beverages as an additional cancer risk factor beyond ethanol metabolism. J. Exp. Clin. Cancer Res. 2011, 30, 3. [Google Scholar] [CrossRef] [PubMed]
- Ntuli, R.G.; Saltman, Y.; Ponangi, R.; Jeffery, D.W.; Bindon, K.; Wilkinson, K.L. Impact of fermentation temperature and grape solids content on the chemical composition and sensory profiles of Cabernet Sauvignon wines made from flash détente treated must fermented off-skins. Food Chem. 2022, 369, 130861. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.M.; Sun, Y.H.; Liu, M.Q.; Cui, X.Q.; Wang, J.Q.; Li, J.M.; Han, G.M. Influence of Oxygen Management during the Post-Fermentation Stage on Acetaldehyde, Color, and Phenolics of Vitis vinifera L. Cv. Cabernet Sauvignon Wine. Molecules 2022, 27, 6692. [Google Scholar] [CrossRef] [PubMed]
- Jackowetz, J.N.; Dierschke, S.; de Orduña, R.M. Multifactorial analysis of acetaldehyde kinetics during alcoholic fermentation by Saccharomyces cerevisiae. Food Res. Int. 2011, 44, 310–316. [Google Scholar] [CrossRef]
- Osborne, J.P.; Dubé Morneau, A.; Mira de Orduña, R. Degradation of free and sulfur-dioxide-bound acetaldehyde by malolactic lactic acid bacteria in white wine. J. Appl. Microbiol. 2006, 101, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Liu, L.J.; Wei, Y.Z.; Li, G.P.; Yue, X.L.; An, L.Z. Metabolite Profiling of adh1 Mutant Response to Cold Stress in Arabidopsis. Front. Plant Sci. 2017, 7, 2072. [Google Scholar] [CrossRef]
- de Smidt, O.; du Preez, J.C.; Albertyn, J. The alcohol dehydrogenases of Saccharomyces cerevisiae: A comprehensive review. FEMS Yeast Res. 2008, 8, 967–978. [Google Scholar] [CrossRef]
- Leskovac, V.; Trivic, S.; Pericin, D. The three zinc-containing alcohol dehydrogenases from baker’s yeast, Saccharomyces cerevisiae. FEMS Yeast Res. 2002, 2, 481–494. [Google Scholar] [CrossRef]
- Raj, S.B.; Ramaswamy, S.; Plapp, B.V. Yeast Alcohol Dehydrogenase Structure and Catalysis. Biochemistry 2014, 53, 5791–5803. [Google Scholar] [CrossRef]
- Magonet, E.; Hayen, P.; Delforge, D.; Delaive, E.; Remacle, J. Importance of the structural zinc atom for the stability of yeast alcohol dehydrogenase. Biochem. J. 1992, 287 Pt 2, 361–365. [Google Scholar] [CrossRef]
- Yang, Z.L.; Zhang, Z.S. Engineering strategies for enhanced production of protein and bio-products in Pichia pastoris: A review. Biotechnol. Adv. 2018, 36, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Hirz, M.; Pichler, H.; Schwab, H. Protein expression in Pichia pastoris: Recent achievements and perspectives for heterologous protein production. Appl. Microbiol. Biotechnol. 2014, 98, 5301–5317. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.X.; Gong, J.S.; Shi, J.H.; Liu, C.F.; Li, H.; Su, C.; Jiang, M.; Xu, Z.H.; Shi, J.S. High-efficiency secretory expression and characterization of the recombinant type III human-like collagen in Pichia pastoris. Bioresour. Bioprocess. 2022, 9, 117. [Google Scholar] [CrossRef]
- Li, C.; Lin, Y.; Zheng, X.Y.; Pang, N.; Liao, X.H.; Liu, X.X.; Huang, Y.Y.; Liang, S.L. Combined strategies for improving expression of Citrobacter amalonaticus phytase in Pichia pastoris. BMC Biotechnol. 2015, 15, 88. [Google Scholar] [CrossRef] [PubMed]
- Hartner, F.S.; Ruth, C.; Langenegger, D.; Johnson, S.N.; Hyka, P.; Lin-Cereghino, G.P.; Lin-Cereghino, J.; Kovar, K.; Cregg, J.M.; Glieder, A. Promoter library designed for fine-tuned gene expression in Pichia pastoris. Nucleic Acids Res. 2008, 36, e76. [Google Scholar] [CrossRef] [PubMed]
- Karaoglan, M.; Karaoglan, F.E.; Inan, M. Comparison of ADH3 promoter with commonly used promoters for recombinant protein production in Pichia pastoris. Protein Expr. Purif. 2016, 121, 112–117. [Google Scholar] [CrossRef]
- Chen, M.Q.; Zeng, X.; Zhu, Q.J.; Wang, D.G.; Han, S.Y.; Liang, S.L.; Lin, Y. Effective synthesis of Rebaudioside A by whole-cell biocatalyst Pichia pastoris. Biochem. Eng. J. 2021, 175, 108117. [Google Scholar] [CrossRef]
- Suwannarangsee, S.; Oh, D.B.; Seo, J.W.; Kim, C.H.; Rhee, S.K.; Kang, H.A.; Chulalaksananukul, W.; Kwon, O. Characterization of alcohol dehydrogenase 1 of the thermotolerant methylotrophic yeast Hansenula polymorpha. Appl. Microbiol. Biotechnol. 2010, 88, 497–507. [Google Scholar] [CrossRef]
- Han, G.M.; Wang, H.; Webb, M.R.; Waterhouse, A.L. A rapid, one step preparation for measuring selected free plus SO2-bound wine carbonyls by HPLC-DAD/MS. Talanta 2015, 134, 596–602. [Google Scholar] [CrossRef]
- Pongtharangkul, T.; Chuekitkumchorn, P.; Suwanampa, N.; Payongsri, P.; Honda, K.; Panbangred, W. Kinetic properties and stability of glucose dehydrogenase from Bacillus amyloliquefaciens SB5 and its potential for cofactor regeneration. Amb Express 2015, 5, 68. [Google Scholar] [CrossRef]
- Campo-Martinez, J.F.; Gonzalez-Castro, M.J.; Ensenat-Berea, M.L.; Fernandez-Paz, J. Validation of an Automated Enzymatic Method for the Determination of Fermentable Sugars in Wines. Food Anal. Methods 2022, 15, 1851–1858. [Google Scholar] [CrossRef]
- Wood, D.W. New trends and affinity tag designs for recombinant protein purification. Curr. Opin. Struct. Biol. 2014, 26, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.B.; Ben Ali, Y.; Aissa, I.; Gargouri, Y. Eukaryotic Expression System Pichia pastoris Affects the Lipase Catalytic Properties: A Monolayer Study. PLoS ONE 2014, 9, e104221. [Google Scholar] [CrossRef]
- Esposito, D.; Chatterjee, D.K. Enhancement of soluble protein expression through the use of fusion tags. Curr. Opin. Biotechnol. 2006, 17, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Yeon, Y.J.; Park, H.J.; Park, H.Y.; Yoo, Y.J. Effect of His-tag location on the catalytic activity of 3-hydroxybutyrate dehydrogenase. Biotechnol. Bioprocess Eng. 2014, 19, 798–802. [Google Scholar] [CrossRef]
- Sabaty, M.; Grosse, S.; Adryanczyk, G.; Boiry, S.; Biaso, F.; Arnoux, P.; Pignol, D. Detrimental effect of the 6 His C-terminal tag on YedY enzymatic activity and influence of the TAT signal sequence on YedY synthesis. BMC Biochem. 2013, 14, 28. [Google Scholar] [CrossRef]
- Aslantas, Y.; Surmeli, N.B. Effects of N-Terminal and C-Terminal Polyhistidine Tag on the Stability and Function of the Thermophilic P450 CYP119. Bioinorg. Chem. Appl. 2019, 2019, 8080697. [Google Scholar] [CrossRef]
- Meng, L.J.; Liu, Y.Y.; Yin, X.J.; Zhou, H.S.; Wu, J.P.; Wu, M.B.; Yang, L.R. Effects of His-tag on Catalytic Activity and Enantioselectivity of Recombinant Transaminases. Appl. Biochem. Biotechnol. 2020, 190, 880–895. [Google Scholar] [CrossRef]
- Takagi, S.; Tsutsumi, N.; Terui, Y.; Kong, X.Y.; Yurimoto, H.; Sakai, Y. Engineering the expression system for Komagataella phaffii (Pichia pastoris): An attempt to develop a methanol-free expression system. FEMS Yeast Res. 2019, 19, foz059. [Google Scholar] [CrossRef]
- Gätjen, D.; Tomszak, F.; Dettmann, J.C.; Droste, M.; Nölle, V.; Wieczorek, M. Design of a novel switchable antibody display system in Pichia pastoris. Appl. Microbiol. Biotechnol. 2022, 106, 6209–6224. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, Y.W.; Zhang, X.; Yan, C.L.; Zhang, Y.H.; Xu, X.X.; Zhang, W. Discovery of a rhamnose utilization pathway and rhamnose-inducible promoters in Pichia pastoris. Sci. Rep. 2016, 6, 27352. [Google Scholar] [CrossRef] [PubMed]
- Uzbas, F.; Sezerman, U.; Hartl, L.; Kubicek, C.P.; Seiboth, B. A homologous production system for Trichoderma reesei secreted proteins in a cellulase-free background. Appl. Microbiol. Biotechnol. 2012, 93, 1601–1608. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.L.; Zou, C.J.; Lin, Y.; Zhang, X.W.; Ye, Y.R. Identification and characterization of P-GCW14: A novel, strong constitutive promoter of Pichia pastoris. Biotechnol. Lett. 2013, 35, 1865–1871. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.L.; Qian, J.C.; Yao, G.F.; Zhuang, Y.P.; Zhang, S.L.; Chu, J. GAP Promoter Library for Fine-Tuning of Gene Expression in Pichia pastoris. Appl. Environ. Microbiol. 2011, 77, 3600–3608. [Google Scholar] [CrossRef]
- Nong, L.Y.; Zhang, Y.M.; Duan, Y.H.; Hu, S.L.; Lin, Y.; Liang, S.L. Engineering the regulatory site of the catalase promoter for improved heterologous protein production in Pichia pastoris. Biotechnol. Lett. 2020, 42, 2703–2709. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lin, Y.; Zheng, X.Y.; Yuan, Q.Y.; Pang, N.; Liao, X.H.; Huang, Y.Y.; Zhang, X.Y.; Liang, S.L. Recycling of a selectable marker with a self-excisable plasmid in Pichia pastoris. Sci. Rep. 2017, 7, 11113. [Google Scholar] [CrossRef]
- Weninger, A.; Fischer, J.E.; Raschmanová, H.; Kniely, C.; Vogl, T.; Glieder, A. Expanding the CRISPR/Cas9 toolkit for Pichia pastoris with efficient donor integration and alternative resistance markers. J. Cell. Biochem. 2018, 119, 3183–3198. [Google Scholar] [CrossRef]
- Yang, J.J.; Nie, L.; Chen, B.; Liu, Y.M.; Kong, Y.M.; Wang, H.B.; Diao, L.Y. Hygromycin-resistance vectors for gene expression in Pichia pastoris. Yeast 2014, 31, 115–125. [Google Scholar] [CrossRef]
- Marrufo-Curtido, A.; Ferreira, V.; Escudero, A. Can aldehyde accumulation rates of red wines undergoing oxidation be predicted in accelerated conditions? The controverted role of aldehyde-polyphenol reactivity. J. Sci. Food Agric. 2022, 102, 3869–3878. [Google Scholar] [CrossRef]
- Osborne, J.P.; Mira de Orduña, R.; Pilone, G.J.; Liu, S.Q. Acetaldehyde metabolism by wine lactic acid bacteria. FEMS Microbiol. Lett. 2000, 191, 51–55. [Google Scholar] [CrossRef]
- Guittin, C.; Maçna, F.; Picou, C.; Perez, M.; Barreau, A.; Poitou, X.; Sablayrolles, J.M.; Mouret, J.R.; Farines, V. New Online Monitoring Approaches to Describe and Understand the Kinetics of Acetaldehyde Concentration during Wine Alcoholic Fermentation: Access to Production Balances. Fermentation 2023, 9, 299. [Google Scholar] [CrossRef]
- Picariello, L.; Slaghenaufi, D.; Ugliano, M. Fermentative and post-fermentative oxygenation of Corvina red wine: Influence on phenolic and volatile composition, colour and wine oxidative response. J. Sci. Food Agric. 2020, 100, 2522–2533. [Google Scholar] [CrossRef] [PubMed]
- Korban, A.; Zavoralová, L.; Cabala, R.; Egorov, V. Perspectives on using ethanol as an internal standard for the quantification of volatile compounds in alcoholic products by headspace SPME- GC/FID without sample preparation. Monatsh. Chem. 2023, 154, 987–991. [Google Scholar] [CrossRef]
- Marisco, G.; Saito, S.T.; Ganda, I.S.; Brendel, M.; Pungartnik, C. Low ergosterol content in yeast adh1 mutant enhances chitin maldistribution and sensitivity to paraquat-induced oxidative stress. Yeast 2011, 28, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Sunder, A.V.; Singh, P.; Wangikar, P.P. Characterization and Application of a Robust Glucose Dehydrogenase from Paenibacillus pini for Cofactor Regeneration in Biocatalysis. Indian J. Microbiol. 2020, 60, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Peinado, R.A.; Mauricio, J.C.; Moreno, J. Aromatic series in sherry wines with gluconic acid subjected to different biological aging conditions by Saccharomyces cerevisiae var. capensis. Food Chem. 2006, 94, 232–239. [Google Scholar] [CrossRef]
- Li, X.Y.; Huang, J.Y.; Zhou, Q.; Xu, Y.Y.; Prabhu, P.; Zhang, Y.W. Immobilization of Alcohol Dehydrogenase, Acetaldehyde Lyase, and NADH Oxidase for Cascade Enzymatic Conversion of Ethanol to Acetoin. Energies 2022, 15, 4242. [Google Scholar] [CrossRef]
- Xu, X.; Niu, C.T.; Liu, C.F.; Li, Q. Unraveling the Mechanisms for Low-Level Acetaldehyde Production during Alcoholic Fermentation in Saccharomyces pastorianus Lager Yeast. J. Agric. Food Chem. 2019, 67, 2020–2027. [Google Scholar] [CrossRef]
- Xu, X.; Bao, M.; Niu, C.T.; Wang, J.J.; Liu, C.F.; Zheng, F.Y.; Li, Y.X.; Li, Q. Engineering the cytosolic NADH availability in lager yeast to improve the aroma profile of beer. Biotechnol. Lett. 2019, 41, 363–369. [Google Scholar] [CrossRef]
- Xu, X.; Song, Y.M.; Guo, L.Y.; Cheng, W.Q.; Niu, C.T.; Wang, J.J.; Liu, C.F.; Zheng, F.Y.; Zhou, Y.; Li, X.G.; et al. Higher NADH Availability of Lager Yeast Increases the Flavor Stability of Beer. J. Agric. Food Chem. 2020, 68, 584–590. [Google Scholar] [CrossRef]
- Xu, X.; Wang, J.J.; Bao, M.; Niu, C.T.; Liu, C.F.; Zheng, F.Y.; Li, Y.X.; Li, Q. Reverse metabolic engineering in lager yeast: Impact of the NADH/NAD+ ratio on acetaldehyde production during the brewing process. Appl. Microbiol. Biotechnol. 2019, 103, 869–880. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Effect | Efficiency | Reference |
|---|---|---|---|
| Temperature | With the fermentation temperature adjusted from 16 to 24 °C, the acetaldehyde content is decreased from 19 to 13 mg/L | 31.58% | [12] |
| Ventilation | The acetaldehyde content is decreased from 10.92 to 4.21 mg/L in low-level oxygenation compared with high-level oxygenation | 61.45% | [13] |
| SO2 | The addition of 30 mg/L SO2 increases the acetaldehyde content from 24 to 36 mg/L | 33.33% | [14] |
| Microorganism | By introducing Oenococcus oeni into the malolactic fermentation, the acetaldehyde content of white wine is reduced from 90 mg/L to almost none | Over 90% | [15] |
| Plasmid | Expression Cassette | Copy Number of Gene ADH1 |
|---|---|---|
| pPIC9K-ADH1N-6×His | PAOX1-ADH1N-6×His-AOX1 TT | 1 |
| pPIC9K-ADH1C-6×His | PAOX1-ADH1C-6×His-AOX1 TT | 1 |
| pHKA-ADH1N-6×His | PAOX1-ADH1N-6×His-AOX1 TT | 1 |
| pHKA-ADH1N-6×His-2Copies | PAOX1-ADH1N-6×His-AOX1 TT | 2 |
| pHKAOXm-ADH1N-6×His | PAOXm-ADH1N-6×His-AOX1 TT | 1 |
| pHKAOXm-ADH1N-6×His-2Copies | PAOXm-ADH1N-6×His-AOX1 TT | 2 |
| pHKAOXm-ADH1N-6×His-3Copies | PAOXm-ADH1N-6×His-AOX1 TT | 3 |
| pHKADH3-ADH1N-6×His | PADH3-ADH1N-6×His-AOX1 TT | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geng, K.; Lin, Y.; Zheng, X.; Li, C.; Chen, S.; Ling, H.; Yang, J.; Zhu, X.; Liang, S. Enhanced Expression of Alcohol Dehydrogenase I in Pichia pastoris Reduces the Content of Acetaldehyde in Wines. Microorganisms 2024, 12, 38. https://doi.org/10.3390/microorganisms12010038
Geng K, Lin Y, Zheng X, Li C, Chen S, Ling H, Yang J, Zhu X, Liang S. Enhanced Expression of Alcohol Dehydrogenase I in Pichia pastoris Reduces the Content of Acetaldehyde in Wines. Microorganisms. 2024; 12(1):38. https://doi.org/10.3390/microorganisms12010038
Chicago/Turabian StyleGeng, Kun, Ying Lin, Xueyun Zheng, Cheng Li, Shuting Chen, He Ling, Jun Yang, Xiangyu Zhu, and Shuli Liang. 2024. "Enhanced Expression of Alcohol Dehydrogenase I in Pichia pastoris Reduces the Content of Acetaldehyde in Wines" Microorganisms 12, no. 1: 38. https://doi.org/10.3390/microorganisms12010038