

Impact of Sleeve Gastrectomy on Fecal Microbiota in Individuals with Morbid Obesity
 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
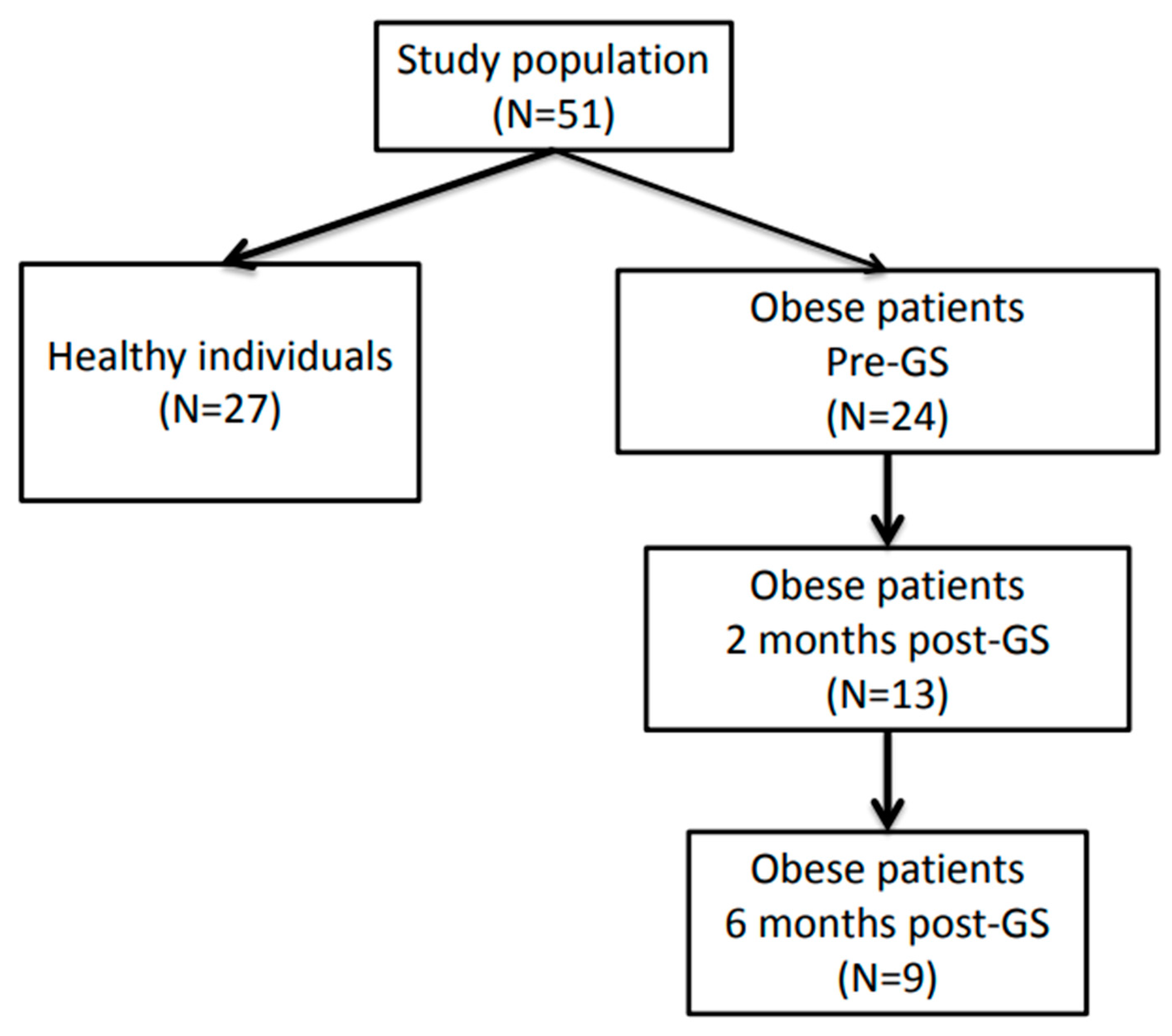
2.1. Individuals with Morbid Obesity
2.2. Control Subjects
2.3. Informed Consent Procedure
2.4. Biomarkers in Feces and Blood and Determination of Intestinal Permeability
2.5. DNA Isolation of Fecal Samples
2.6. Sequencing
2.7. Data Analysis
3. Results
3.1. Study Population
3.2. Biomarkers
3.3. Sequencing Data
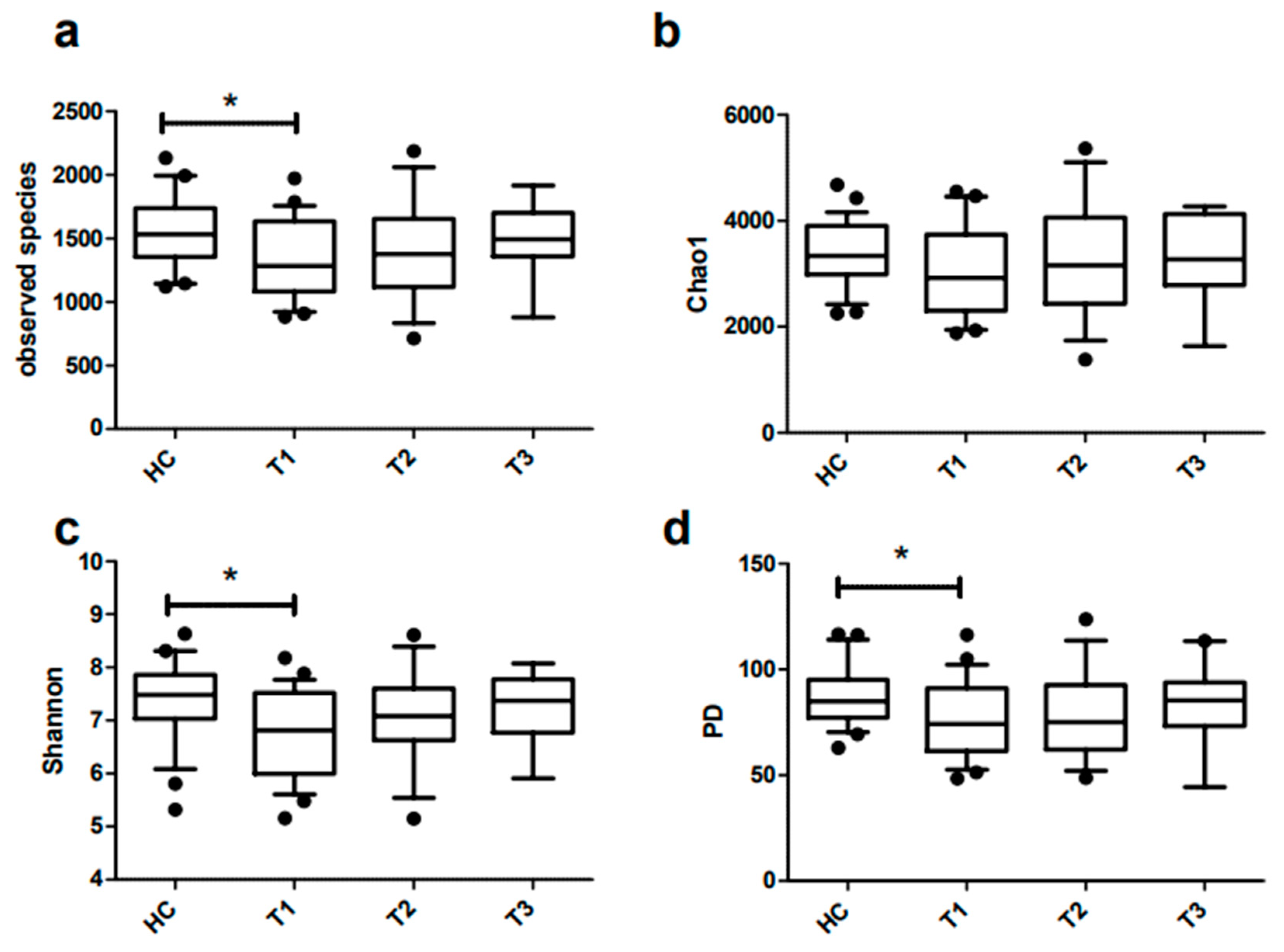
3.4. Richness and Diversity of Fecal Microbiota of Obese and Lean Individuals
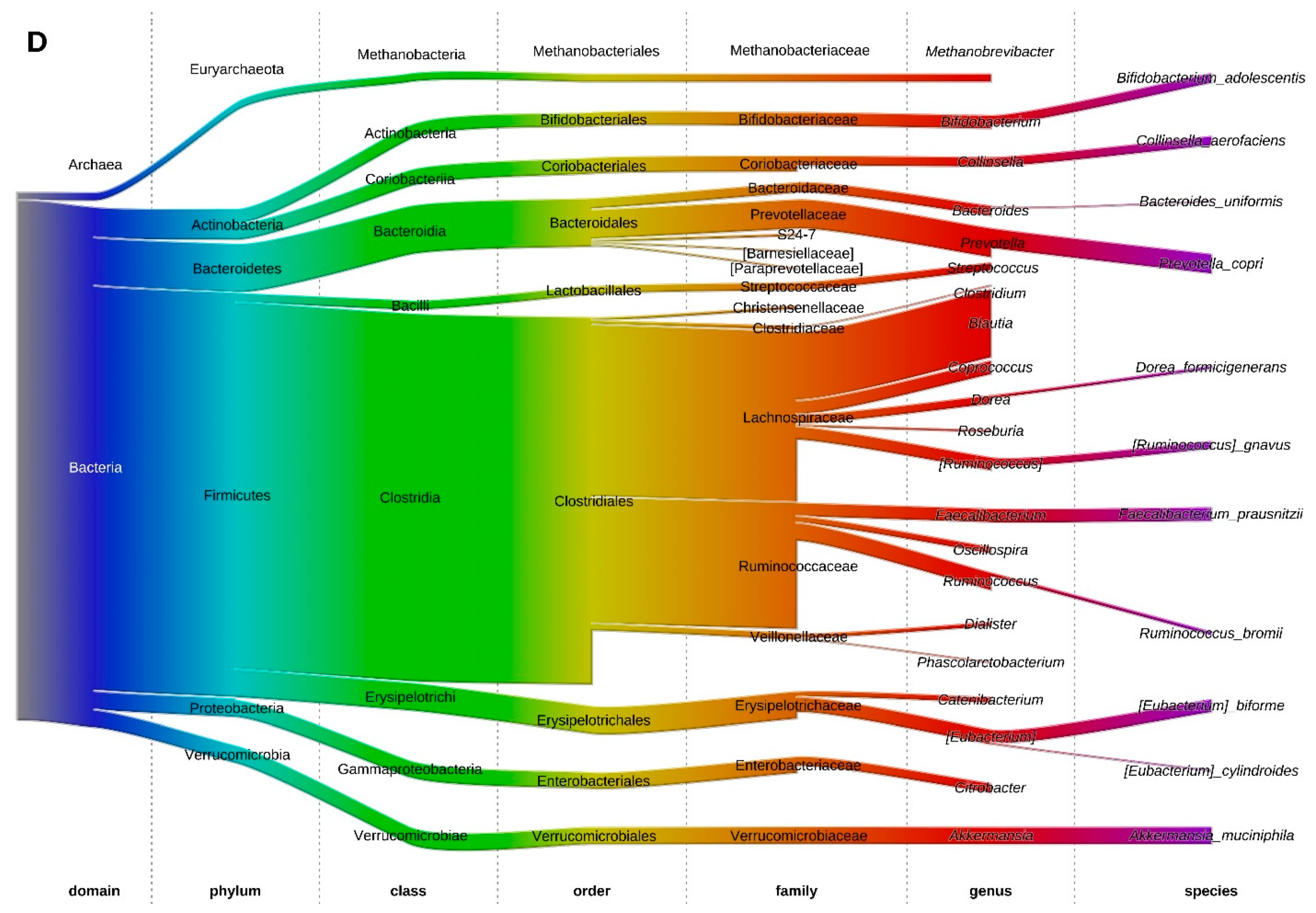
3.5. Fecal Microbiota at the Phylum Level
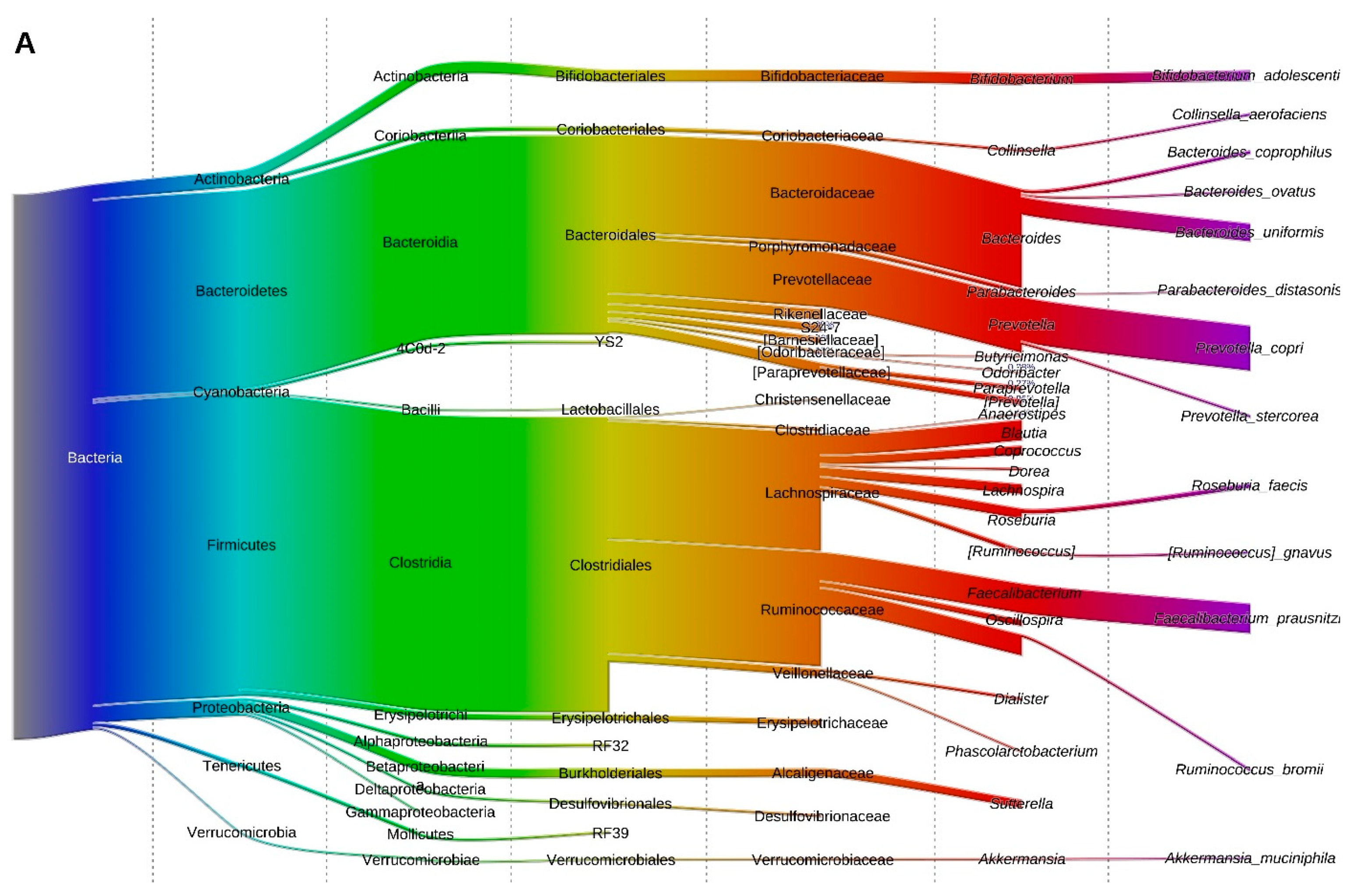
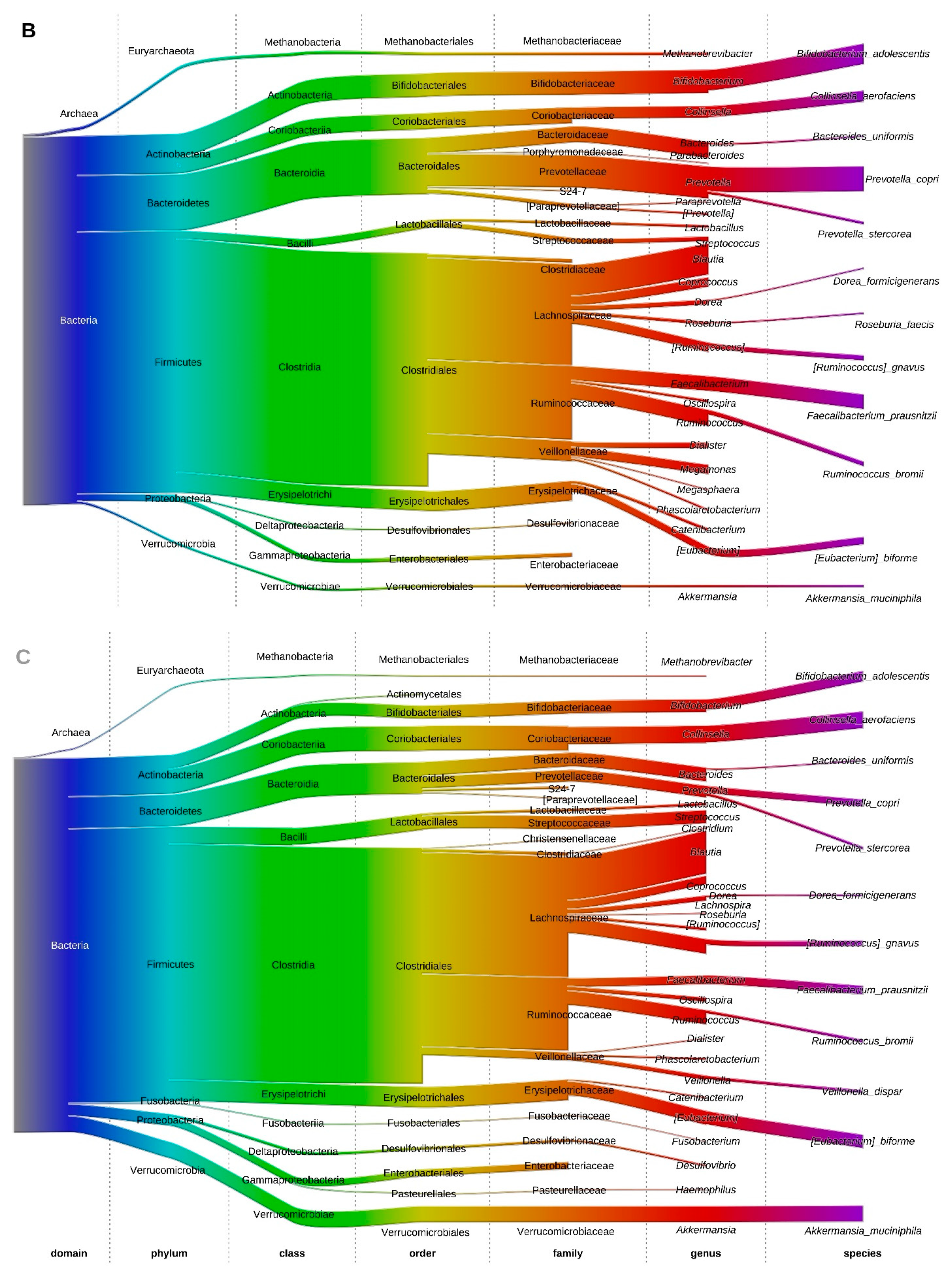
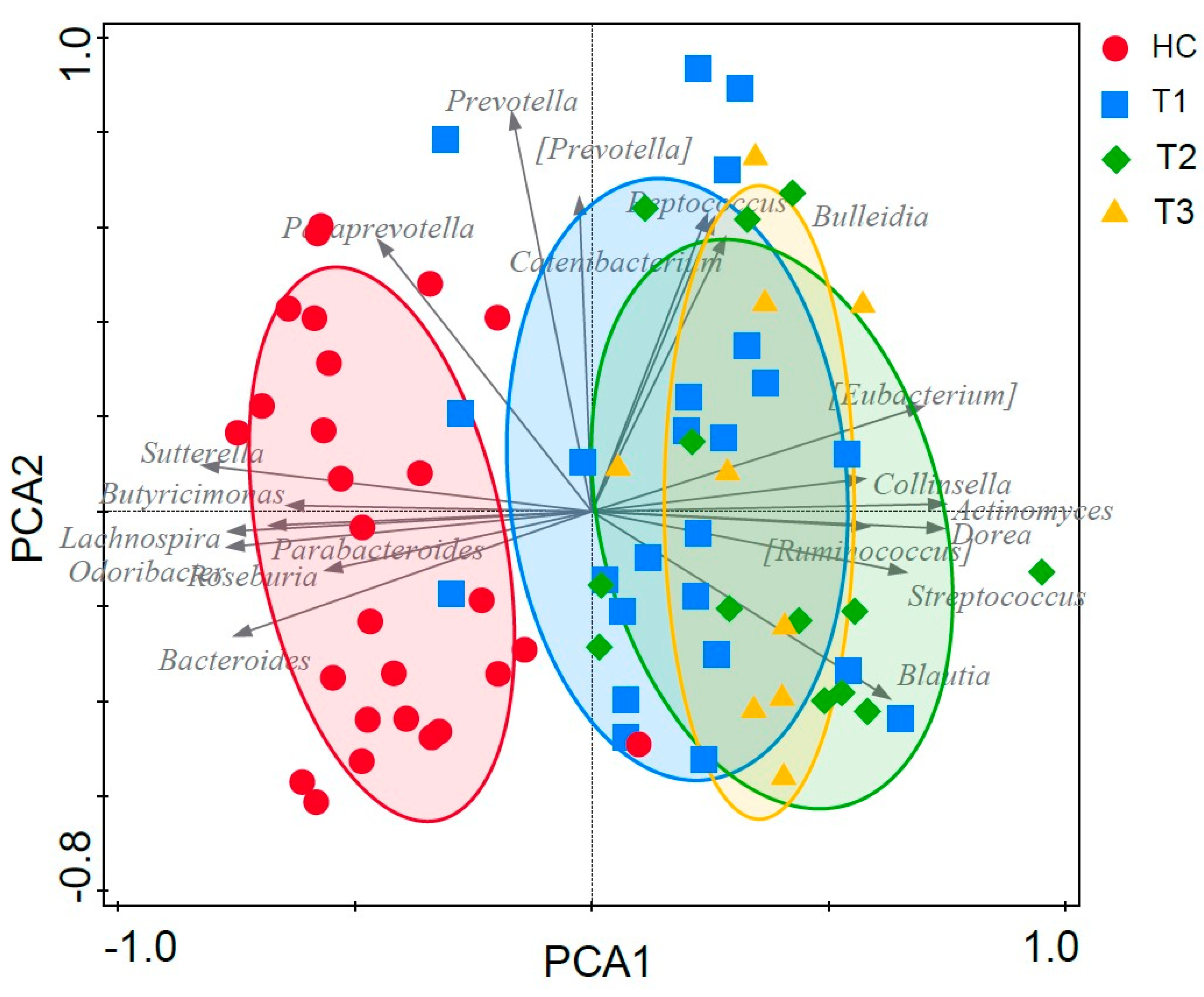
3.6. Fecal Microbiota Structure of Obese and Lean Individuals
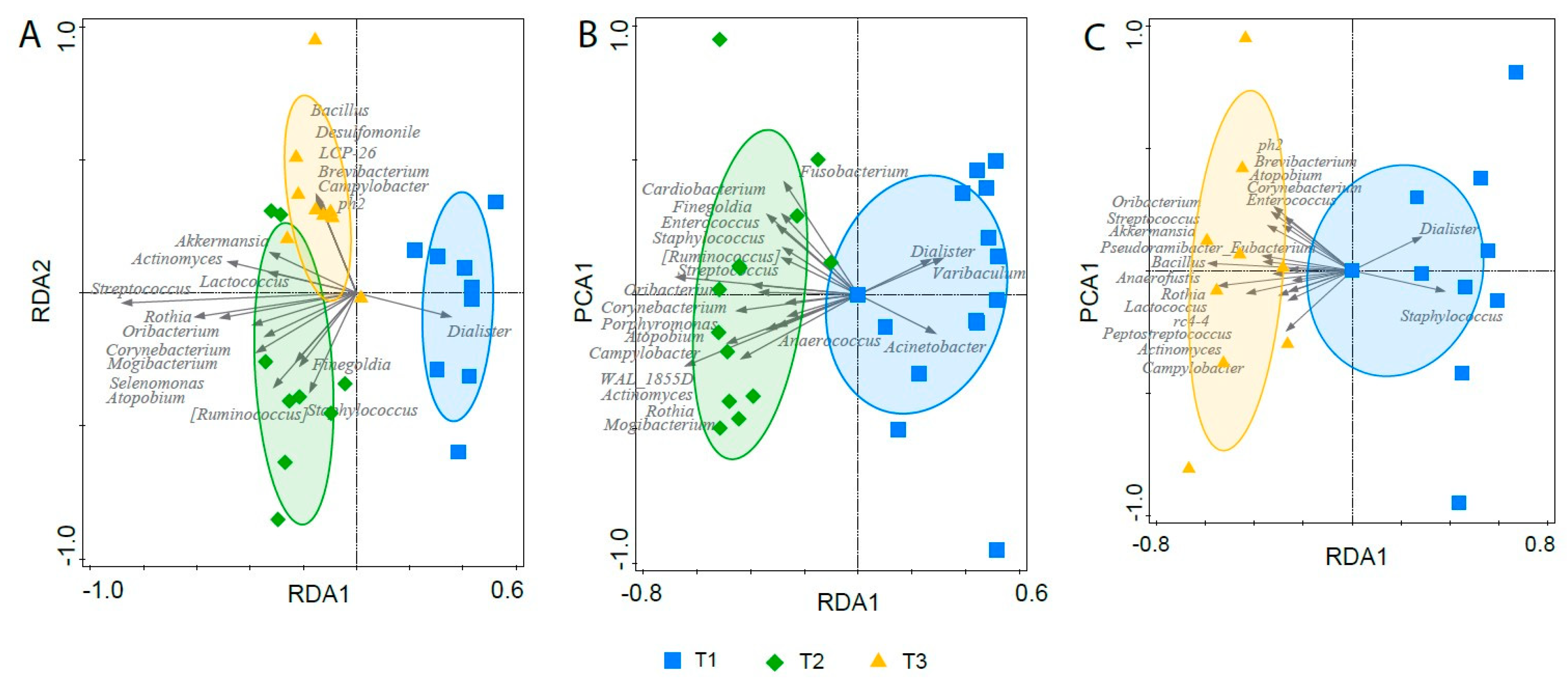
3.7. Effect of Gastric Sleeve Procedure on the Fecal Microbiota in Morbidly Obese Individuals
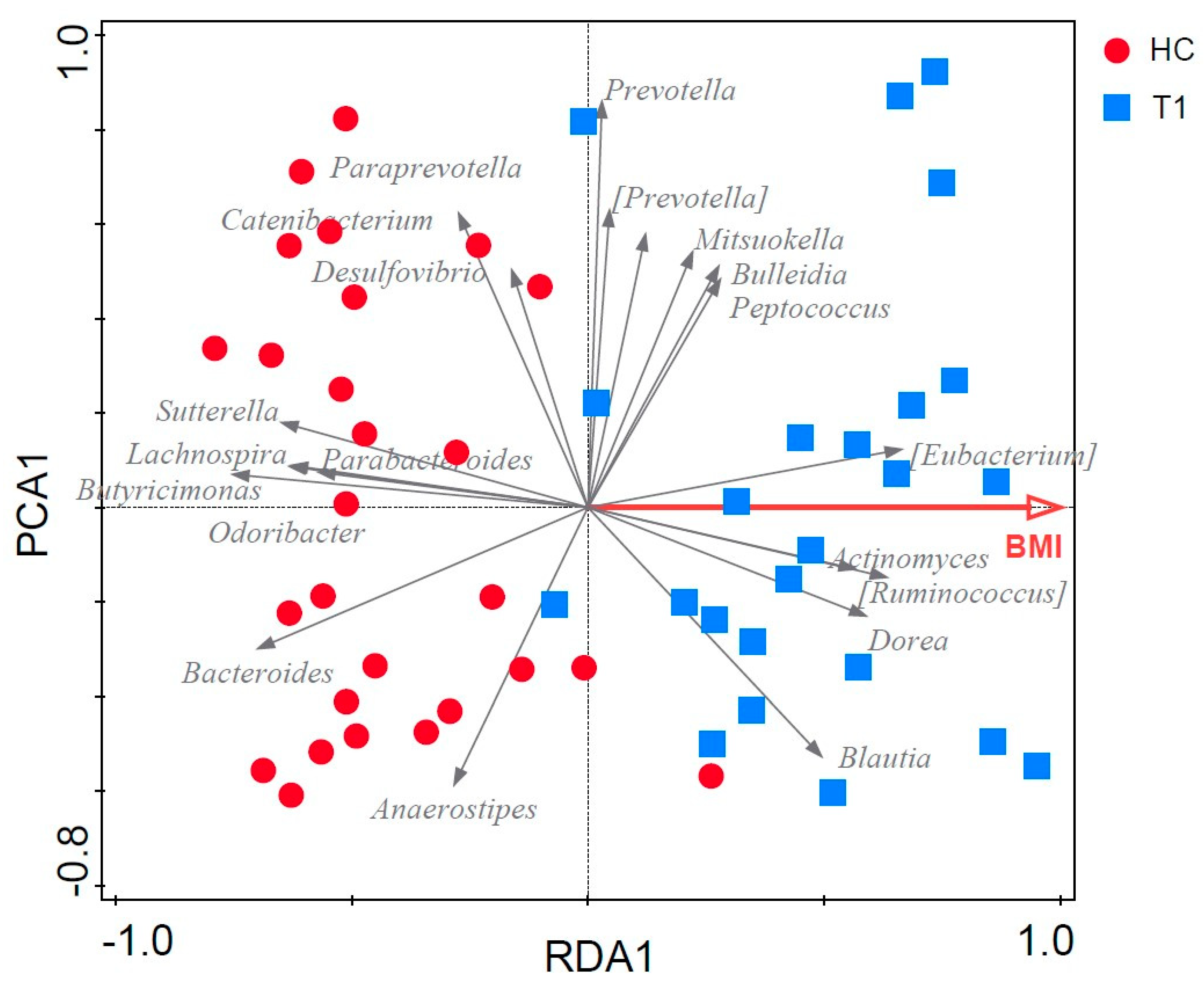
3.8. Potential Confounding Effects on the Microbiota
3.9. Associations between Fecal Microbiota Composition and Biomarkers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swinburn, B.A.; Sacks, G.; Hall, K.D.; McPherson, K.; Finegood, D.T.; Moodie, M.L.; Gortmaker, S.L. The global obesity pandemic: Shaped by global drivers and local environments. Lancet 2011, 378, 804–814. [Google Scholar] [CrossRef]
- Bellanger, T.M.; Bray, G.A. Obesity related morbidity and mortality. J. State Med. Soc. 2005, 157, S42–S49; quiz 49. [Google Scholar]
- Wang, Y.C.; McPherson, K.; Marsh, T.; Gortmaker, S.L.; Brown, M. Health and economic burden of the projected obesity trends in the USA and the UK. Lancet 2011, 378, 815–825. [Google Scholar] [CrossRef]
- Gerard, P. Gut microbiota and obesity. Cell Mol. Life Sci. 2016, 73, 147–162. [Google Scholar] [CrossRef]
- Zhao, L. The gut microbiota and obesity: From correlation to causality. Nat. Rev. Microbiol. 2013, 11, 639–647. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Cheng, J.; Duncan, A.E.; Kau, A.L.; Griffin, N.W.; Lombard, V.; Henrissat, B.; Bain, J.R.; et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 2013, 341, 1241214. [Google Scholar] [CrossRef]
- Armougom, F.; Henry, M.; Vialettes, B.; Raccah, D.; Raoult, D. Monitoring bacterial community of human gut microbiota reveals an increase in Lactobacillus in obese patients and Methanogens in anorexic patients. PLoS ONE 2009, 4, e7125. [Google Scholar] [CrossRef]
- Furet, J.P.; Kong, L.C.; Tap, J.; Poitou, C.; Basdevant, A.; Bouillot, J.L.; Mariat, D.; Corthier, G.; Dore, J.; Henegar, C.; et al. Differential adaptation of human gut microbiota to bariatric surgery-induced weight loss: Links with metabolic and low-grade inflammation markers. Diabetes 2010, 59, 3049–3057. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Arora, T.; Backhed, F. The gut microbiota and metabolic disease: Current understanding and future perspectives. J. Intern. Med. 2016, 280, 339–349. [Google Scholar] [CrossRef]
- Schwiertz, A.; Taras, D.; Schafer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef]
- Jumpertz, R.; Le, D.S.; Turnbaugh, P.J.; Trinidad, C.; Bogardus, C.; Gordon, J.I.; Krakoff, J. Energy-balance studies reveal associations between gut microbes, caloric load, and nutrient absorption in humans. Am. J. Clin. Nutr. 2011, 94, 58–65. [Google Scholar] [CrossRef]
- Duncan, S.H.; Lobley, G.E.; Holtrop, G.; Ince, J.; Johnstone, A.M.; Louis, P.; Flint, H.J. Human colonic microbiota associated with diet, obesity and weight loss. Int. J. Obes. 2008, 32, 1720–1724. [Google Scholar] [CrossRef]
- Graham, C.; Mullen, A.; Whelan, K. Obesity and the gastrointestinal microbiota: A review of associations and mechanisms. Nutr. Rev. 2015, 73, 376–385. [Google Scholar] [CrossRef]
- Tehrani, A.B.; Nezami, B.G.; Gewirtz, A.; Srinivasan, S. Obesity and its associated disease: A role for microbiota? Neurogastroenterol. Motil. 2012, 24, 305–311. [Google Scholar] [CrossRef]
- Woting, A.; Blaut, M. The Intestinal Microbiota in Metabolic Disease. Nutrients 2016, 8, 202. [Google Scholar] [CrossRef]
- Caricilli, A.M.; Saad, M.J.A. The role of gut microbiota on insulin resistance. Nutrients 2013, 5, 829–851. [Google Scholar] [CrossRef]
- Gkolfakis, P.; Dimitriadis, G.; Triantafyllou, K. Gut microbiota and non-alcoholic fatty liver disease. Hepatobiliary Pancreat. Dis. Int. 2015, 14, 572–581. [Google Scholar] [CrossRef]
- Teixeira, T.F.S.; Collado, M.C.; Ferreira, C.L.L.F.; Bressan, J.; Peluzio, M.d.C.G. Potential mechanisms for the emerging link between obesity and increased intestinal permeability. Nutr. Res. 2012, 32, 637–647. [Google Scholar] [CrossRef]
- Kral, J.G.; Kava, R.A.; Catalano, P.M.; Moore, B.J. Severe obesity: The neglected epidemic. Obes. Facts 2012, 5, 254–269. [Google Scholar] [CrossRef]
- Mosinski, J.D.; Pagadala, M.R.; Mulya, A.; Huang, H.; Dan, O.; Shimizu, H.; Batayyah, E.; Pai, R.K.; Schauer, P.R.; Brethauer, S.A.; et al. Gastric bypass surgery is protective from high-fat diet-induced non-alcoholic fatty liver disease and hepatic endoplasmic reticulum stress. Acta Physiol. 2016, 217, 141–151. [Google Scholar] [CrossRef]
- Sjostrom, L.; Peltonen, M.; Jacobson, P.; Sjostrom, C.D.; Karason, K.; Wedel, H.; Ahlin, S.; Anveden, A.; Bengtsson, C.; Bergmark, G.; et al. Bariatric surgery and long-term cardiovascular events. JAMA 2012, 307, 56–65. [Google Scholar] [CrossRef]
- Mehaffey, J.H.; LaPar, D.J.; Clement, K.C.; Turrentine, F.E.; Miller, M.S.; Hallowell, P.T.; Schirmer, B.D. 10-Year Outcomes After Roux-en-Y Gastric Bypass. Ann. Surg. 2016, 264, 121–126. [Google Scholar] [CrossRef]
- Shi, X.; Karmali, S.; Sharma, A.M.; Birch, D.W. A Review of Laparoscopic Sleeve Gastrectomy for Morbid Obesity. Obes. Surg. 2010, 20, 1171–1177. [Google Scholar] [CrossRef]
- Debedat, J.; Clement, K.; Aron-Wisnewsky, J. Gut Microbiota Dysbiosis in Human Obesity: Impact of Bariatric Surgery. Curr. Obes. Rep. 2019, 8, 229–242. [Google Scholar] [CrossRef]
- Gutierrez-Repiso, C.; Garrido-Sanchez, L.; Alcaide-Torres, J.; Cornejo-Pareja, I.; Ocana-Wilhelmi, L.; Garcia-Fuentes, E.; Moreno-Indias, I.; Tinahones, F.J. Predictive Role of Gut Microbiota in Weight Loss Achievement after Bariatric Surgery. J. Am. Coll. Surg. 2022, 234, 861–871. [Google Scholar] [CrossRef]
- Morales-Marroquin, E.; Hanson, B.; Greathouse, L.; de la Cruz-Munoz, N.; Messiah, S.E. Comparison of methodological approaches to human gut microbiota changes in response to metabolic and bariatric surgery: A systematic review. Obes. Rev. 2020, 21, e13025. [Google Scholar] [CrossRef]
- Wang, F.G.; Bai, R.X.; Yan, W.M.; Yan, M.; Dong, L.Y.; Song, M.M. Differential composition of gut microbiota among healthy volunteers, morbidly obese patients and post-bariatric surgery patients. Exp. Ther. Med. 2019, 17, 2268–2278. [Google Scholar] [CrossRef]
- Kellerer, T.; Brandl, B.; Buttner, J.; Lagkouvardos, I.; Hauner, H.; Skurk, T. Impact of Laparoscopic Sleeve Gastrectomy on Gut Permeability in Morbidly Obese Subjects. Obes. Surg. 2019, 29, 2132–2143. [Google Scholar] [CrossRef]
- Palmisano, S.; Campisciano, G.; Silvestri, M.; Guerra, M.; Giuricin, M.; Casagranda, B.; Comar, M.; de Manzini, N. Changes in Gut Microbiota Composition after Bariatric Surgery: A New Balance to Decode. J. Gastrointest. Surg. 2020, 24, 1736–1746. [Google Scholar] [CrossRef] [PubMed]
- Paganelli, F.L.; Luyer, M.; Hazelbag, C.M.; Uh, H.W.; Rogers, M.R.C.; Adriaans, D.; Berbers, R.M.; Hendrickx, A.P.A.; Viveen, M.C.; Groot, J.A.; et al. Roux-Y Gastric Bypass and Sleeve Gastrectomy directly change gut microbiota composition independent of surgery type. Sci. Rep. 2019, 9, 10979. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Caixas, A.; Ahlers, M.; Patel, K.; Gao, Z.; Dutia, R.; Blaser, M.J.; Clemente, J.C.; Laferrere, B. Longitudinal changes of microbiome composition and microbial metabolomics after surgical weight loss in individuals with obesity. Surg. Obes. Relat. Dis. 2019, 15, 1367–1373. [Google Scholar] [CrossRef]
- van Wijck, K.; Verlinden, T.J.; van Eijk, H.M.; Dekker, J.; Buurman, W.A.; Dejong, C.H.; Lenaerts, K. Novel multi-sugar assay for site-specific gastrointestinal permeability analysis: A randomized controlled crossover trial. Clin. Nutr. 2013, 32, 245–251. [Google Scholar] [CrossRef]
- Wilbrink, J.; Bernards, N.; Mujagic, Z.; van Avesaat, M.; Pijls, K.; Klaassen, T.; van Eijk, H.; Nienhuijs, S.; Stronkhorst, A.; Wilms, E.; et al. Intestinal barrier function in morbid obesity: Results of a prospective study on the effect of sleeve gastrectomy. Int J Obes. 2020, 44, 368–376. [Google Scholar] [CrossRef]
- Mujagic, Z.; Ludidi, S.; Keszthelyi, D.; Hesselink, M.A.; Kruimel, J.W.; Lenaerts, K.; Hanssen, N.M.; Conchillo, J.M.; Jonkers, D.M.; Masclee, A.A. Small intestinal permeability is increased in diarrhoea predominant IBS, while alterations in gastroduodenal permeability in all IBS subtypes are largely attributable to confounders. Aliment. Pharmacol. Ther. 2014, 40, 288–297. [Google Scholar] [CrossRef]
- Pijls, K.E.; Koek, G.H.; Elamin, E.E.; de Vries, H.; Masclee, A.A.; Jonkers, D.M. Large intestine permeability is increased in patients with compensated liver cirrhosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G147–G153. [Google Scholar] [CrossRef]
- Koning, C.J.; Jonkers, D.M.; Stobberingh, E.E.; Mulder, L.; Rombouts, F.M.; Stockbrugger, R.W. The effect of a multispecies probiotic on the intestinal microbiota and bowel movements in healthy volunteers taking the antibiotic amoxycillin. Am. J. Gastroenterol. 2008, 103, 178–189. [Google Scholar] [CrossRef]
- Tedjo, D.I.; Jonkers, D.M.; Savelkoul, P.H.; Masclee, A.A.; van Best, N.; Pierik, M.J.; Penders, J. The effect of sampling and storage on the fecal microbiota composition in healthy and diseased subjects. PLoS ONE 2015, 10, e0126685. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Lazarevic, V.; Derrien, M.; Girard, M.; Muccioli, G.G.; Muccioli, G.M.; Neyrinck, A.M.; Possemiers, S.; Van Holle, A.; François, P.; et al. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes 2011, 60, 2775–2786. [Google Scholar] [CrossRef] [PubMed]
- Walters, W.A.; Xu, Z.; Knight, R. Meta-analyses of human gut microbes associated with obesity and IBD. FEBS Lett. 2014, 588, 4223–4233. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Fei, N.; Pang, X.; Shen, J.; Wang, L.; Zhang, B.; Zhang, M.; Zhang, X.; Zhang, C.; Li, M.; et al. A gut microbiota-targeted dietary intervention for amelioration of chronic inflammation underlying metabolic syndrome. FEMS Microbiol. Ecol. 2014, 87, 357–367. [Google Scholar] [CrossRef]
- Zhang, H.; DiBaise, J.K.; Zuccolo, A.; Kudrna, D.; Braidotti, M.; Yu, Y.; Parameswaran, P.; Crowell, M.D.; Wing, R.; Rittmann, B.E.; et al. Human gut microbiota in obesity and after gastric bypass. Proc. Natl. Acad. Sci. USA 2009, 106, 2365–2370. [Google Scholar] [CrossRef]
- Fernandes, J.; Su, W.; Rahat-Rozenbloom, S.; Wolever, T.M.; Comelli, E.M. Adiposity, gut microbiota and faecal short chain fatty acids are linked in adult humans. Nutr. Diabetes 2014, 4, e121. [Google Scholar] [CrossRef]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef]
- Teixeira, T.F.; Grzeskowiak, L.; Franceschini, S.C.; Bressan, J.; Ferreira, C.L.; Peluzio, M.C. Higher level of faecal SCFA in women correlates with metabolic syndrome risk factors. Br. J. Nutr. 2013, 109, 914–919. [Google Scholar] [CrossRef]
- Remely, M.; Tesar, I.; Hippe, B.; Gnauer, S.; Rust, P.; Haslberger, A.G. Gut microbiota composition correlates with changes in body fat content due to weight loss. Benef. Microbes 2015, 6, 431–439. [Google Scholar] [CrossRef]
- Lee, H.; Ko, G. Effect of Metformin on Metabolic Improvement and Gut Microbiota. Appl. Environ. Microbiol. 2014, 80, 5935–5943. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, T.; Aida, M.; Yoshida, Y.; Matsumoto, S.; Tanaka, M.; Nakayama, J.; Nagao, Y.; Nakata, R.; Oki, E.; Akahoshi, T.; et al. Alteration in faecal bile acids, gut microbial composition and diversity after laparoscopic sleeve gastrectomy. Br. J. Surg. 2020, 107, 1673–1685. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zhuang, J.; Cui, Q.; Jiang, S.; Tao, W.; Chen, W.; Yu, S.; Wu, L.; Yang, W.; Liu, F.; et al. Two Bariatric Surgical Procedures Differentially Alter the Intestinal Microbiota in Obesity Patients. Obes. Surg. 2020, 30, 2345–2361. [Google Scholar] [CrossRef] [PubMed]
- Dao, M.C.; Everard, A.; Clément, K.; Cani, P.D. Losing weight for a better health: Role for the gut microbiota. Clin. Nutr. Exp. 2016, 6, 39–58. [Google Scholar] [CrossRef]
- van den Bogert, B.; Erkus, O.; Boekhorst, J.; de Goffau, M.; Smid, E.J.; Zoetendal, E.G.; Kleerebezem, M. Diversity of human small intestinal Streptococcus and Veillonella populations. FEMS Microbiol. Ecol. 2013, 85, 376–388. [Google Scholar] [CrossRef]
- Aron-Wisnewsky, J.; Dore, J.; Clement, K. The importance of the gut microbiota after bariatric surgery. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 590–598. [Google Scholar] [CrossRef]
- Braghetto, I.; Davanzo, C.; Korn, O.; Csendes, A.; Valladares, H.; Herrera, E.; Gonzalez, P.; Papapietro, K. Scintigraphic evaluation of gastric emptying in obese patients submitted to sleeve gastrectomy compared to normal subjects. Obes. Surg. 2009, 19, 1515–1521. [Google Scholar] [CrossRef]
- Damms-Machado, A.; Mitra, S.; Schollenberger, A.E.; Kramer, K.M.; Meile, T.; Königsrainer, A.; Huson, D.H.; Bischoff, S.C. Effects of surgical and dietary weight loss therapy for obesity on gut microbiota composition and nutrient absorption. BioMed Res. Int. 2015, 2015, 806248. [Google Scholar] [CrossRef]
- Steenackers, N.; Wauters, L.; Van der Schueren, B.; Augustijns, P.; Falony, G.; Koziolek, M.; Lannoo, M.; Mertens, A.; Meulemans, A.; Raes, J.; et al. Effect of obesity on gastrointestinal transit, pressure and pH using a wireless motility capsule. Eur. J. Pharm. Biopharm. 2021, 167, 1–8. [Google Scholar] [CrossRef]
- Verdam, F.J.; Fuentes, S.; de Jonge, C.; Zoetendal, E.G.; Erbil, R.; Greve, J.W.; Buurman, W.A.; de Vos, W.M.; Rensen, S.S. Human intestinal microbiota composition is associated with local and systemic inflammation in obesity. Obesity 2013, 21, E607–E615. [Google Scholar] [CrossRef]
- Brignardello, J.; Morales, P.; Diaz, E.; Romero, J.; Brunser, O.; Gotteland, M. Pilot study: Alterations of intestinal microbiota in obese humans are not associated with colonic inflammation or disturbances of barrier function. Aliment. Pharmacol. Ther. 2010, 32, 1307–1314. [Google Scholar] [CrossRef] [PubMed]
- Carswell, K.A.; Vincent, R.P.; Belgaumkar, A.P.; Sherwood, R.A.; Amiel, S.A.; Patel, A.G.; le Roux, C.W. The effect of bariatric surgery on intestinal absorption and transit time. Obes. Surg. 2014, 24, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Di Palo, D.M.; Garruti, G.; Di Ciaula, A.; Molina-Molina, E.; Shanmugam, H.; De Angelis, M.; Portincasa, P. Increased Colonic Permeability and Lifestyles as Contributing Factors to Obesity and Liver Steatosis. Nutrients 2020, 12, 564. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, T.F.; Souza, N.C.; Chiarello, P.G.; Franceschini, S.C.; Bressan, J.; Ferreira, C.L.; Peluzio Mdo, C. Intestinal permeability parameters in obese patients are correlated with metabolic syndrome risk factors. Clin. Nutr. 2012, 31, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Ciobarca, D.; Catoi, A.F.; Copaescu, C.; Miere, D.; Crisan, G. Bariatric Surgery in Obesity: Effects on Gut Microbiota and Micronutrient Status. Nutrients 2020, 12, 235. [Google Scholar] [CrossRef] [PubMed]
- Anhe, F.F.; Varin, T.V.; Schertzer, J.D.; Marette, A. The Gut Microbiota as a Mediator of Metabolic Benefits after Bariatric Surgery. Can. J. Diabetes 2017, 41, 439–447. [Google Scholar] [CrossRef]
- Tu, J.; Wang, Y.; Jin, L.; Huang, W. Bile acids, gut microbiota and metabolic surgery. Front. Endocrinol. 2022, 13, 929530. [Google Scholar] [CrossRef]
- Winston, J.A.; Rivera, A.; Cai, J.; Patterson, A.D.; Theriot, C.M. Secondary bile acid ursodeoxycholic acid alters weight, the gut microbiota, and the bile acid pool in conventional mice. PLoS ONE 2021, 16, e0246161. [Google Scholar] [CrossRef]
- Winston, J.A.; Theriot, C.M. Diversification of host bile acids by members of the gut microbiota. Gut Microbes 2020, 11, 158–171. [Google Scholar] [CrossRef]
- Goodrich, J.K.; Di Rienzi, S.C.; Poole, A.C.; Koren, O.; Walters, W.A.; Caporaso, J.G.; Knight, R.; Ley, R.E. Conducting a microbiome study. Cell 2014, 158, 250–262. [Google Scholar] [CrossRef]
- Conlon, M.A.; Bird, A.R. The impact of diet and lifestyle on gut microbiota and human health. Nutrients 2015, 7, 17–44. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lean Subjects (N = 27) | Subjects with Obesity T1 (N = 24) pre-GS | T2 (N = 13) Two Months Post-GS | T3 (N = 9) Six Months Post-GS | |
|---|---|---|---|---|
| BMI (kg/m2) (median, IQR) | 23.0 (21.5–24.4) * | 43.5 (39.4–48.3) *$ | 37.9 (36.0–43.0) *$ | 34.8 (31.5–39.4) *$ |
| Total weight loss (%) (median, IQR) | Not applicable | Not applicable | 14.8 (11.8–20.4) | 25.1 (21.4–27.4) |
| Age (yrs) (median, IQR) | 47 (24–61) | 43 (37–55) | 50 (41.5–55) | 50.0 (42–55) |
| Male, number (%) | 12 (44.4) | 11 (45.8) | 7 (53.8) | 5 (55.6) |
| Current smokers, n (%) | 3 (11.1) | 7 (29.2) | 3 (23.1) | 3 (33.3) |
| Hypertension n (%) | 0 (0) # | 14 (58.3) # | 9 (69.2) # | 5 (55.6) # |
| Type 2 diabetes (n, %) | 0 (0) # | 10 (41.7) # | 6 (46.2) # | 4 (44.4) # |
| Medication (n, %) | ||||
| Insulin | 0 (0) # | 4 (16.7)# | 3 (23.1) | 0 (0) |
| Oral antidiabetics 1 | 0 (0) | 13 (54.1) | 7 (53.8) | 8 (88.9) |
| PPI use (%) 2 | 1 (3.7) | 2 (3.8) | 3 (23.1) | 1 (11.1) |
| Antibiotics 3 | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Past antibiotics 4 | 1 (3.7) # | 0 (0) | 0 (0) | 1 (11.1) |
| Plasma markers | ||||
| hsCRP (mg/l, median, IQR) | 0.6 (0.4–0.9) * | 6.3 (3.0–4.0) *$ | 7.3 (5.4–11.2) *& | 3.4 (1.9–4.1) *$& |
| Hba1c (mM Hba1c/M HB median, IQR) | Not available | 49 (40–73) $ | 41 (35–55) $ | 43 (33–53) $ |
| Fecal markers | ||||
| Calprotectin (µg/gram feces, median, IQR) | 1.4 (0.8–3.4) * | 3.7 (2.0–9.7) *$& | 15.6 (11.9–21.0) *$ | 10.7 (8.6–13.7) *& |
| Protein content (µg protein/mg feces, median, IQR) | 117.4 (92.2–192.5) | 127.0 (68.5–200.6) | 162.6 (52.7–237.6) | 162.2 (101.2–231.4) |
| Urinary markers | ||||
| Sucrose (µg, median, IQR) | 1958.8 (1042.8–3716.6) * | 5642.1 (1956.3–24,273.7) * | 1846.2 (450.7–4565.1) | 2758.5 (1174.0–5694.4) |
| Lactulose/rhamnose (median, IQR) | 0.04 (0.03–0.08) | 0.05 (0.02–0.07) $ | 0.05 (0.03–0.06) & | 0.045 (0.04–0.10) $& |
| Sucralose/erythritol (median, IQR) | 0.06 (0.02–0.08) | 0.05 (0.02–0.06) $ | 0.08 (0.03–0.09) $ | 0.04 (0.02–0.06) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tedjo, D.I.; Wilbrink, J.A.; Boekhorst, J.; Timmerman, H.M.; Nienhuijs, S.W.; Stronkhorst, A.; Savelkoul, P.H.M.; Masclee, A.A.M.; Penders, J.; Jonkers, D.M.A.E. Impact of Sleeve Gastrectomy on Fecal Microbiota in Individuals with Morbid Obesity. Microorganisms 2023, 11, 2353. https://doi.org/10.3390/microorganisms11092353
Tedjo DI, Wilbrink JA, Boekhorst J, Timmerman HM, Nienhuijs SW, Stronkhorst A, Savelkoul PHM, Masclee AAM, Penders J, Jonkers DMAE. Impact of Sleeve Gastrectomy on Fecal Microbiota in Individuals with Morbid Obesity. Microorganisms. 2023; 11(9):2353. https://doi.org/10.3390/microorganisms11092353
Chicago/Turabian StyleTedjo, Danyta I., Jennifer A. Wilbrink, Jos Boekhorst, Harro M. Timmerman, Simon W. Nienhuijs, Arnold Stronkhorst, Paul H. M. Savelkoul, Ad A. M. Masclee, John Penders, and Daisy M. A. E. Jonkers. 2023. "Impact of Sleeve Gastrectomy on Fecal Microbiota in Individuals with Morbid Obesity" Microorganisms 11, no. 9: 2353. https://doi.org/10.3390/microorganisms11092353