
Characterization of the Mycoremediation of n-Alkanes and Branched-Chain Alkanes by Filamentous Fungi from Oil-Polluted Soil Samples in Kazakhstan
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Cultures
2.2. Substrates
2.3. Strain Identification
2.3.1. Microscopy
2.3.2. Cell Lysis and Polymerase Chain Reaction (PCR)
2.4. Growth Experiment on Substrates
2.5. Biotransformation
2.5.1. Cultivation
2.5.2. Incubation
2.5.3. Determination of Dry Weight
2.6. Liquid–Liquid Extraction
2.7. Methylation and Gas Chromatography–Mass Spectrometry (GC-MS)
3. Results
3.1. Identification of the Examined Fungal Strains
3.2. Ability of the Tested Fungi to Use Oil Components as Growth Substrates
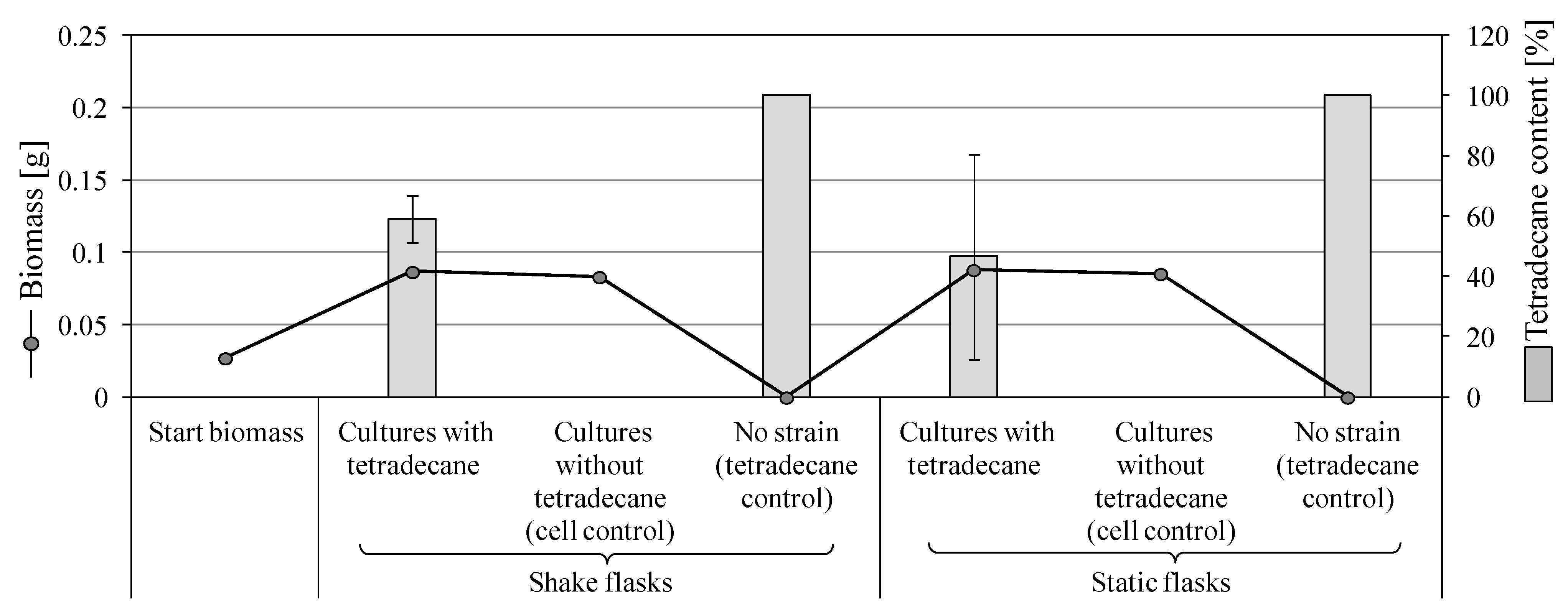
3.2.1. Growth on Tetradecane
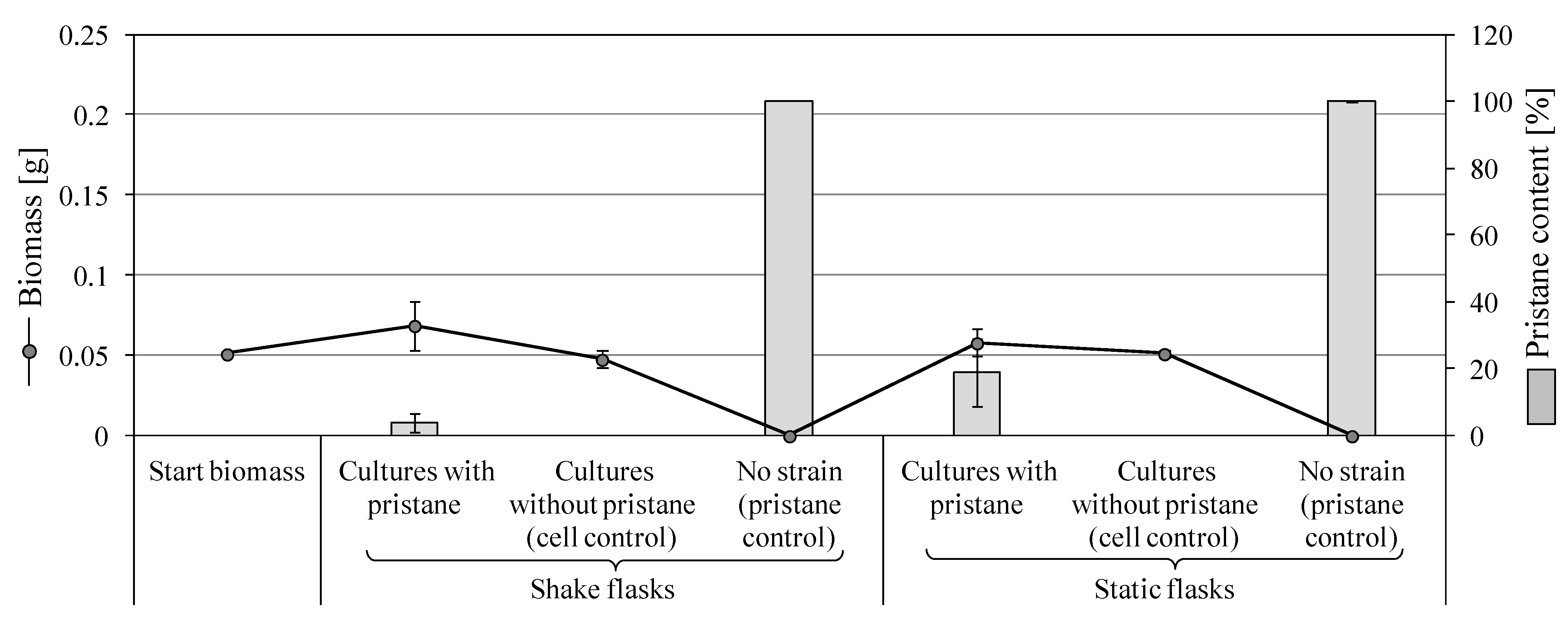
3.2.2. Growth on Pristane
3.3. Biodegradation of Oil Components as Substrates by the Tested Fungal Strains
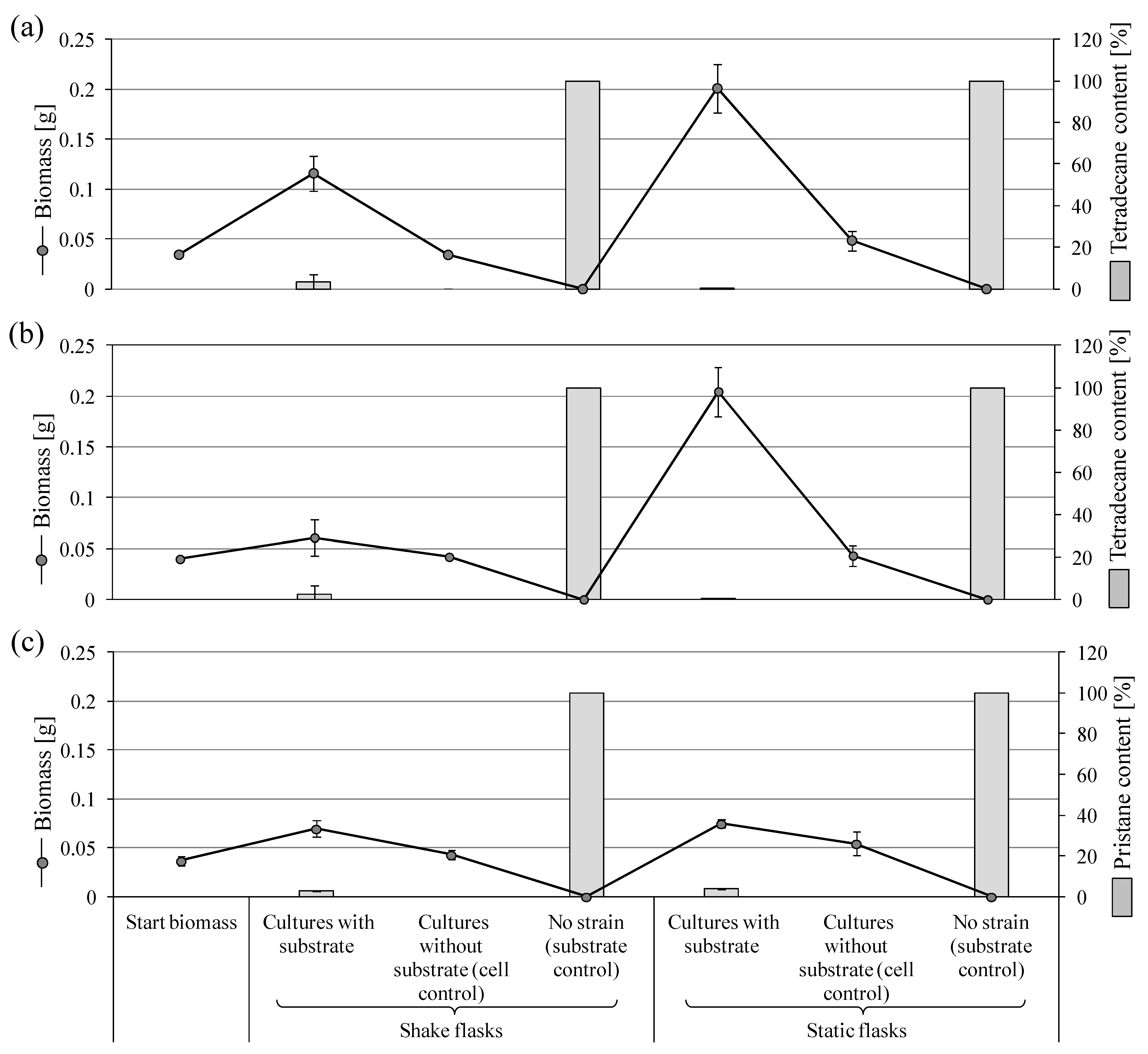
3.3.1. Biodegradation of Tetradecane and Pristane by P. javanicum
3.3.2. Degradation of Tetradecane by S. boydii SBUG-M1749
3.3.3. Degradation of Pristane by F. oxysporum SBUG-M1747
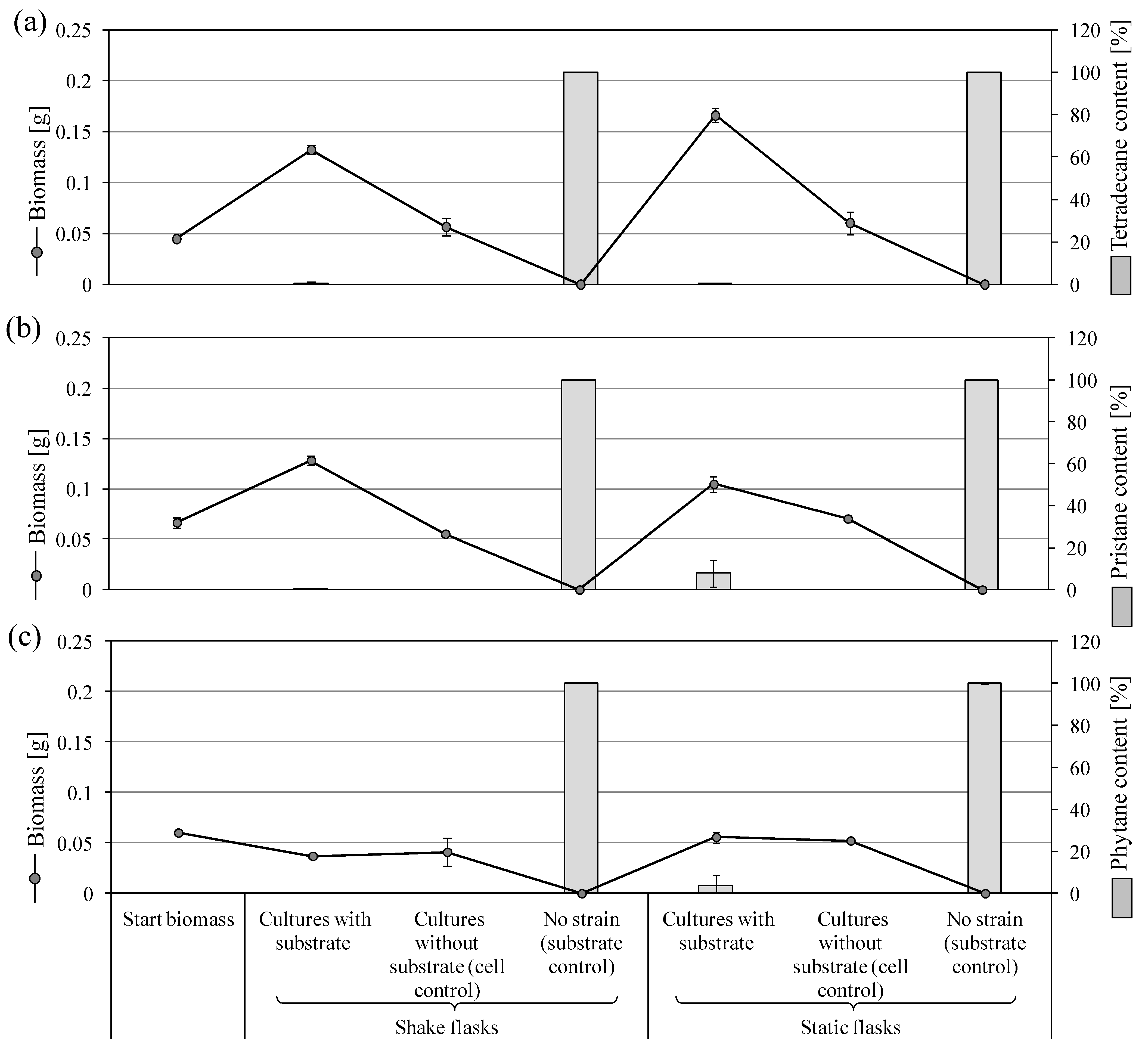
3.3.4. Biodegradation of Tetradecane, Pristane, and Phytane by P. lilacinum SBUG-M1751
4. Discussion
4.1. Degradation Potential of Tetradecane
4.2. Degradation Potential of Pristane
4.3. Degradation Potential of Phytane
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Focht, D.D. Diffusional Constraints on Microbial Processes in Soil. Soil Sci. 1992, 154, 300–307. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Blagodatskaya, E. Microbial Hotspots and hot moments in soil: Concept & review. Soil Biol. Biochem. 2015, 83, 184–199. [Google Scholar] [CrossRef]
- Castorena, E.V.G.; Gutiérrez-Castorena, M.C.; Vargas, T.C.; Bontemps, L.C.; Martínez, J.D.; Méndez, E.S.; Solorio, C.A.O. Micromapping of microbial hotspots and biofilms from different crops using digital image mosaics of soil thin sections. Geoderma 2016, 279, 11–21. [Google Scholar] [CrossRef]
- Dewi, W.S.; Amalina, D.D.; Romadhon, M.R. Microbial Biofilm for Soil Health, Plant Growth, and Productivity under Multi Stress. A Review. IOP Conf. Ser. Earth Environ. Sci. 2023, 1162, 012008. [Google Scholar] [CrossRef]
- Bundesamtes für Seeschifffahrt und Hydrographie (BSH). Öl im Meer—Risiken, Vorsorge und Bekämpfung. 2011; Nr. 48; ISSN 0946-6010. Available online: https://www.bsh.de/DE/PUBLIKATIONEN/_Anlagen/Downloads/Meer_und_Umwelt/Berichte-des-BSH/Berichte-des-BSH_48.html (accessed on 1 July 2023).
- Baruah, P.; Saikia, R.R.; Baruah, P.P.; Deka, S. Effect of crude oil contamination on the chlorophyll content and morpho-anatomy of Cyperus brevifolius (Rottb.) Hassk. Environ. Sci. Pollut. Res. Int. 2014, 21, 12530–12538. [Google Scholar] [CrossRef]
- Joye, S.B. Deepwater Horizon, 5 years on. Science 2015, 349, 592–593. [Google Scholar] [CrossRef]
- Rako, L.K.; Costrut, A.M.; Braun, M.; Bendels, M.H.K. Der Untergang der Deep Water Horizon im Golf von Mexiko. Zbl. Arbeitsmed. 2018, 68, 346–351. [Google Scholar] [CrossRef]
- World Trade Organization (WTO) Annual Report. 2017. Available online: www.wto.org (accessed on 1 July 2023).
- Bekmurzaev, N. Kazakhstan Exports Oil to Germany as Russia Keeps a Close Eye. Eurasia Dly. Monit. 2023, 20. Available online: https://jamestown.org/program/kazakhstan-exports-oil-to-germany-as-russia-keeps-a-close-eye/ (accessed on 1 July 2023).
- German Federal Institute for Risk Assessment (BFR) 2016. Available online: https://www.bfr.bund.de/de/bfr_wissenschaft_2016.html (accessed on 1 July 2023).
- Rüffer, M. Ölsperren für Feuerwehren: Praxistipps. Available online: https://www.feuerwehrmagazin.de/wissen/oelsperren-tipps-fuer-diebeschaffung-und-den-einsatz-54054 (accessed on 13 July 2023).
- Mullin, J.V.; Champ, M.A. Introduction/Overview to In Situ Burning of oil spills. Spill Sci. Technol. Bull. 2003, 8, 323–330. [Google Scholar] [CrossRef]
- Little, D.I.; Sheppard, S.R.J.; Hulme, D. A perspective on oil spills: What we should have learned about global warming. Ocean. Coast. Manag. 2021, 202, 105509. [Google Scholar] [CrossRef]
- Oil spills: Microorganisms to the rescue? Nat. Rev. Microbiol. 2010, 8, 462. [CrossRef]
- Van Beilen, J.B.; Funhoff, E.G. Alkane hydroxylases involved in microbial alkane degradation. Appl. Microbiol. Biotechnol. 2007, 74, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Rojo, F. Degradation of alkanes by bacteria. Environ. Microbiol. 2009, 11, 2477–2490. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Gan, L.; Chen, Z.; Naidu, R. Biodegradation of tetradecane using Acinetobacter venetianus immobilized on bagasse. Biochem. Eng. J. 2015, 100, 76–82. [Google Scholar] [CrossRef]
- Ali, N.; Khanafer, M.; Al-Awadhi, H. Indigenous oil-degrading bacteria more efficient in soil bioremediation than microbial consortium and active even in super oil-saturated soils. Front. Microbiol. 2022, 13, 950051. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, G. Allgemeine Mikrobiologie; Georg Thieme Verlag: Stuttgart, Germany, 2017; ISBN 978-3-13-241885-1. Available online: https://eref.thieme.de/ebooks/1851823#/ebook_1851823_SL72320860 (accessed on 10 July 2023).
- Al-Hawash, A.B.; Alkooranee, J.T.; Abbood, H.A.; Zhang, J.; Sun, J.; Zhang, Y.; Ma, F. Isolation and characterization of two crude oil-degrading fungi strains from Rumaila oil field, Iraq. Biotechnol. Rep. 2018, 17, 104–109. [Google Scholar] [CrossRef]
- Daâssi, D.; Almaghrabi, F.Q. Petroleum-Degrading Fungal Isolates for the Treatment of Soil Microcosms. Microorganisms 2023, 11, 1351. [Google Scholar] [CrossRef]
- Vaksmaa, A.; Guerrero-Cruz, S.; Ghosh, P.; Zeghal, E.; Hernando-Morales, V.; Niemann, H. Role of fungi in bioremediation of emerging pollutants. Front. Mar. Sci. 2023, 10, 1070905. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, J.; Liu, Y.; Fen, W. Biodegradation of hydrocarbons by Purpureocillium lilacinum and Penicillium chrysogenum from heavy oil sludge and their potential for bioremediation of contaminated soils. Int. Biodeterior. Biodegrad. 2023, 178, 105566. [Google Scholar] [CrossRef]
- Schmitz, A. Kasachstan: Neue Führungsmacht im postsowjetischen Raum? Stiftung Wissenschaft und Politik: Berlin, Germany, 2009; pp. 1–30. Available online: https://www.swp-berlin.org/publikation/kasachstan/ (accessed on 10 July 2023).
- Dahl, C.; Kuralbayeva, K. Energy and the environment in Kazakhstan. Energy Policy 2001, 29, 429–440. [Google Scholar] [CrossRef]
- Müller, F. Mikrobielle Diversität von Kasachischen Schadstoffbelasteten Böden und Ihr Potential Hinsichtlich des Abbaus von Erdölbestandteilen. Master‘s Thesis, Universität Greifswald, Greifswald, Germany, 27 November 2018. [Google Scholar]
- Hornei, S.; Köhler, M.; Weide, H. Das Fettsäurespektrum eines Candida Stammes nach Kultur auf n-Alkanen. Z. Mikrobiol. 1972, 12, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, W. Der Einfluß der Kohlenstoffquelle auf das Wachstum, den Proteingehalt und das Enzymmuster. Z. Allg. Mikrobiol. 1968, 8, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Van der Walt, J.P.; van Kerken, A.E. The wine yeast of the cape: Part V.—Studies on the occurrence of Brettanomyces intermedius and Brettanomyces schanderlii. Antonie Van Leeuwenhoek 1961, 27, 81–90. [Google Scholar] [CrossRef]
- Mikolasch, A.; Donath, M.; Reinhard, A.; Herzer, C.; Zayadan, B.; Urich, T.; Schauer, F. Diversity and degradative capabilities of bacteria and fungi isolated from oil-contaminated and hydrocarbon-polluted soils in Kazakhstan. Appl. Microbiol. Biotechnol. 2019, 103, 7261–7274. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols. A Guide to Methods and Applications; Innis, M.A., Ed.; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Hadziavdic, K.; Lekang, K.; Lanzen, A.; Jonassen, I.; Thompson, E.M.; Troedsson, C. Characterization of the 18S RRNA Gene for Designing Universal Eukaryote Specific Primers. PLoS ONE 2014, 9, e87624. [Google Scholar] [CrossRef]
- Ray, J.L.; Althammer, J.; Skaar, K.S.; Simonelli, P.; Larsen, A.; Stoecker, D.; Sazhin, A.; Ijaz, U.Z.; Quince, C.; Nejstgaard, J.C.; et al. Metabarcoding and Metabolome Analyses of Copepod Grazing Reveal Feeding Preference and Linkage to Metabolite Classes in Dynamic Microbial Plankton Communities. Mol. Ecol. 2016, 25, 5585–5602. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Das, N.; Chandran, P. Microbial degradation of petroleum hydrocarbon contaminants: An overview. Biotechnol. Res. Int. 2011, 2011, 941810. [Google Scholar] [CrossRef]
- Rudakiya, D.M.; Tripathi, A.; Gupte, S.G.; Gupte, A. Fungal bioremediation: A step towards cleaner environment In Advancing Frontiers in Mycology & Mycotechnology; Satyanarayana, T., Deshmukh, S., Deshpande, M., Eds.; Springer: Singapore, 2019; pp. 229–249. [Google Scholar] [CrossRef]
- Al-Hussieny, A.A.; Imran, S.G.; Jabur, Z.A. The use of local blue-green algae in the bioremediation of hydrocarbon pollutants in wastewater from oil refineries. Plant Arch. 2020, 20, 797–802. Available online: http://www.plantarchives.org/SPL%20ISSUE%2020-2/115__797-802_.pdf (accessed on 10 July 2023).
- Arun, K.B.; Madhavan, A.; Tarafdar, A.; Sirohi, R.; Anoopkumar, A.N.; Kuriakose, L.L.; Awasthi, M.K.; Binod, P.; Varjani, S.; Sindhu, R. Filamentous fungi for pharmaceutical compounds degradation in the environment: A sustainable approach. Environ. Technol. Innov. 2023, 31, 103182. [Google Scholar] [CrossRef]
- Ghosh, S.; Rusyn, I.; Dmytruk, O.V.; Dmytruk, K.V.; Onyeaka, H.; Gryzenhout, M.; Gafforov, Y. Filamentous fungi for sustainable remediation of pharmaceutical compounds, heavy metal and oil hydrocarbons. Front. Bioeng. Biotechnol. 2023, 11, 1106973. [Google Scholar] [CrossRef] [PubMed]
- Broadway, N.M.; Dickinson, F.M.; Ratledge, C. The enzymology of dicarboxylic acid formation by Corynebacterium sp. strain 7E1C grown on n-alkanes. J. Gen. Microbiol. 1993, 139, 1337–1344. [Google Scholar] [CrossRef]
- Margesin, R.; Gander, S.; Zacke, G.; Gounot, A.M.; Schinner, F. Hydrocarbon degradation and enzyme activities of cold adapted bacteria and yeasts. Extremophiles 2003, 7, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Kadri, T.; Rouissi, T.; Brar, S.K.; Cledon, M.; Sarma, S.; Verma, M. Biodegradation of Polycyclic Aromatic Hydrocarbons (PAHs) by fungal enzymes: A review. J. Environ. Sci. 2017, 51, 52–74. [Google Scholar] [CrossRef]
- Perera, M.; Wijesundera, S.; Wijayarathna, C.D.; Seneviratne, G.; Jayasena, S. Identification of long-chain alkane-degrading (LadA) monooxygenases in Aspergillus flavus via in silico analysis. Front. Microbiol. 2022, 13, 898456. [Google Scholar] [CrossRef]
- Oudot, J.; Dupont, J.; Haloui, S.; Roquebert, M.F. Biodegradation potential of hydrocarbon-assimilating tropical fungi. Soil Biol. Biochem. 1993, 25, 1167–1173. [Google Scholar] [CrossRef]
- Chaîneau, C.H.; Morel, J.; Dupont, J.; Bury, E.; Oudot, J. Comparison of the fuel oil biodegradation potential of hydrocarbon-assimilating microorganisms isolated from a temperate agricultural soil. Sci. Total Environ. 227. 1999, 227, 237–247. [Google Scholar] [CrossRef]
- Elshafie, A.; AlKindi, A.Y.; Al-Busaidi, S.; Bakheit, C.; Albahry, S.N. Biodegradation of crude oil and n-alkanes by fungi isolated from Oman. Mar. Pollut. Bull. 2007, 54, 1692–1696. [Google Scholar] [CrossRef]
- Garzoli, L.; Gnavi, G.; Tamma, F.; Tosi, S.; Varese, G.; Picco, A.M. Sink or swim: Updated knowledge on marine fungi associated with wood substrates in the Mediterranean Sea and hints about their potential to remediate hydrocarbons. Prog. Oceanogr. 2015, 137, 140–148. [Google Scholar] [CrossRef]
- Yuan, X.; Zhang, X.; Chen, X.; Kong, D.; Liu, X.; Shen, S. Synergistic degradation of crude oil by indigenous bacterial consortium and exogenous fungus Scedosporium boydii. Bioresour. Technol. 2018, 264, 190–197. [Google Scholar] [CrossRef]
- Atakpa, E.O.; Zhou, H.; Jiang, L.; Ma, Y.; Liang, Y.; Li, Y.; Zhang, D.; Zhang, C. Improved degradation of petroleum hydrocarbons by co-culture of fungi and biosurfactant-producing bacteria. Chemosphere 2022, 290, 133337. [Google Scholar] [CrossRef] [PubMed]
- Nhi-Cong, L.T.; Mikolasch, A.; Klenk, H.-P.; Schauer, F. Degradation of the multiple branched alkane 2,6,10,14-tetramethyl-pentadecane (pristane) in Rhodococcus ruber and Mycobacterium neoaurum. Int. Biodeterior Biodegrad. 2009, 63, 201–207. [Google Scholar] [CrossRef]
- Rontani, J.F.; Bertrand, J.C.; Blanc, F.; Giusti, G. Gas chromatography and gas chromatography/mass spectrometry applied to the determination of a new pathway of pristane degradation by a marine mixed bacterial population. Mar. Chem. 1986, 18, 9–16. [Google Scholar] [CrossRef]
- Kunihiro, N.; Haruki, M.; Takano, K.; Morikawa, M.; Kanaya, S. Isolation and characterization of Rhodococcus sp. strains TMP2 and T12 that degrade 2,6,10,14-tetramethylpentadecane (pristane) at moderately low temperatures. J. Biotechnol. 2005, 115, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Simister, R.L.; Poutasse, C.M.; Thurston, A.M.; Reeve, J.L.; Baker, M.C.; White, H.K. Degradation of oil by fungi isolated from Gulf of Mexico beaches. Mar. Pollut. Bull. 2015, 100, 327–333. [Google Scholar] [CrossRef]
- Al-Otibi, F.; Al-Zahrani, R.M.; Marraiki, N. Biodegradation of Selected Hydrocarbons by Fusarium Species Isolated from Contaminated Soil Samples in Riyadh, Saudi Arabia. J. Fungi 2023, 9, 216. [Google Scholar] [CrossRef]
- Donath, M.S. Untersuchungen zum Mikrobiellen Abbau von Erdölbestandteilen unter Besonderer Berücksichtigung von Verzweigten und Cyclischen Kohlenwasserstoffen und dem Strukturanalytischen Nachweis Gebildeter Produkte. Master‘s Thesis, Universität Greifswald, Greifswald, Germany, 22 December 2015. [Google Scholar]
- Mikolasch, A.; Berzhanova, R.; Omirbekova, A.; Reinhard, A.; Zühlke, D.; Meister, M.; Mukasheva, T.; Riedel, K.; Urich, T.; Schauer, F. Moniliella spathulata, an oil-degrading yeast, which promotes growth of barley in oil-polluted soil. Appl. Microbiol. Biotechnol. 2021, 105, 401–415. [Google Scholar] [CrossRef]
- Silva, R.A.; Grossi, V.; Alvarez, H.M. Biodegradation of phytane (2,6,10,14-tetramethylhexadecane) and accumulation of related isoprenoid waxesters by Mycobacterium ratisbonense strain SD4 under nitrogen-starved conditions. FEMS Microbiol. Lett. 2007, 272, 220–228. [Google Scholar] [CrossRef]
- Nakajima, K.; Sato, A.; Takahara, Y.; Lida, T. Microbial oxidation of Isoprenoid Alkanes, Phytane, Norpristane and Farnesane. Agric. Biol. Chem. 1985, 49, 1993–2002. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | GenBank Accession N° of Isolate | NCBI ITS Database | Mycobank ITS Database | |||||
|---|---|---|---|---|---|---|---|---|
| Query Length (nt) * | Best Hit (Accession N°) ** | Identity (%) | Best Hit (Description) *** | Score | Overlap (%) | Identity (%) | ||
| SBUG-M1741 | OR335318 | 529 | Penicillium javanicum (MH865296.1) | 99.81 | Eupenicillium javanicum **** (CBS 291.53) | 835 | 100 | 99.81 |
| SBUG-M1742 | OR335319 | 521 | Penicillium javanicum (MH865296.1) | 99.81 | Eupenicillium javanicum **** (CBS 291.53) | 824 | 100 | 99.81 |
| SBUG-M1747 | OR335322 | 482 | Fusarium oxysporum (MK074845.1) | 100 | F. oxysporum species complex (LC13769 MW016603) | 765 | 100 | 100 |
| SBUG-M1749 | OR335324 | 552 | Scedosporium boydii (KP132690.1) | 100 | Scedosporium boydii (CNRMA16.348) | 876 | 100 | 100 |
| SBUG-M1751 | OR335323 | 569 | Purpureocillium lilacinum (MH426603.1) | 100 | Purpureocillium lilacinum (CNRMA18.195) | 901 | 99.82 | 100 |
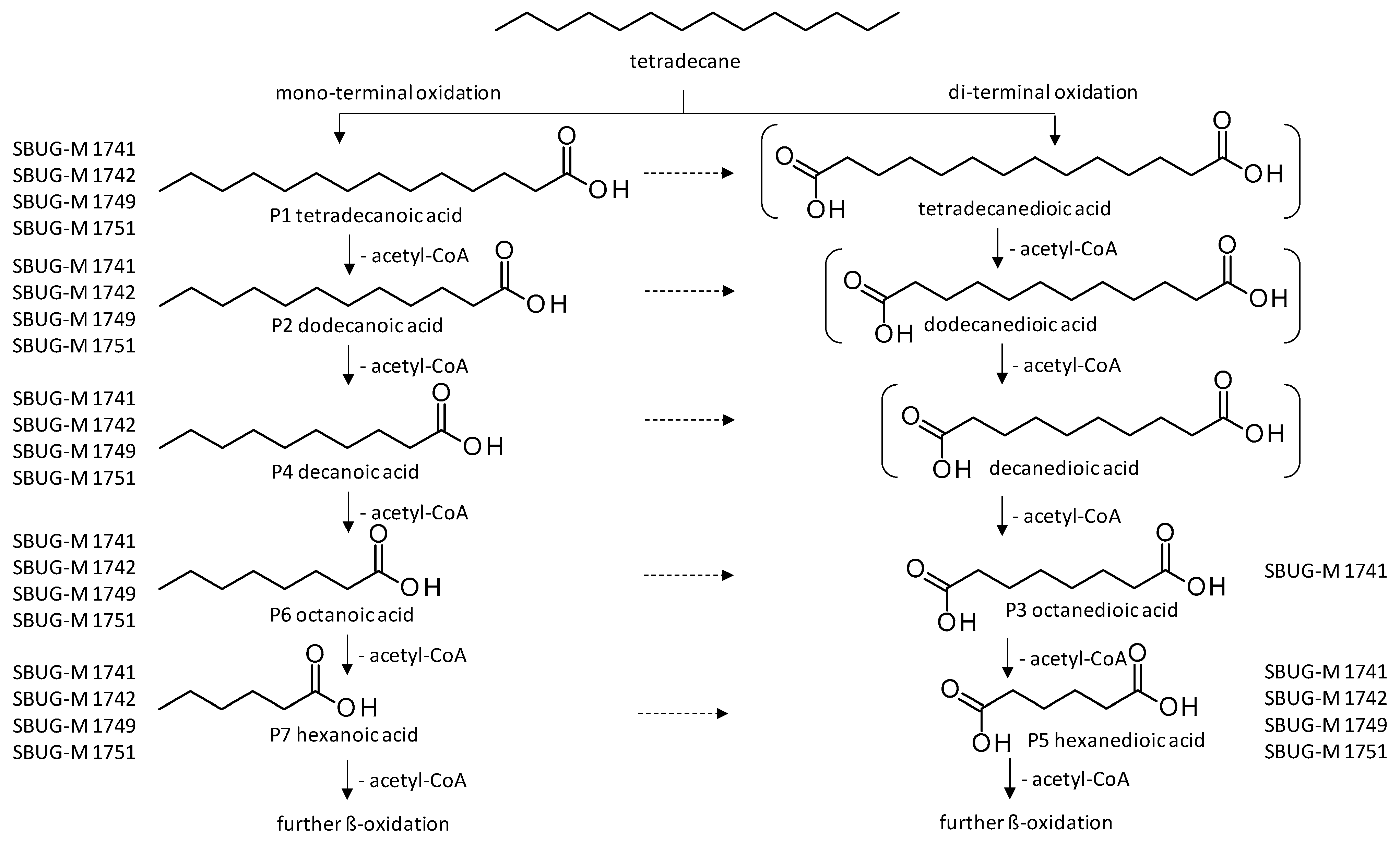
| Products | Retention Time (min) | P. javanicum | S. boydii SBUG-M1749 | P. lilacinum SBUG-M1751 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SBUG-M1741 | SBUG-M1742 | |||||||||
| Type of Culture | ||||||||||
| Shake | Static | Shake | Static | Shake | Static | Shake | Static | |||
| P1 | Tetradecanoic acid  | 32.59 | + | + | + | + | + | + | + | + |
| P2 | Dodecanoic acid  | 26.95 | + | + | + | + | + | + | + | + |
| P3 | Octanedioic acid  | 24.9 | + | + | - | - | - | - | - | - |
| P4 | Decanoic acid  | 20.65 | + | + | + | - | + | + | + | + |
| P5 | Hexanedioic acid | 18.0 | + | + | + | - | + | + | - | + |
| P6 | Octanoic acid  | 13.86 | + | + | + | + | + | + | + | + |
| P7 | Hexanoic acid  | 7.4 | + | + | + | + | + | - | - | + |
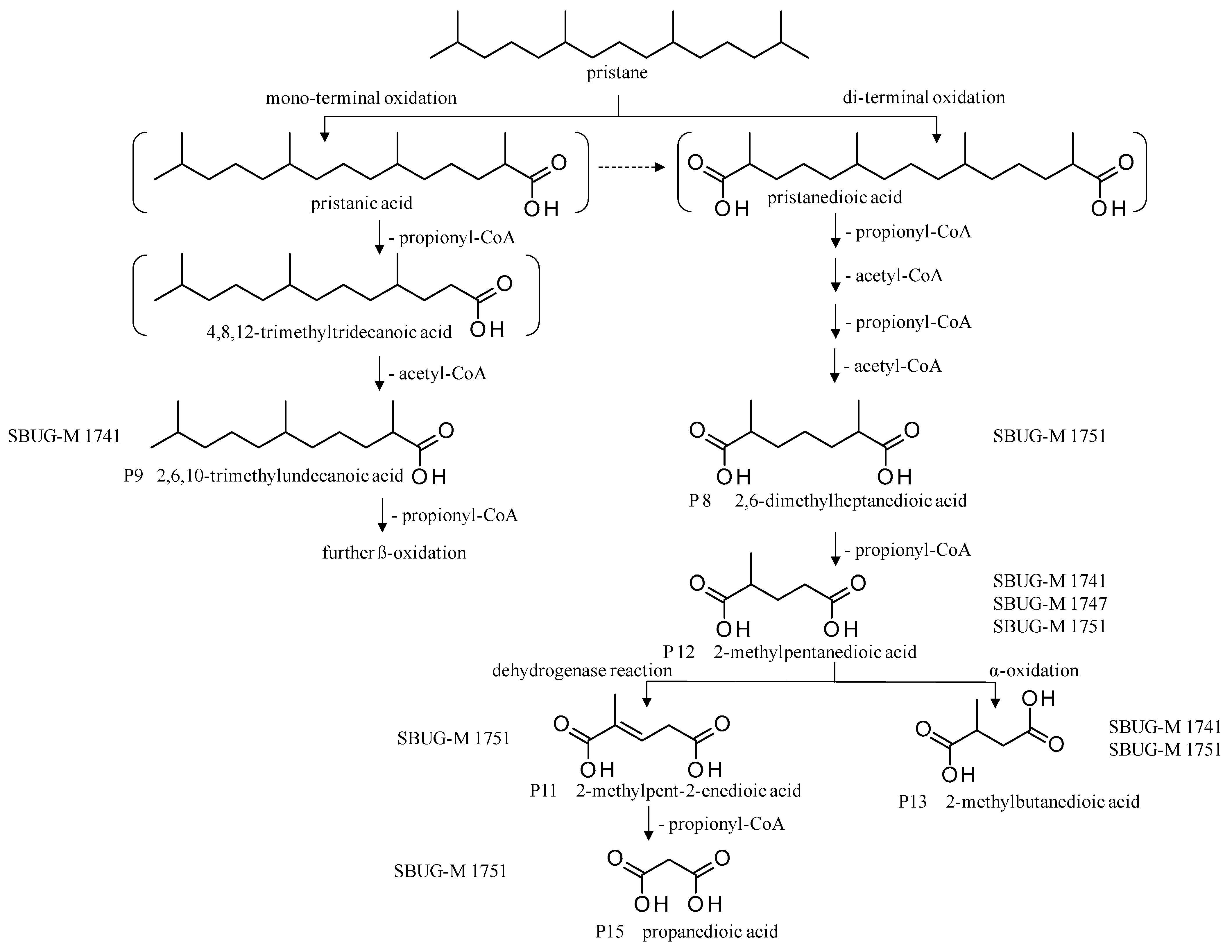
| Products | Retention Time (min) | P. javanicum SBUG-M1741 | F. oxysporum SBUG-M1747 | P. lilacinum SBUG-M1751 | ||||
|---|---|---|---|---|---|---|---|---|
| Type of Cultures | ||||||||
| Shake | Static | Shake | Static | Shake | Static | |||
| P8 | 2,6-Dimethylheptanedioic acid | 23.6 | - | - | - | - | + | + |
| P9 | 2,6,10-Trimethylundecanoic acid | 23.36 | + | - | - | - | - | - |
| P10 | 3-Methylhexanedioic acid | 19.3 | - | - | - | - | - | - |
| P11 | 2-Methylpent-2-enedioic acid | 17.7 | - | - | - | - | + | + |
| P12 | 2-Methylpentanedioic acid  | 15.6 | + | - | + | + | + | + |
| P13 | 2-Methylbutanedioic acid | 11.4 | + | - | - | - | + | + |
| P14 | 4-Methylpentanoic acid | 10.1 | - | - | - | - | - | - |
| P15 | Propanedioic acid | 7.4 | - | - | - | - | + | + |
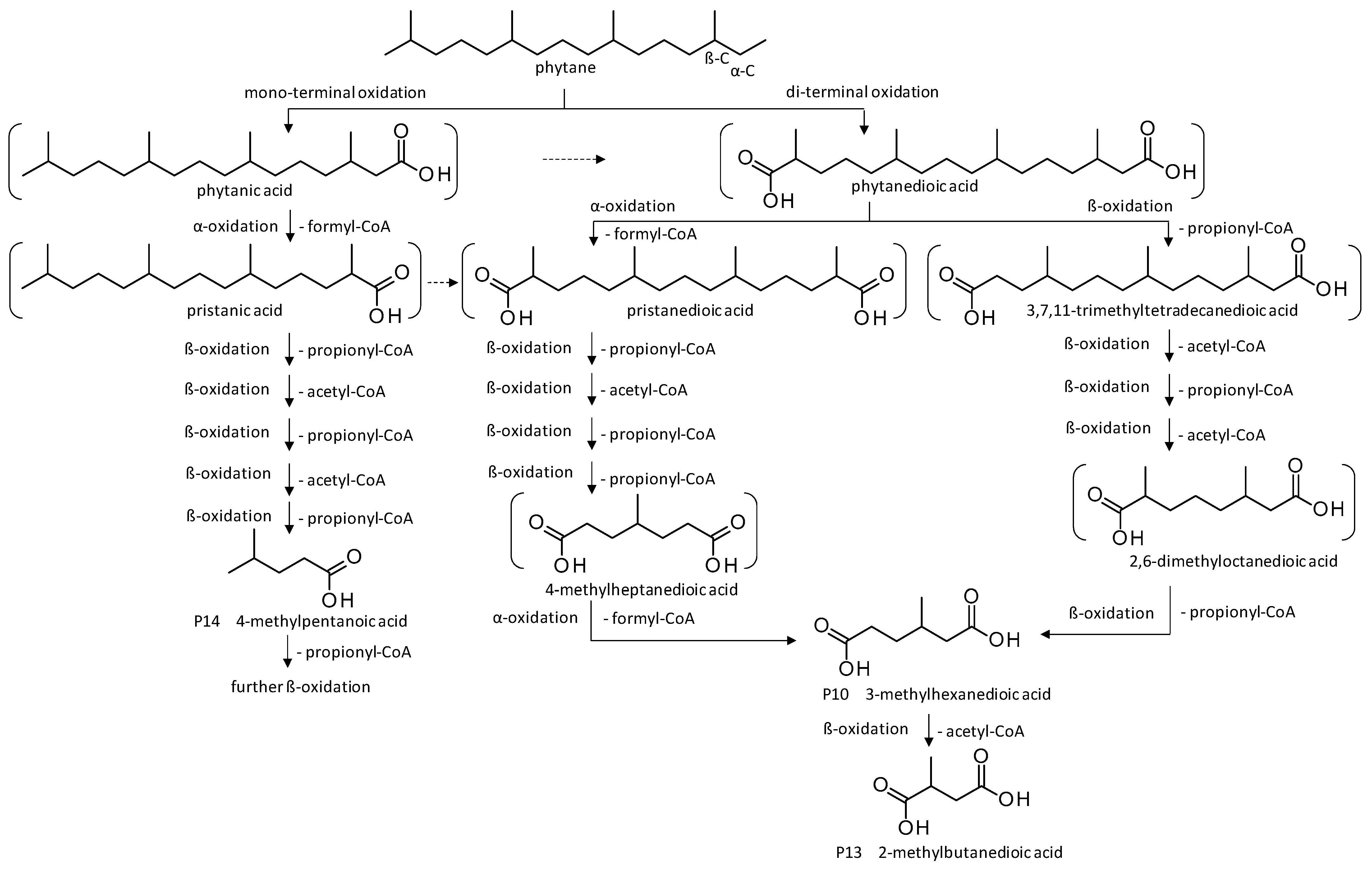
| Products | Retention Time (min) | P. lilacinum SBUG-M1751 | ||
|---|---|---|---|---|
| Type of Cultures | ||||
| Shake | Static | |||
| P10 | 3-Methylhexanedioic acid | 19.3 | + | - |
| P13 | 2-Methylbutanedioic acid | 11.4 | + | + |
| P14 | 4-Methylpentanoic acid | 10.1 | + | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaid, M.; Pöpke, D.; Reinhard, A.; Berzhanova, R.; Mukasheva, T.; Urich, T.; Mikolasch, A. Characterization of the Mycoremediation of n-Alkanes and Branched-Chain Alkanes by Filamentous Fungi from Oil-Polluted Soil Samples in Kazakhstan. Microorganisms 2023, 11, 2195. https://doi.org/10.3390/microorganisms11092195
Gaid M, Pöpke D, Reinhard A, Berzhanova R, Mukasheva T, Urich T, Mikolasch A. Characterization of the Mycoremediation of n-Alkanes and Branched-Chain Alkanes by Filamentous Fungi from Oil-Polluted Soil Samples in Kazakhstan. Microorganisms. 2023; 11(9):2195. https://doi.org/10.3390/microorganisms11092195
Chicago/Turabian StyleGaid, Mariam, Doreen Pöpke, Anne Reinhard, Ramza Berzhanova, Togzhan Mukasheva, Tim Urich, and Annett Mikolasch. 2023. "Characterization of the Mycoremediation of n-Alkanes and Branched-Chain Alkanes by Filamentous Fungi from Oil-Polluted Soil Samples in Kazakhstan" Microorganisms 11, no. 9: 2195. https://doi.org/10.3390/microorganisms11092195