

The Good, the Bad, and the Useable Microbes within the Common Alder (Alnus glutinosa) Microbiome—Potential Bio-Agents to Combat Alder Dieback

Abstract
:1. Introduction
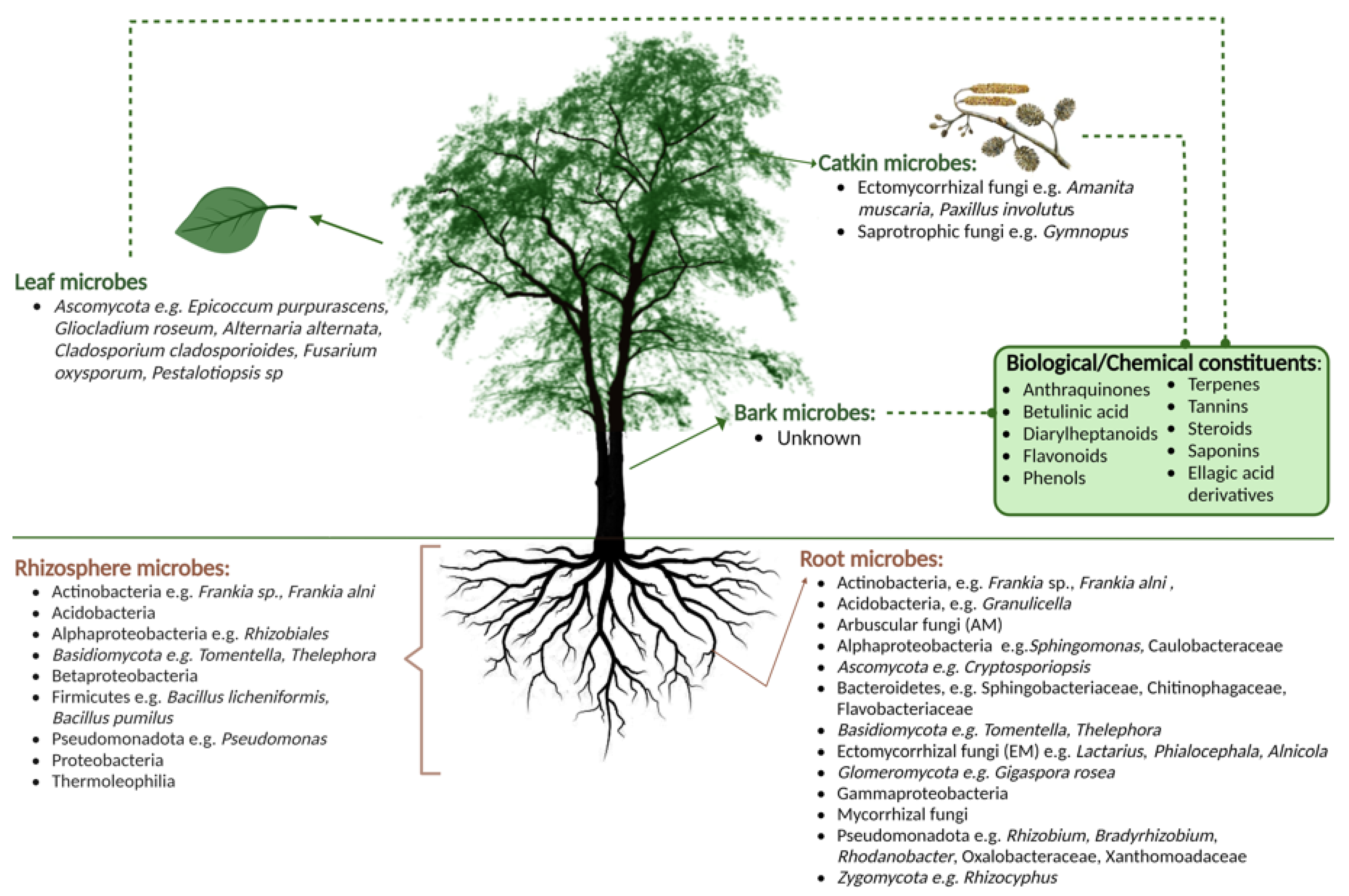
2. The Microbiome of Alder Species
What Is Known about the Microbiome of Alder?
3. Microbial Pathogens of Alder Species

4. The Potential of Microbial Biocontrol Agents in the Fight against Alder Diseases
4.1. Improving Plant Resistance via Microbe Inoculation and Genetic Resistant Breeding

{kind=link}
{kind=link}
{kind=link}
| Potential BCA | Classification | Pathogens They Potentially Control | Mechanism Expressed Showing Bio-Control Abilities | In Vitro/In Planta/In Vivo/Field/ Commercial | Reference |
|---|---|---|---|---|---|
| A. extensa, A. stenoptera, A. alata, A. pulchella | Legume (Acacia) | Phytophthora cinnamomi | Suppression and containment of Phytophthora cinnamomi inoculum in soil-infested areas to protect susceptible species and enhance species diversity | In planta | [125] |
| Bacillus amyloliquefaciens | Firmicutes | Phytophthora cactorum | Induces systemic resistance | In vitro | [151] |
| Pseudomonas chlororaphis | Gammaproteobacteria | Phytophthora palmivor | Antibiosis and suppression; reduce symptom severity; production of lytic enzymes, siderophores, and bio-surfactants | In vitro/in planta | [150] |
| Pseudomonas putida 06909 | Gammaproteobacteria | Phytophthora spp. | Hypovirulence and suppression; colonise pathogenic hyphae and reduce pathogenic populations | Field | [149] |
| Enterobacter aerogenes | Proteobacteria | Phytophthora lateralis | Suppression; reduce disease symptoms and promote plant growth | Field | [124] |
| Hypoxylon rubiginosum | Ascomycetes | Hymenoscyphus fraxineus | Antibiosis; production of metabolites that are toxic to H. fraxineus | In vitro/in planta | [148] |
| Aureimonas altamirensis | Alphaproteobacteria | Hymenoscyphus fraxineus | Colonisation resistance; exopolysaccharide production; protein secretion; stress adaptation genes | In vitro/in planta | [129] |
| Bacillus subtilis | Firmicutes | Fusarium sp. | Antibiosis and antagonistic effects | In vitro | [144] |
| Pseudomonas fluorescens | Pseudomonas | Fusarium oxysporum, Rhizoctonia solani, Macrophomira phaseolina, Fusarium sp. | Antibiosis, hypovirulence, and suppression; inhibit plant disease, minimise fungal infections, protect seeds and roots from infection; production of secondary metabolites | In vitro/field | [51,52,167,168] |
| daidzein | Isoflavones | No specific pathogen | Antibiosis; antioxidant, anti-inflammatory, and anticancer effects | In vitro/in vivo | [133,134] |
| Streptomyces | Actinobacteria | Acidovorax spp., Fusarium sp., Ralstonia solanacearum, Xanthomonas spp., Sclerotinia sclerotiorum, Erwinia amylovora | Antibiosis and induced resistance; source of bioactive compounds like antimetabolites, antibiotics, extracellular enzymes, and antitumor agents | In vitro/in vivo | [48,130,131,132,133] |
| Chitinase | Enzyme | Phytophthora cinnamomi | Suppression; reduced spread, reduced sporangia production, applied in agriculture during crop cultivation for disease management, improved growth, and greater yields | Commercial/in vitro | [142,143] |
| IAA | Auxin Plant Hormone | Fusarium sp. | Systemic resistance; regulate growth and development processes | Commercial/in vitro | [38,53,145] |
4.2. Microbiome Engineering and Its Potential Phytophtoria Control
5. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Savill, P. The Silviculture of Trees Used in British Forestry. CABI Int. 2013, 2, 29–37. [Google Scholar]
- Lalancette, S.; Lerat, S.; Roy, S.; Beaulieu, C. Fungal Endophytes of Alnus incana ssp. rugosa and Alnus alnobetula ssp. crispa and their potential to tolerate heavy metals and to promote plant growth. Mycobiology 2019, 47, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Claessens, H.; Oosterbaan, A.; Savill, P.; Rondeux, J. A review of the characteristics of black alder (Alnus glutinosa (L.) Gaertn.) and their implications for silvicultural practices. For. Int. J. For. Res. 2010, 83, 163–175. [Google Scholar] [CrossRef]
- Durrant, T.; de Rigo, D.; Caudullo, G. Alnus glutinosa in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016; pp. 64–65. [Google Scholar]
- Petruzzello, M. Alder. Encyclopedia Britannica. 2021. Available online: https://www.britannica.com/plant/alder (accessed on 26 September 2022).
- Mabberley, D.J. Mabberley’s Plant-Book: A Portable Dictionary of Plants, Their Classifications and Uses; Cambridge University Press: Cambridge, UK, 2008; Volume 3, pp. 1–1021. [Google Scholar]
- Parnell, J.; Curtis, T.G. Webb’s an Irish Flora; Cork University Press: Cork, Ireland, 2012; pp. 1–556. [Google Scholar]
- Roy, S.; Khasa, D.; Greer, C. Combining alders, frankiae, and mycorrhizae for the revegetation and remediation of contaminated ecosystems. Can. J. Bot. 2011, 85, 237–251. [Google Scholar] [CrossRef]
- McVean, D.N. Alnus glutinosa (L.) Gaertn. J. Ecol. 1953, 41, 447–466. [Google Scholar] [CrossRef]
- Buckley, D.J. Observations on Two Non-Native Alder Species (Betulaceae) Naturalising in Ireland. Br. Ir. Bot. 2021, 3, 90–98. [Google Scholar] [CrossRef]
- Woodland Trust. Alder (Alnus glutinosa). 2022. Available online: https://www.woodlandtrust.org.uk/trees-woods-and-wildlife/british-trees/a-z-of-british-trees/alder/ (accessed on 27 September 2022).
- Keet, J.H.; Robertson, M.P.; Richardson, D.M. Alnus glutinosa (Betulaceae) in South Africa: Invasive potential and management options. South Afr. J. Bot. 2020, 135, 280–293. [Google Scholar] [CrossRef]
- Funk, D.T. Alnus glutinosa (L.) Gaertn. European Alder. In Agriculture Handbook; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990; Volume 654, pp. 239–256. [Google Scholar]
- Kajba, D.; Gračan, J. EUFORGEN Technical Guidelines for Genetic Conservation and Use Black Alder (Alnus glutinosa); International Plant Genetic Resources Institute (IPGRI): Rome, Italy, 2003; p. 4. [Google Scholar]
- Moore, C. Alder, Why the tree of death? The Pallasboy Project. 2015. Available online: https://thepallasboyvessel.wordpress.com/2015/11/18/alder-why-the-tree-of-death/ (accessed on 9 December 2022).
- Sheridan, O. Alder (Alnus glutinosa) More Than Just a Native Species. 2020. Available online: https://www.teagasc.ie/news--events/daily/forestry/alder-alnus-glutinosa-more-than-just-a-native-species.php (accessed on 19 September 2022).
- Sati, S.C.; Sati, N.; Sati, O.P. Bioactive constituents and medicinal importance of genus Alnus. Pharmacogn. Rev. 2011, 5, 174–183. [Google Scholar] [CrossRef]
- Acero, N.; Muñoz-Mingarro, D. Effect on tumor necrosis factor-α production and antioxidant ability of black alder, as factors related to its anti-inflammatory properties. J. Med. Food 2012, 15, 542–548. [Google Scholar] [CrossRef]
- Nawirska-Olszańska, A.; Zaczyńska, E.; Czarny, A.; Kolniak-Ostek, J. Chemical Characteristics of Ethanol and Water Extracts of Black Alder (Alnus glutinosa L.) Acorns and Their Antibacterial, Anti-Fungal and Antitumor Properties. Molecules 2022, 27, 2804. [Google Scholar] [CrossRef] [PubMed]
- Thiem, D.; Gołębiewski, M.; Hulisz, P.; Piernik, A.; Hrynkiewicz, K. How Does Salinity Shape Bacterial and Fungal Microbiomes of Alnus glutinosa Roots? Front. Microbiol. 2018, 9, 651. [Google Scholar] [CrossRef]
- Fennessy, J. Common alder (Alnus glutinosa) as a forest tree in Ireland. Coford Connect. 2004, 80–84. Available online: http://www.coford.ie/media/coford/content/publications/projectreports/cofordconnects/Alder-reprod.pdf (accessed on 2 November 2022).
- Kennedy, C.E.J.; Southwood, T.R.E. The Number of Species of Insects Associated with British Trees: A Re-Analysis. J. Anim. Ecol. 1984, 53, 455–478. [Google Scholar] [CrossRef]
- Bjelke, U.; Boberg, J.; Oliva, J.; Tattersdill, K.; Mckie, B.G. Dieback of riparian alder caused by the Phytophthora alni complex: Projected consequences for stream ecosystems. Freshw. Biol. 2016, 61, 565–579. [Google Scholar] [CrossRef]
- Klaassen, R.K.W.M.; Creemers, J.G.M. Wooden foundation piles and its underestimated relevance for cultural heritage. J. Cult. Herit. 2012, 13, 123–128. [Google Scholar] [CrossRef]
- European Commission. New EU forest Strategy for 2030. 2023. Available online: https://environment.ec.europa.eu/strategy/forest-strategy_en#:~:text=New%20EU%20forest%20strategy%20for%202030&text=The%20strategy%20will%20contribute%20to,and%20climate%20neutrality%20by%202050 (accessed on 18 July 2023).
- Valor, T.; Camprodon, J.; Buscarini, S.; Casals, P. Drought-induced dieback of riparian black alder as revealed by tree rings and oxygen isotopes. For. Ecol. Manag. 2020, 478, 1–9. [Google Scholar] [CrossRef]
- Teshome, D.T.; Zharare, G.E.; Naidoo, S. The Threat of the Combined Effect of Biotic and Abiotic Stress Factors in Forestry Under a Changing Climate. Front. Plant Sci. 2020, 11, 1874. [Google Scholar] [CrossRef]
- O’Hanlon, R.; Wilson, J.; Cox, D. Investigations into Phytophthora dieback of alder along the river Lagan in Belfast, Northern Ireland, ed. B. Tobin. Ir. For. Soc. Ir. For. 2020, 77, 33–43. [Google Scholar]
- Mercado-Blanco, J.; Abrantes, I.; Barra Caracciolo, A.; Bevivino, A.; Ciancio, A.; Grenni, P.; Hrynkiewicz, K.; Kredics, L.; Proença, D.N. Belowground Microbiota and the Health of Tree Crops. Front. Microbiol. 2018, 9, 1006. [Google Scholar] [CrossRef] [PubMed]
- Gałązka, A.; Marzec-Grządziel, A.; Varsadiya, M.; Niedźwiecki, J.; Gawryjołek, K.; Furtak, K.; Przybyś, M.; Grządziel, J. Biodiversity and Metabolic Potential of Bacteria in Bulk Soil from the Peri-Root Zone of Black Alder (Alnus glutinosa), Silver Birch (Betula pendula) and Scots Pine (Pinus sylvestris). Int. J. Mol. Sci. 2022, 23, 2633. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P. Forest microbiome: Diversity, complexity and dynamics. FEMS Microbiol. Rev. 2016, 41, 109–130. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, V.; Selvaraj, G.; Bais, H.P. Functional soil microbiome: Belowground solutions to an aboveground problem. Plant Physiol 2014, 166, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Lin, X.; Tian, J.; Ji, L.; Chen, Y.; Tran, L.-S.P.; Tian, C. Research Advances of Beneficial Microbiota Associated with Crop Plants. Int. J. Mol. Sci. 2020, 21, 1792. [Google Scholar] [CrossRef] [PubMed]
- Gafur, A. Plant growth promoting microbes (PGPM) for the sustainability of tropical plantation forests in Indonesia. Acad. Open 2021, 1–3. [Google Scholar] [CrossRef]
- Vocciante, M.; Grifoni, M.; Fusini, D.; Petruzzelli, G.; Franchi, E. The Role of Plant Growth-Promoting Rhizobacteria (PGPR) in Mitigating Plants Environmental Stresses. Appl. Sci. 2022, 12, 1231. [Google Scholar] [CrossRef]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Forest Soil Bacteria: Diversity, Involvement in Ecosystem Processes, and Response to Global Change. Microbiol. Mol. Biol. Rev. 2017, 81, e00063-16. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef]
- Fu, S.F.; Wei, J.Y.; Chen, H.W.; Liu, Y.Y.; Lu, H.Y.; Chou, J.Y. Indole-3-acetic acid: A widespread physiological code in interactions of fungi with other organisms. Plant Signal. Behav. 2015, 10, e1048052. [Google Scholar] [CrossRef]
- Polko, J.K.; Kieber, J.J. 1-Aminocyclopropane 1-Carboxylic Acid and Its Emerging Role as an Ethylene-Independent Growth Regulator. Front. Plant Sci. 2019, 10, 1602. [Google Scholar] [CrossRef]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Sulochana, M.B.; Jayachandra, S.Y.; Kumar, S.A.; Parameshwar, A.B.; Reddy, K.M.; Dayanand, A. Siderophore as a potential plant growth-promoting agent produced by Pseudomonas aeruginosa JAS-25. Appl. Biochem. Biotechnol. 2014, 174, 297–308. [Google Scholar]
- Behera, B.C.; Sethi, B.K.; Mishra, R.R.; Dutta, S.K.; Thatoi, H.N. Microbial cellulases—Diversity & biotechnology with reference to mangrove environment: A review. J. Genet. Eng. Biotechnol. 2017, 15, 197–210. [Google Scholar] [PubMed]
- Bhattacharyya, C.; Banerjee, S.; Acharya, U.; Mitra, A.; Mallick, I.; Haldar, A.; Haldar, S.; Ghosh, A.; Ghosh, A. Evaluation of plant growth promotion properties and induction of antioxidative defense mechanism by tea rhizobacteria of Darjeeling, India. Sci. Rep. 2020, 10, 15536. [Google Scholar]
- Phitsuwan, P.; Laohakunjit, N.; Kerdchoechuen, O.; Kyu, K.L.; Ratanakhanokchai, K. Present and potential applications of cellulases in agriculture, biotechnology, and bioenergy. Folia Microbiol. 2013, 58, 163–176. [Google Scholar]
- Richter, B.S.; Ivors, K.; Shi, W.; Benson, D.M. Cellulase activity as a mechanism for suppression of phytophthora root rot in mulches. Phytopathology 2011, 101, 223–230. [Google Scholar]
- Confortin, T.C.; Spannemberg, S.S.; Todero, I.; Luft, L.; Brun, T.; Albornoz, E.; Kuhn, R.C.; Mazutti, M. Chapter 21—Microbial Enzymes as Control Agents of Diseases and Pests in Organic Agriculture. In New and Future Developments in Microbial Biotechnology and Bioengineering; Gupta, V.K., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 321–332. [Google Scholar]
- Callender, K.L.; Roy, S.; Khasa, D.P.; Whyte, L.G.; Greer, C.W. Actinorhizal Alder Phytostabilization Alters Microbial Community Dynamics in Gold Mine Waste Rock from Northern Quebec: A Greenhouse Study. PLoS ONE 2016, 11, e0150181. [Google Scholar]
- McEwan, N.R.; Wilkinson, T.J.; Girdwood, S.E.; Snelling, T.J.; Peate, T.; Dougal, K.; Jones, D.L.; Godbold, D.L. Evaluation of the microbiome of decaying alder nodules by next generation sequencing. Endocytobiosis Cell Res. 2017, 28, 14–19. [Google Scholar]
- Zappelini, C.; Alvarez-Lopez, V.; Capelli, N.; Guyeux, C.; Chalot, M. Streptomyces Dominate the Soil Under Betula Trees That Have Naturally Colonized a Red Gypsum Landfill. Front. Microbiol. 2018, 9, 1772. [Google Scholar] [PubMed]
- Elo, S.; Maunuksela, L.; Salkinoja-Salonen, M.; Smolander, A.; Haahtela, K. Humus bacteria of Norway spruce stands: Plant growth promoting properties and birch, red fescue and alder colonizing capacity. FEMS Microbiol. Ecol. 2000, 31, 143–152. [Google Scholar] [PubMed]
- Dukunde, A.; Schneider, D.; Schmidt, M.; Veldkamp, E.; Daniel, R. Tree Species Shape Soil Bacterial Community Structure and Function in Temperate Deciduous Forests. Front. Microbiol. 2019, 10, 1519. [Google Scholar]
- Probanza, A.; Lucas, J.A.; Acero, N.; Gutierrez Mañero, F.J. The influence of native rhizobacteria on European alder (Alnus glutinosa (L.) Gaertn.) Growth. Plant Soil 1996, 182, 59–66. [Google Scholar]
- Gutiérrez-Mañero, F.J.; Ramos, B.; Probanza, A.; Mehouachi, J.; Tadeo, F.R.; Talon, M. The plant-growth-promoting rhizobacteria Bacillus pumilus and Bacillus licheniformis produce high amounts of physiologically active gibberellins. Physiol. Plant. 2001, 111, 206–211. [Google Scholar]
- Egamberdiyeva, D. Effect of plant growth promoting bacteria on growth of Scots pine and silver birch seedlings. Uzb. J. Agric. 2005, 1, 66–69. [Google Scholar]
- Imperato, V.; Kowalkowski, L.; Portillo-Estrada, M.; Gawronski, S.W.; Vangronsveld, J.; Thijs, S. Characterisation of the Carpinus betulus L. Phyllomicrobiome in Urban and Forest Areas. Front. Microbiol. 2019, 10, 1110. [Google Scholar]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological Control of Plant Pathogens: A Global Perspective. Microorganisms 2022, 10, 596. [Google Scholar]
- Dąbrowska, G.B.; Garstecka, Z.; Trejgell, A.; Dąbrowski, H.P.; Konieczna, W.; Szyp-Borowska, I. The Impact of Forest Fungi on Promoting Growth and Development of Brassica napus L. Agronomy 2021, 11, 2475. [Google Scholar]
- Thiem, D.; Piernik, A.; Hrynkiewicz, K. Ectomycorrhizal and endophytic fungi associated with Alnus glutinosa growing in a saline area of central Poland. Symbiosis 2018, 75, 17–28. [Google Scholar]
- Thiem, D.; Tyburski, J.; Golebiewski, M.; Hrynkiewicz, K. Halotolerant fungi stimulate growth and mitigate salt stress in Alnus glutinosa Gaertn. Dendrobiology 2020, 83, 30–42. [Google Scholar]
- Xu, R.; Li, T.; Cui, H.; Wang, J.; Yu, X.; Ding, Y.; Wang, C.; Yang, Z.; Zhao, Z. Diversity and characterization of Cd-tolerant dark septate endophytes (DSEs) associated with the roots of Nepal alder (Alnus nepalensis) in a metal mine tailing of southwest China. Appl. Soil Ecol. 2015, 93, 11–18. [Google Scholar]
- Cordero, R.J.; Casadevall, A. Functions of fungal melanin beyond virulence. Fungal Biol. Rev. 2017, 31, 99–112. [Google Scholar]
- Hrynkiewicz, K.; Szymańska, S.; Piernik, A.; Thiem, D. Ectomycorrhizal Community Structure of Salix and Betula spp. at a Saline Site in Central Poland in Relation to the Seasons and Soil Parameters. Water Air Soil Pollut. 2015, 226, 99. [Google Scholar] [PubMed]
- Sousa, N.R.; Franco, A.R.; Ramos, M.A.; Oliveria, R.S.; Castro, P.M.L. The response of Betula pubescens to inoculation with an ectomycorrhizal fungus and a plant growth promoting bacterium is substrate-dependent. Ecol. Eng. 2015, 81, 439–443. [Google Scholar]
- Badalamenti, E.; Catania, V.; Sofia, S.; Sardina, M.T.; Sala, G.; La Mantia, T.; Quatrini, P. The Root Mycobiota of Betula aetnensis Raf., an Endemic Tree Species Colonizing the Lavas of Mt. Etna (Italy). Forests 2021, 12, 1624. [Google Scholar]
- Orfanoudakis, M.; Wheeler, C.T.; Hooker, J.E. Both the arbuscular mycorrhizal fungus Gigaspora rosea and Frankia increase root system branching and reduce root hair frequency in Alnus glutinosa. Mycorrhiza 2010, 20, 117–126. [Google Scholar]
- Karunarathna, S.C.; Ashwath, N.; Jeewon, R. Editorial: The Potential of Fungi for Enhancing Crops and Forestry Systems. Front. Microbiol. 2021, 12, 813051. [Google Scholar] [PubMed]
- Argumedo-Delira, R.; Gómez-Martínez, M.J.; Mora-Delgado, J. Plant Growth Promoting Filamentous Fungi and Their Application in the Fertilization of Pastures for Animal Consumption. Agronomy 2022, 12, 3033. [Google Scholar]
- Aguiar-Pulido, V.; Huang, W.; Suarez-Ulloa, V.; Cickovski, T.; Mathee, K.; Narasimhan, G. Metagenomics, Metatranscriptomics, and Metabolomics Approaches for Microbiome Analysis. Evol. Bioinform Online 2016, 12 (Suppl. S1), 5–16. [Google Scholar]
- Pujic, P.; Alloisio, N.; Miotello, G.; Armengaud, J.; Abrouk, D.; Fournier, P.; Normand, P. The Proteogenome of Symbiotic Frankia alni in Alnus glutinosa Nodules. Microorganisms 2022, 10, 651. [Google Scholar]
- Gomes Marques, I.; Faria, C.; Conceição, S.I.R.; Jansson, R.; Corcobando, T.; Milanovic, S.; Laurent, Y.; Bernez, I.; Dufour, S.; Mandák, B.; et al. Germination and seed traits in common alder (Alnus spp.): The potential contribution of rear-edge populations to ecological restoration success. Restor. Ecol. 2022, 30, e13517. [Google Scholar]
- Zaspel, I.; Naujoks, G.; Krüger, L.; Pham, L.H. Promotion of resistance of black alder clones (Alnus glutinosa (L.) Gaertn.) against Phytophthora alni ssp. alni by cyclolipopeptide producing bacteria. Silvae Genet. 2014, 63, 222–229. [Google Scholar]
- Vacek, Z.; Vacek, S.; Cukor, J.; Bulušek, D.; Slávik, M.; Lukáčik, I.; Štefančík, I.; Sitková, Z.; Eşen, D.; Ripullone, F.; et al. Dendrochronological data from twelve countries proved definite growth response of black alder ([L.] Gaertn.) to climate courses across its distribution range. Cent. Eur. For. J. 2022, 68, 139–153. [Google Scholar] [CrossRef]
- Asghari, R.; Rahimian, H.; Babaeizad, V. Isolation of Rhizobium strains from Alder root nodules in Mazandaran. In Proceedings of the 22nd Iran Plant Protection Congress, Karaj, Iran, 27–30 August 2016; Volume 2, p. 103. [Google Scholar]
- Bogar, L.M.; Dickie, I.A.; Kennedy, P.G. Testing the co-invasion hypothesis: Ectomycorrhizal fungal communities on Alnus glutinosa and Salix fragilis in New Zealand. Divers. Distrib. 2015, 21, 268–278. [Google Scholar] [CrossRef]
- Sukhikh, S.; Ivanova, S.; Skrypnik, L.; Bakhtiyarova, A.; Larina, V.; Krol, O.; Prosekov, A.; Frolov, A.; Povydysh, M.; Babich, O. Study of the Antioxidant Properties of Filipendula ulmaria and Alnus glutinosa. Plants 2022, 11, 2415. [Google Scholar] [CrossRef]
- Mushkina, V. Effect of wound healing in gels containing tinctures of Alnus glutinosa (L) leaves. Clin. Phytoscience 2021, 7, 62. [Google Scholar] [CrossRef]
- Altınyay, Ç.; Eryılmaz, M.; Yazgan, A.N.; Sever Yılmaz, B.; Altun, M.L. Antimicrobial activity of some Alnus species. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4671–4674. [Google Scholar]
- Kayini, A.; Pandey, R.R. Phyllosphere Fungi of Alnus nepalensis, Castanopsis hystrix and Schima walichii in a Subtropical Forest of North East India. J. Am. Sci. 2010, 6, 118–123. [Google Scholar]
- Berges, R.; Seemüller, E. Impact of phytoplasma infection of common alder (Alnus glutinosa) depends on strain virulence. For. Pathol. 2002, 32, 357–363. [Google Scholar] [CrossRef]
- Atanasova, B.; Spasov, D.; Jakovljević, M.; Jović, J.; Krstić, O.; Mitrović, M.; Cvrković, T. First Report of Alder Yellows Phytoplasma Associated with Common Alder (Alnus glutinosa) in the Republic of Macedonia. Plant Dis. 2014, 98, 1268. [Google Scholar] [CrossRef] [PubMed]
- Marcone, C.; Franco-Lara, L.; Toševski, I. Major Phytoplasma Diseases of Forest and Urban Trees. In Phytoplasmas: Plant Pathogenic Bacteria—I; Rao, G., Bertaccini, A., Fiore, N., Liefting, L., Eds.; Springer: Singapore, 2018; pp. 287–312. [Google Scholar]
- Cvrković, T.; Jović, J.; Mitrović, M.; Petrović, A.; Krnjajić, S.; Malembic-Maher, S.; Toševski, I. First report of alder yellows phytoplasma on common alder (Alnus glutinosa) in Serbia. Plant Pathol. 2008, 57, 773. [Google Scholar] [CrossRef]
- Forbes, H. Plant of the Week—16 November—Alder tongue (Taphrina alni) (fungus), in Botany in Scotland. 2020. Available online: https://botsocscot.wordpress.com/2020/11/15/plant-of-the-week-date-alder-tongue-taphrina-alni-fungus/ (accessed on 25 March 2023).
- Sims, L. Alder (Alnus spp.)-Leaf Spot. 2012. Available online: https://pnwhandbooks.org/plantdisease/host-disease/alder-alnus-spp-leaf-spot (accessed on 25 March 2023).
- Moricca, S.; Benigno, A.; Oliveira Longa, C.M.; Cacciola, S.O.; Maresi, G. First Documentation of Life Cycle Completion of the Alien Rust Pathogen Melampsoridium hiratsukanum in the Eastern Alps Proves Its Successful Establishment in This Mountain Range. J. Fungi 2021, 7, 617. [Google Scholar] [CrossRef]
- Tomoshevich, M.; Kirichenko, N.; Holmes, K.A.; Kenis, M.; Stenlid, J. Foliar fungal pathogens of European woody plants in Siberia: An early warning of potential threats? For. Pathol. 2013, 43, 345–359. [Google Scholar] [CrossRef]
- Surico, G.; Mugnai, L.; Pastorelli, R.; Giovannetti, L.; Stead, D.E. Erwinia alni, a New Species Causing Bark Cankers of Alder (Alnus Miller) Species. Int. J. Syst. Evol. Microbiol. 1996, 46, 720–726. [Google Scholar] [CrossRef]
- Scortichini, M. Leaf necrosis and sucker and twig dieback of Alnus glutinosa incited by Pseudomonas syringae pv. syringae. Eur. J. For. Pathol. 1997, 27, 331–336. [Google Scholar] [CrossRef]
- Dolch, R.; Tscharntke, T. Defoliation of alders (Alnus glutinosa) affects herbivory by leaf beetles on undamaged neighbours. Oecologia 2000, 125, 504–511. [Google Scholar] [CrossRef] [PubMed]
- The Sawflies (Symphyta) of Britain and Ireland. Monsoma pulveratum (Retzius 1783). 2021. Available online: https://www.sawflies.org.uk/monsoma-pulveratum/ (accessed on 25 March 2023).
- The Sawflies (Symphyta) of Britain and Ireland. Eriocampa ovata (Linnaeus 1760). 2021. Available online: https://www.sawflies.org.uk/eriocampa-ovata/ (accessed on 25 March 2023).
- Edmunds, H.A.; Springate, N.D. Cimbex connatus (Schrank) (Hymenoptera: Cimbicidae): A rare species of sawfly in the British Isles. Br. J. Entomol. Nat. Hist. 1998, 11, 65–68. [Google Scholar]
- Érsek, L. Fenusa Dohrnii (Tischbein 1846) European Alder Leafminer. 2019. Available online: https://bladmineerders.nl/parasites/animalia/arthropoda/insecta/hymenoptera/symphyta/tenthredinoidea/tenthredinidae/heterarthrinae/fenusa/fenusa-dohrnii/ (accessed on 26 March 2023).
- Sims, L.L. Phytophthora Species and Riparian Alder Tree Damage in Western Oregon State University. Ph.D. Thesis, Botany and Plant Pathology, Western Oregon State University, Monmouth, OR, USA, 2014; pp. 1–12. [Google Scholar]
- Kroon, L.P.; Brouwer, H.; de Cock, A.W.A.M.; Govers, F. The genus Phytophthora anno 2012. Phytopathology 2012, 102, 348–364. [Google Scholar] [CrossRef]
- Brasier, C.; Scanu, B.; Cooke, D.; Jung, T. Phytophthora: An ancient, historic, biologically and structurally cohesive and evolutionarily successful generic concept in need of preservation. IMA Fungus 2022, 13, 12. [Google Scholar] [CrossRef]
- Haque, M. Identification, characterization and pathogenicity of Phytophthora spp. associated with the mortality of Alnus glutinosa in Spain. Ph.D. Thesis, University of Valladolid, Valladolid, Spain, 2014; p. 6. [Google Scholar]
- Cai, G.; Hillman, B.I. Chapter Twelve—Phytophthora Viruses. In Advances in Vrius Research; Ghabrial, S.A., Ed.; Academic Press: Cambridge, MA, USA, 2013; Volume 86, pp. 327–350. [Google Scholar]
- Wingfield, M.J. Pathology|Disease Affecting Exotic Plantation Species. In Encyclopedia of Forest Sciences; Burley, J., Ed.; Elsevier: Oxford, UK, 2004; pp. 816–822. [Google Scholar]
- Abad, Z.G.; Burgess, T.I.; Redford, A.J.; Bienapfl, J.C.; Srivastava, S.; Mathew, R.; Jennings, K. IDphy: An international online resource for molecular and morphological identification of Phytophthora. Plant Dis. 2023, 107, 987–998. [Google Scholar] [CrossRef]
- Oßwald, W.; Fleischmann, F.; Rigling, D.; Coelho, A.C.; Cravador, A.; Diez, J.; Dalio, R.J.; Horta Jung, M.; Pfanz, H.; Robin, C.; et al. Strategies of attack and defence in woody plant–Phytophthora interactions. For. Pathol. 2014, 44, 169–190. [Google Scholar] [CrossRef]
- Beckerman, J.; Creswell, T. Phytophthora Diseases in Ornamentals. Plant Pathology in the Landscape Series. 20201-6. Available online: https://www.extension.purdue.edu/extmedia/BP/BP-215-W.pdf (accessed on 11 January 2023).
- Nave, C.; Schwan, J.; Werres, S.; Riebesehl, J. Alnus glutinosa Threatened by Alder Phytophthora: A Histological Study of Roots. Pathogens 2021, 10, 977. [Google Scholar] [CrossRef]
- Gibbs, J.; van Dijk, C.; Webber, J. Phytophthora disease of alder in Europe. In Forestry Commission Bulletin 126; Forestry Commission: Edinburgh, UK, 2003; pp. 1–82. [Google Scholar]
- Thoirain, B.; Husson, C.; Marçais, B. Risk factors for the phytophthora-induced decline of alder in northeastern France. Phytopathology 2007, 97, 99–105. [Google Scholar] [CrossRef]
- Černý, K.; Strnadová, V. Phytophthora Alder Decline: Disease Symptoms, Causal Agent and its Distribution in the Czech Republic. Plant Prot. Sci. 2010, 46, 12–18. [Google Scholar] [CrossRef]
- Aguayo, J.; Halkett, F.; Husson, C.; Nagy, Z.Á.; Szigethy, A.; Bakonyi, J.; Frey, P.; Marçais, B. Genetic Diversity and Origins of the Homoploid-Type Hybrid Phytophthora alni. Appl. Environ. Microbiol. 2016, 82, 7142–7153. [Google Scholar] [CrossRef]
- Gibbs, J.N. Phytophthora root disease of alder in Britain. EPPO Bull. 1995, 25, 661–664. [Google Scholar] [CrossRef]
- Marçais, B. Phytophthora Alni Species Complex (Alder Phytophthora); CABI International; CABI Compendium: Wallingford, UK, 2022. [Google Scholar] [CrossRef]
- Brasier, C.M.; Kirk, S.A.; Delcan, J.; Cooke, D.E.; Jung, T.; Man in’t Veld, W.A. Phytophthora alni sp. nov. and its variants: Designation of emerging heteroploid hybrid pathogens spreading on Alnus trees. Mycol. Res. 2004, 108 Pt 10, 1172–1184. [Google Scholar] [CrossRef] [PubMed]
- Bregant, C.; Sanna, G.P.; Bottos, A.; Maddau, L.; Montecchio, L.; Linaldeddu, B.T. Diversity and Pathogenicity of Phytophthora Species Associated with Declining Alder Trees in Italy and Description of Phytophthora alpina sp. nov. Forests 2020, 11, 848. [Google Scholar] [CrossRef]
- Husson, C.; Aguayo, J.; Revellin, C.; Frey, P.; Ioos, R.; Marçais, B. Evidence for homoploid speciation in Phytophthora alni supports taxonomic reclassification in this species complex. Fungal Genet. Biol. 2015, 77, 12–21. [Google Scholar] [CrossRef]
- Trzewik, A.; Maciorowski, R.; Orlikowska, T. Pathogenicity of Phytophthora & times; alni isolates obtained from symptomatic trees, soil and water against Alder. Forests 2022, 13, 20. [Google Scholar]
- Jung, T.; Pérez-Sierra, A.; Durán, A.; Horta Jung, M.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil- and airborne Phytophthora species in forests and woodlands. Persoonia 2018, 40, 182–220. [Google Scholar] [CrossRef]
- Seddaiu, S.; Linaldeddu, B.T. First Report of Phytophthora acerina, plurivora, and P. pseudocryptogea Associated with Declining Common Alder Trees in Italy. Plant Dis. 2020, 104, 1874. [Google Scholar] [CrossRef]
- Hansen, E.; Reeser, P.; Rooney-Latham, S. Forest Phytophthoras of the World: Phytophthora siskiyouensis. For. Phytophthoras 2012, 1. [Google Scholar] [CrossRef]
- Matsiakh, I.; Kramarets, V.; Cleary, M. Occurrence and diversity of Phytophthora species in declining broadleaf forests in western Ukraine. For. Pathol. 2021, 51, e12662. [Google Scholar] [CrossRef]
- Tkaczyk, M.; Sikora, K.; Galko, J.; Kunka, A. Occurrence of Phytophthora species in riparian stands of black alder (Alnus glutinosa) in Slovakia. For. Pathol. 2023, 53, e12800. [Google Scholar] [CrossRef]
- Zamora-Ballesteros, C.; Haque, M.M.U.; Diez, J.J.; Martín-García, J. Pathogenicity of Phytophthora alni complex and P. plurivora in Alnus glutinosa seedlings. For. Pathol. 2017, 47, e12299. [Google Scholar] [CrossRef]
- Haque, M.M.U.; Martín-García, J.; Diez, J.J. Variation in pathogenicity among the three subspecies of Phytophthora alni on detached leaves, twigs and branches of Alnus glutinosa. For. Pathol. 2015, 45, 484–491. [Google Scholar] [CrossRef]
- Downing, M.C.; Jung, T.; Thomas, V.; Blaschke, M.; Tuffly, M.F.; Reich, R. Estimating the Susceptibility to Phytophthora alni Globally Using Both Statistical Analyses and Expert Knowledge; General Technical Report; Pacific Northwest Research Station, USDA Forest Service: Washington, DC, USA, 2010; Volume 802, pp. 559–570. [Google Scholar]
- Smeriglio, A.; D’Angelo, V.; Cacciola, A.; Ingegneri, M.; Raimondo, F.M.; Trombetta, D.; Germanò, M.P. New Insights on Phytochemical Features and Biological Properties of Alnus glutinosa Stem Bark. Plants 2022, 11, 2499. [Google Scholar] [CrossRef] [PubMed]
- Pirttilä, A.M.; Mohammad Parast Tabas, H.; Baruah, N.; Koskimäki, J.J. Biofertilizers and Biocontrol Agents for Agriculture: How to Identify and Develop New Potent Microbial Strains and Traits. Microorganisms 2021, 9, 817. [Google Scholar] [CrossRef]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of Action of Microbial Biological Control Agents against Plant Diseases: Relevance beyond Efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [PubMed]
- Utkhede, R.; Stephen, B.; Wong, S. Control of Phytophthora lateralis root rot of Lawson cypress with Enterobacter aerogenes. J. Arboric. 1997, 23, 144–146. [Google Scholar] [CrossRef]
- D’Souza, N.; Colquhoun, I.J.; Sheared, B.L.; Hardy, G.E. Assessing the potential for biological control of Phytophthora cinnamomi by fifteen native Western Australian jarrah-forest legume species. Australas. Plant Pathol. 2005, 34, 533–540. [Google Scholar] [CrossRef]
- Bonaterra, A.; Badosa, E.; Daranas, N.; Francés, J.; Roselló, G.; Montesinos, E. Bacteria as Biological Control Agents of Plant Diseases. Microorganisms 2022, 10, 1759. [Google Scholar] [CrossRef]
- Ulrich, K.; Becker, R.; Behrendt, U.; Kube, M.; Ulrich, A. A Comparative Analysis of Ash Leaf-Colonizing Bacterial Communities Identifies Putative Antagonists of Hymenoscyphus fraxineus. Front. Microbiol. 2020, 11, 966. [Google Scholar] [CrossRef]
- Prospero, S.; Botella, L.; Santini, A.; Robin, A. Biological control of emerging forest diseases: How can we move from dreams to reality? For. Ecol. Manag. 2021, 496, 119377. [Google Scholar] [CrossRef]
- Becker, R.; Ulrich, K.; Behrendt, U.; Schneck, V.; Ulrich, A. Genomic Characterization of Aureimonas altamirensis C2P003—A Specific Member of the Microbiome of Fraxinus excelsior Trees Tolerant to Ash Dieback. Plants 2022, 11, 3487. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and interactions in plant growth promotion. Appl. Microbiol. Biotechnol. 2019, 103, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Le, K.D.; Yu, N.H.; Park, A.R.; Park, D.J.; Kim, C.J.; Kim, J.C. Streptomyces sp. AN090126 as a Biocontrol Agent against Bacterial and Fungal Plant Diseases. Microorganisms 2022, 10, 791. [Google Scholar] [CrossRef]
- Ghanem, G.A.M.; Gebily, D.A.S.; Ragab, M.M.; Ali, A.M.; Soliman, N.E.D.K.; El-Moity, T.H.A. Efficacy of antifungal substances of three Streptomyces spp. against different plant pathogenic fungi. Egypt. J. Biol. Pest Control 2022, 32, 112. [Google Scholar] [CrossRef]
- Liu, N.; Wang, H.; Lie, M.; Gu, Q.; Zheng, W.; Huang, Y. Streptomyces alni sp. nov., a daidzein-producing endophyte isolated from a root of Alnus nepalensis D. Don. Int. J. Syst. Evol. Microbiol. 2009, 59, 254–258. [Google Scholar] [CrossRef]
- Alshehri, M.M.; Sharifi-Rad, J.; Herrera-Bravo, J.; Jara, E.L.; Salazar, L.A.; Kregiel, D.; Uprety, Y.; Akram, M.; Iqbal, M.; Martorell, M.; et al. Therapeutic Potential of Isoflavones with an Emphasis on Daidzein. Oxid Med. Cell Longev. 2021, 2021, 6331630. [Google Scholar] [CrossRef]
- Ekundayo, F.O.; Folorunsho, A.E.; Ibisanmi, T.A.; Olabanji, O.B. Antifungal activity of chitinase produced by Streptomyces species isolated from grassland soils in Futa Area, Akure. Bull. Natl. Res. Cent. 2022, 46, 95. [Google Scholar] [CrossRef]
- Legein, M.; Smets, W.; Vandenheuvel, D.; Eilers, T.; Muyshondt, B.; Prinsen, E.; Samson, R.; Lebeer, S. Modes of Action of Microbial Biocontrol in the Phyllosphere. Front. Microbiol. 2020, 11, 1619. [Google Scholar] [CrossRef]
- Daguerre, Y.; Siegel, K.; Edel-Hermann, V.; Steinberg, C. Fungal proteins and genes associated with biocontrol mechanisms of soil-borne pathogens: A review. Fungal Biol. Rev. 2014, 28, 97–125. [Google Scholar] [CrossRef]
- Nelkner, J.; Tejerizo, G.T.; Hassa, J.; Lin, T.W.; Witte, J.; Verwaaijen, B.; Winkler, A.; Bunk, B.; Spröer, C.; Overmann, J.; et al. Genetic Potential of the Biocontrol Agent Pseudomonas brassicacearum (Formerly P. trivialis) 3Re2-7 Unraveled by Genome Sequencing and Mining, Comparative Genomics and Transcriptomics. Genes 2019, 10, 601. [Google Scholar] [CrossRef] [PubMed]
- Funk, A.J.; McCullumsmith, R.E.; Haroutunian, V.; Meador-Woodruff, J.H. Abnormal activity of the MAPK- and cAMP-associated signaling pathways in frontal cortical areas in postmortem brain in schizophrenia. Neuropsychopharmacology 2012, 37, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Tomaž, Š.; Gruden, K.; Coll, A. TGA transcription factors—Structural characteristics as basis for functional variability. Front. Plant Sci. 2022, 13, 935819. [Google Scholar] [CrossRef]
- Liao, K.; Peng, Y.J.; Yuan, L.B.; Dai, Y.S.; Chen, Q.F.; Yu, L.J.; Bai, M.Y.; Zhang, W.Q.; Xie, L.J.; Xiao, S. Brassinosteroids Antagonize Jasmonate-Activated Plant Defense Responses through BRI1-EMS-SUPPRESSOR1 (BES1). Plant Physiol. 2020, 182, 1066–1082. [Google Scholar] [CrossRef]
- Kumar, M.; Brar, A.; Yadav, M.; Chawade, A.; Vivekanand, V.; Pareek, N. Chitinases—Potential Candidates for Enhanced Plant Resistance towards Fungal Pathogens. Agriculture 2018, 8, 88. [Google Scholar] [CrossRef]
- Trinh, T.H.T.; Nguyen, V.B.; Tran, D.M.; Doan, C.T.; Tran, T.N.; Wang, S.L.; Kosta, K.; Szkladanyi, S.; Le, M.H.; Nguyen, A.D. A Potent Fusarium Antagonistic Bacterium Bacillus subtilis RB.CJ41 Isolated from the Rhizosphere Roots of Black Pepper (Piper nigrum L.). unpublished manuscript.
- Petti, C.; Reiber, K.; Ali, S.S.; Berney, M.; Doohan, F.M. Auxin as a player in the biocontrol of Fusarium head blight disease of barley and its potential as a disease control agent. BMC Plant Biol. 2012, 12, 224. [Google Scholar] [CrossRef]
- Rabiey, M.; Hailey, L.E.; Roy, S.R.; Grenz, K.; Al-Zadjali, M.A.S.; Barrett, G.A.; Jackson, R.W. Endophytes vs tree pathogens and pests: Can they be used as biological control agents to improve tree health? Eur. J. Plant Pathol. 2019, 155, 711–729. [Google Scholar] [CrossRef]
- Kosawang, C.; Amby, D.B.; Bussaban, B.; McKinney, L.V.; Xu, J.; Kjær, E.D.; Collinge, D.B.; Nielsen, L.R. Fungal communities associated with species of Fraxinus tolerant to ash dieback, and their potential for biological control. Fungal Biol. 2018, 122, 110–120. [Google Scholar] [CrossRef]
- Halecker, S.; Wennrich, J.P.; Rodrigo, S.; Andrée, N.; Rabsch, L.; Baschien, C.; Steinert, M.; Standler, M.; Surup, F.; Schulz, B.J. Fungal endophytes for biocontrol of ash dieback: The antagonistic potential of Hypoxylon rubiginosum. Fungal Ecol. 2020, 45, 100918. [Google Scholar] [CrossRef]
- Steddom, K.; Becker, O.; Menge, J.A. Repetitive Applications of the Biocontrol Agent Pseudomonas putida 06909 and Effects on Populations of Phytophthora parasitica in Citrus Orchards. Phytopathology 2002, 92, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Acebo-Guerrero, Y.; Hernández-Rodríguez, A.; Vandeputte, O.; Miguélez-Sierra, Y.; Heydrich-Pérez, M.; Ye, L.; Cornelis, P.; Bertin, P.; El Jaziri, M. Characterization of Pseudomonas chlororaphis from Theobroma cacao L. rhizosphere with antagonistic activity against Phytophthora palmivora (Butler). J. Appl. Microbiol. 2015, 119, 1112–1126. [Google Scholar] [CrossRef]
- Lee, B.D.; Dutta, S.; Ryu, H.; Yoo, S.J.; Suh, D.S.; Park, K. Induction of systemic resistance in Panax ginseng against Phytophthora cactorum by native Bacillus amyloliquefaciens HK34. J. Ginseng. Res. 2015, 39, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Mpika, J.; Kébé, I.B.; Issali, A.E.; N’Guessan, F.K.; Druzhinina, S.; Komon-Zélazowska, M.; Kubicek, C.P.; Aké, S. Antagonist potential of Trichoderma indigenous isolates for biological control of Phytophthora palmivora the causative agent of black pod disease on cocoa (Theobroma cacao L.) in Côte d’Ivoire. Afr. J. Biotechnol. 2009, 8, 5280–5293. [Google Scholar]
- Diánez Martínez, F.; Santos, M.; Carretero, F.; Marín, F. Trichoderma saturnisporum, a new biological control agent. J. Sci. Food Agric. 2016, 96, 1934–1944. [Google Scholar] [CrossRef]
- Abbas, A.; Mubeen, M.; Zheng, H.; Sohail, M.A.; Shakeel, Q.; Solanki, M.K.; Iftikhar, Y.; Sharma, S.; Kashyap, B.K.; Hussain, S.; et al. Trichoderma spp. Genes Involved in the Biocontrol Activity against Rhizoctonia solani. Front. Microbiol. 2022, 13, 884469. [Google Scholar] [CrossRef]
- Palmieri, D.; Ianiri, G.; Del Grosso, C.; Barone, G.; De Curtis, F.; Castoria, R.; Lima, G. Advances and Perspectives in the Use of Biocontrol Agents against Fungal Plant Diseases. Horticulturae 2022, 8, 577. [Google Scholar] [CrossRef]
- Hernando José, B.-A.; González-Rodríguez, V.E.; Almeida, G.R.; Izquierdo-Bueno, I.; Moraga, J.; Carbú, M.; Cantoral, J.M.; Garrido, C. Endophytic Microorganisms as an Alternative for the Biocontrol of Phytophthora spp. In Agro-Economic Risks of Phytophthora and an Effective Biocontrol Approach; Waleed Mohamed Hussain, A., Ed.; IntechOpen: Rijeka, Croatia, 2021; pp. 1–12. [Google Scholar]
- Naidoo, S.; Slippers, B.; Plett, J.M.; Coles, D.; Oates, C.N. The Road to Resistance in Forest Trees. Front. Plant Sci. 2019, 10, 273. [Google Scholar] [CrossRef]
- Moricca, S.; Panzavolta, T. Recent Advances in the Monitoring, Assessment and Management of Forest Pathogens and Pests. Forests 2021, 12, 1623. [Google Scholar]
- Rabiey, M.; Welch, T.; Sanchez-Lucas, R.; Stevens, K.; Raw, M.; Kettles, G.J.; Catoni, M.; McDonald, M.C.; Jackson, R.W.; Luna, E. Scaling-up to understand tree–pathogen interactions: A steep, tough climb or a walk in the park? Curr. Opin. Plant Biol. 2022, 68, 102229. [Google Scholar]
- Chandelier, A.; Husson, C.; Druart, P.; Marçais, B. Assessment of inoculation methods for screening black alder resistance to Phytophthora× alni. Plant Pathol. 2016, 65, 441–450. [Google Scholar]
- Sniezko, R.A.; Koch, J. Breeding trees resistant to insects and diseases: Putting theory into application. Biol. Invasions 2017, 19, 3377–3400. [Google Scholar]
- Woodcock, P.; Marzano, M.; Quine, C.P. Key lessons from resistant tree breeding programmes in the Northern Hemisphere. Ann. For. Sci. 2019, 76, 51. [Google Scholar]
- Pike, C.C.; Koch, J.; Nelson, C.D. Breeding for Resistance to Tree Pests: Successes, Challenges, and a Guide to the Future. J. For. 2020, 119, 96–105. [Google Scholar]
- Wei, Z.; Jousset, A. Plant Breeding Goes Microbial. Trends Plant Sci. 2017, 22, 555–558. [Google Scholar] [PubMed]
- Marco, S.; Loredana, M.; Riccardo, V.; Raffaella, B.; Walter, C.; Luca, C. Microbe-assisted crop improvement: A sustainable weapon to restore holobiont functionality and resilience. Hortic. Res. 2022, 9, uhac160. [Google Scholar]
- Bubner, B. Resistant alder; Selection of Black alder (Alnus glutinosa (L.) resistant against Phytophthora alni. Thünen Institute. 2018. Available online: https://www.thuenen.de/en/institutes/forest-genetics/research-groups/pathogen-resistance-and-seed-quality-research/completed-projects/breeding-of-black-alder (accessed on 26 March 2023).
- Rathore, R.; Vakharia, D.N.; Rathore, D.S. In vitro screening of different Pseudomonas fluorescens isolates to study lytic enzyme production and growth inhibition during antagonism of Fusarium oxysporum f. sp. cumini, wilt causing pathogen of cumin. Egypt. J. Biol. Pest Control 2020, 30, 57. [Google Scholar]
- Ganeshan, G.; Manoj Kumar, A. Pseudomonas fluorescens, a potential bacterial antagonist to control plant diseases. J. Plant Interact. 2005, 1, 123–134. [Google Scholar]
- Foo, J.L.; Ling, H.; Lee, Y.S.; Chang, M.W. Microbiome engineering: Current applications and its future. Biotechnol. J. 2017, 12, 1600099. [Google Scholar]
- Hu, H.; Wang, M.; Huang, Y.; Xu, Z.; Xu, P.; Nie, Y.; Tang, H. Guided by the principles of microbiome engineering: Accomplishments and perspectives for environmental use. mLife 2022, 1, 382–398. [Google Scholar]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; van der Voort, M.; Schneider, J.H.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A.; et al. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [PubMed]
- Santhanam, R.; Luu, V.T.; Weinhold, A.; Goldberg, J.; Oh, Y.; Baldwin, I.T. Native root-associated bacteria rescue a plant from a sudden-wilt disease that emerged during continuous cropping. Proc. Natl. Acad Sci. USA 2015, 112, E5013–E5020. [Google Scholar] [PubMed]
- Mukherjee, A.; Singh, S.; Gaurav, A.K.; KumarChouhan, G.; Jaiswal, D.K.; Pereira, A.P.A.; KumarPassari, A.; Abdel-Azeem, A.; Verma, J.P. Harnessing of phytomicrobiome for developing potential bio stimulant consortium for enhancing the productivity of chickpea and soil health under sustainable agriculture. Sci. Total Environ. 2022, 836, 2–11. [Google Scholar]
- Wicaksono, W.A.; Jones, E.; Casonato, S.; Monk, J.; Ridgway, H.J. Biological control of Pseudomonas syringae pv. actinidiae (Psa), the causal agent of bacterial canker of kiwifruit, using endophytic bacteria recovered from a medicinal plant. Biol. Control 2018, 116, 103–112. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuller, E.; Germaine, K.J.; Rathore, D.S. The Good, the Bad, and the Useable Microbes within the Common Alder (Alnus glutinosa) Microbiome—Potential Bio-Agents to Combat Alder Dieback. Microorganisms 2023, 11, 2187. https://doi.org/10.3390/microorganisms11092187
Fuller E, Germaine KJ, Rathore DS. The Good, the Bad, and the Useable Microbes within the Common Alder (Alnus glutinosa) Microbiome—Potential Bio-Agents to Combat Alder Dieback. Microorganisms. 2023; 11(9):2187. https://doi.org/10.3390/microorganisms11092187
Chicago/Turabian StyleFuller, Emma, Kieran J. Germaine, and Dheeraj Singh Rathore. 2023. "The Good, the Bad, and the Useable Microbes within the Common Alder (Alnus glutinosa) Microbiome—Potential Bio-Agents to Combat Alder Dieback" Microorganisms 11, no. 9: 2187. https://doi.org/10.3390/microorganisms11092187