
Fungal Community Composition at the Last Remaining Wild Site of Yellow Early Marsh Orchid (Dactylorhiza incarnata ssp. ochroleuca)
 and
and
Abstract
:1. Introduction
2. Methods
2.1. Sampling Site
2.2. Soil Processing and Chemical Analyses
2.3. Soil DNA Extraction
2.4. Metabarcoding
2.5. Statistical Analysis
3. Results
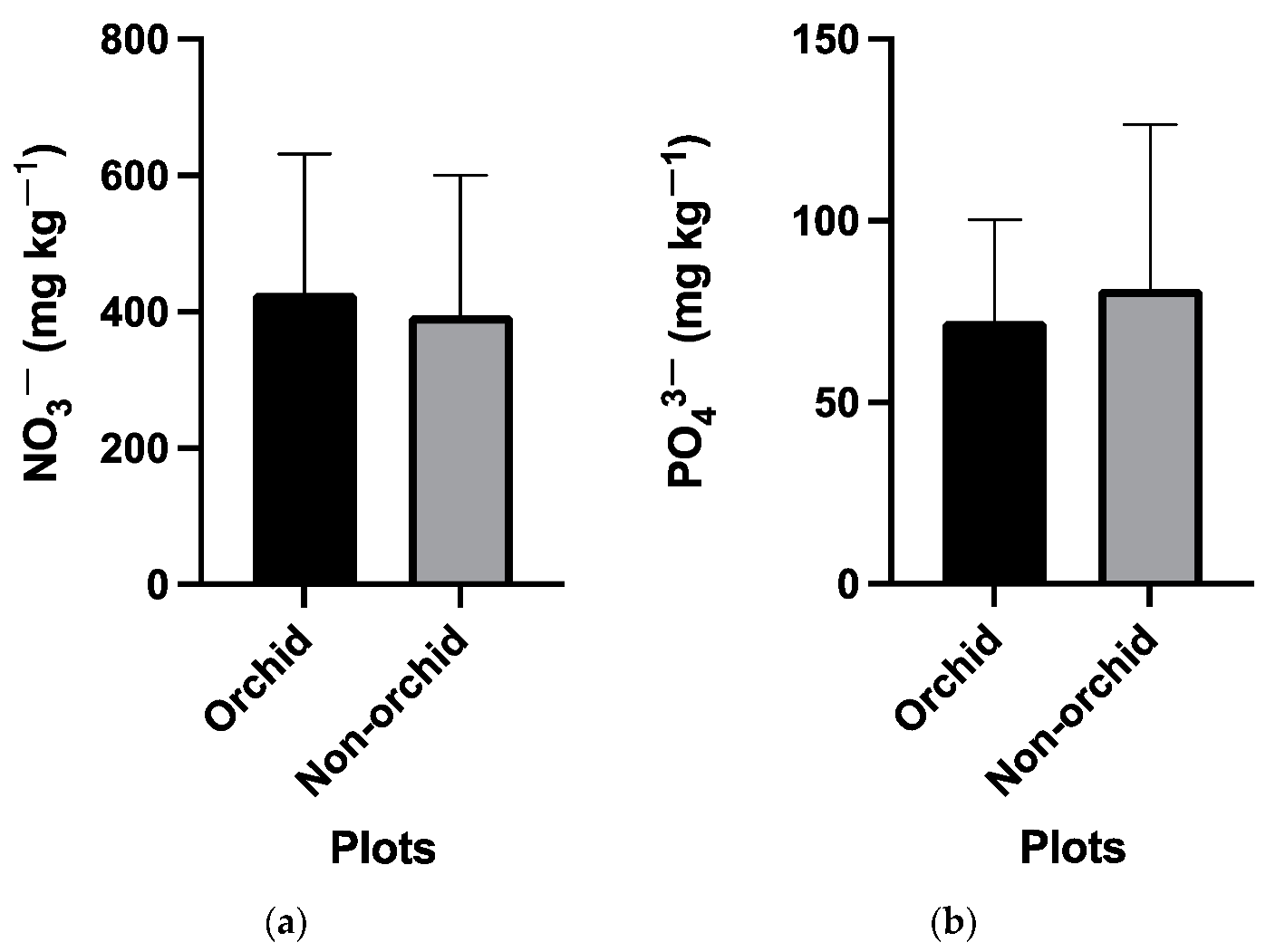
3.1. Abiotic Soil Characteristics
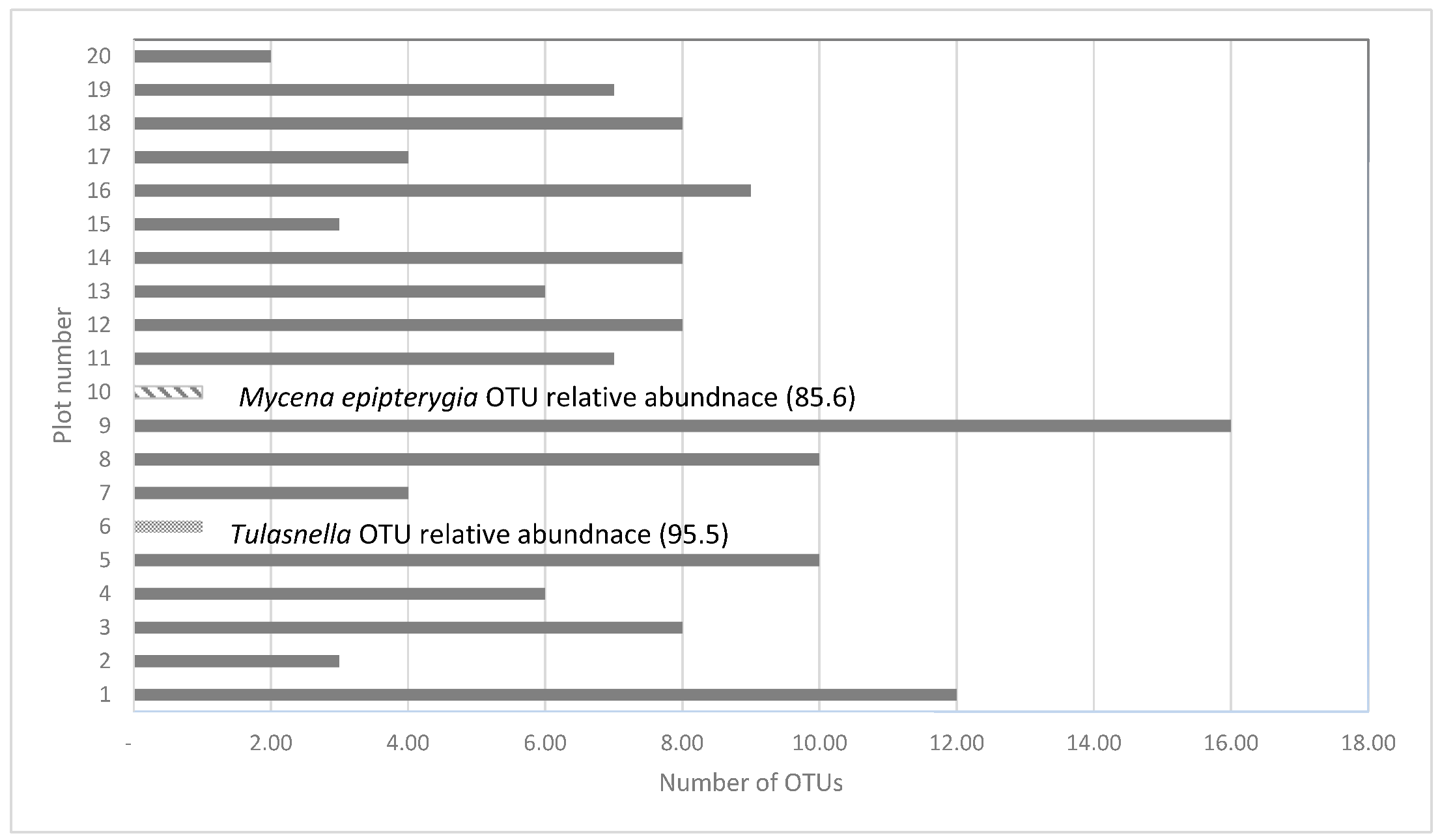
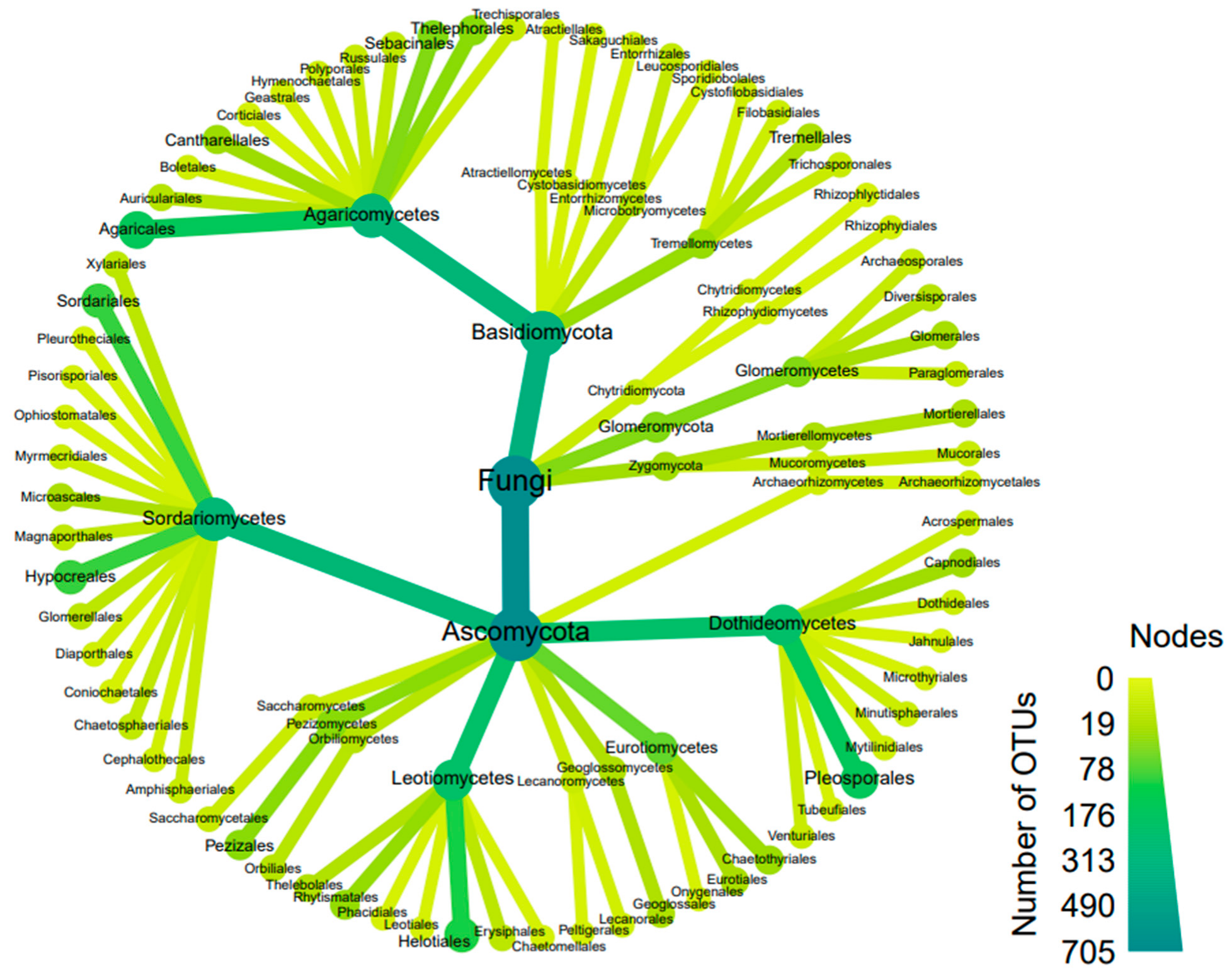
3.2. Soil DNA Concentrations and Fungal Community Composition by Plot
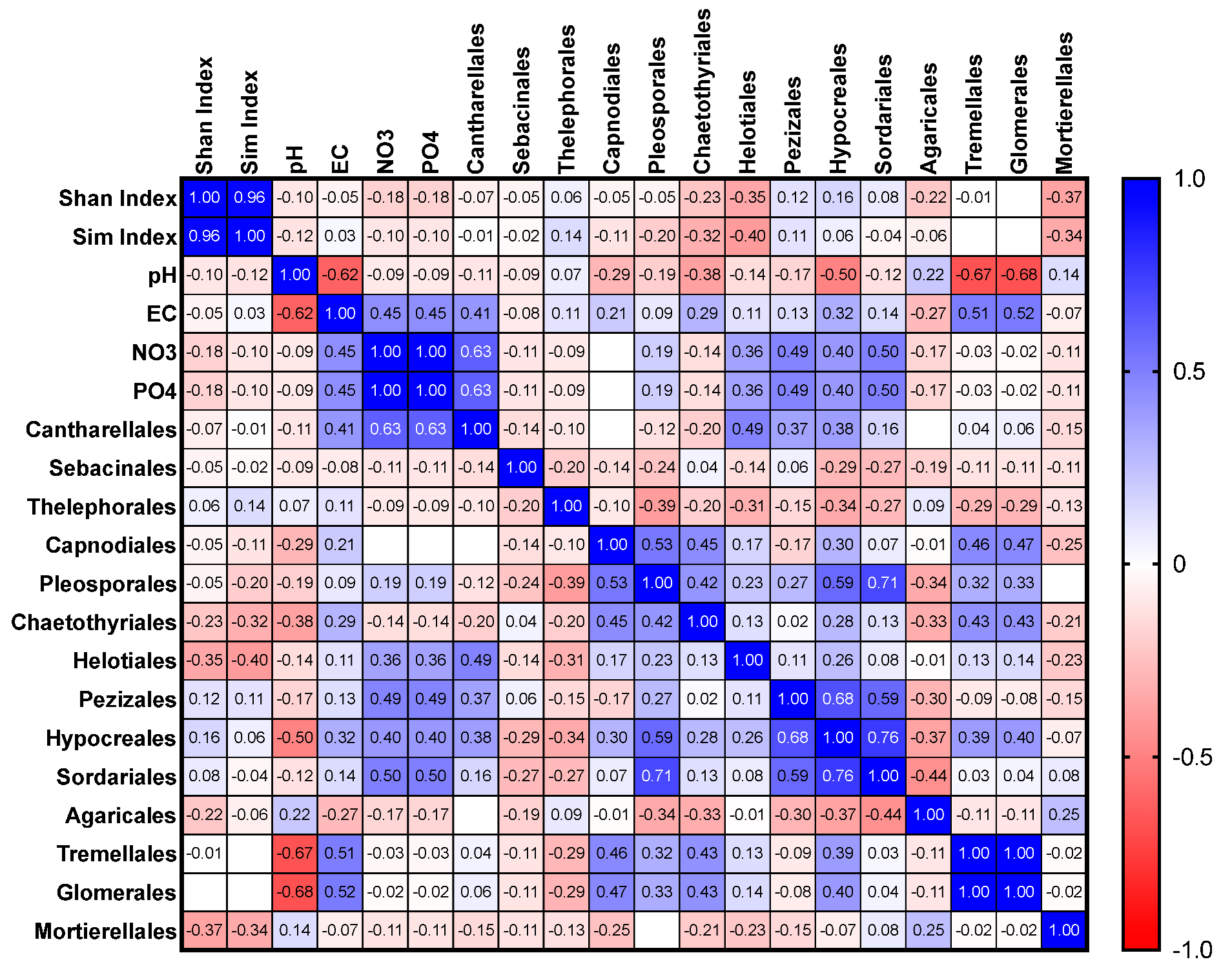
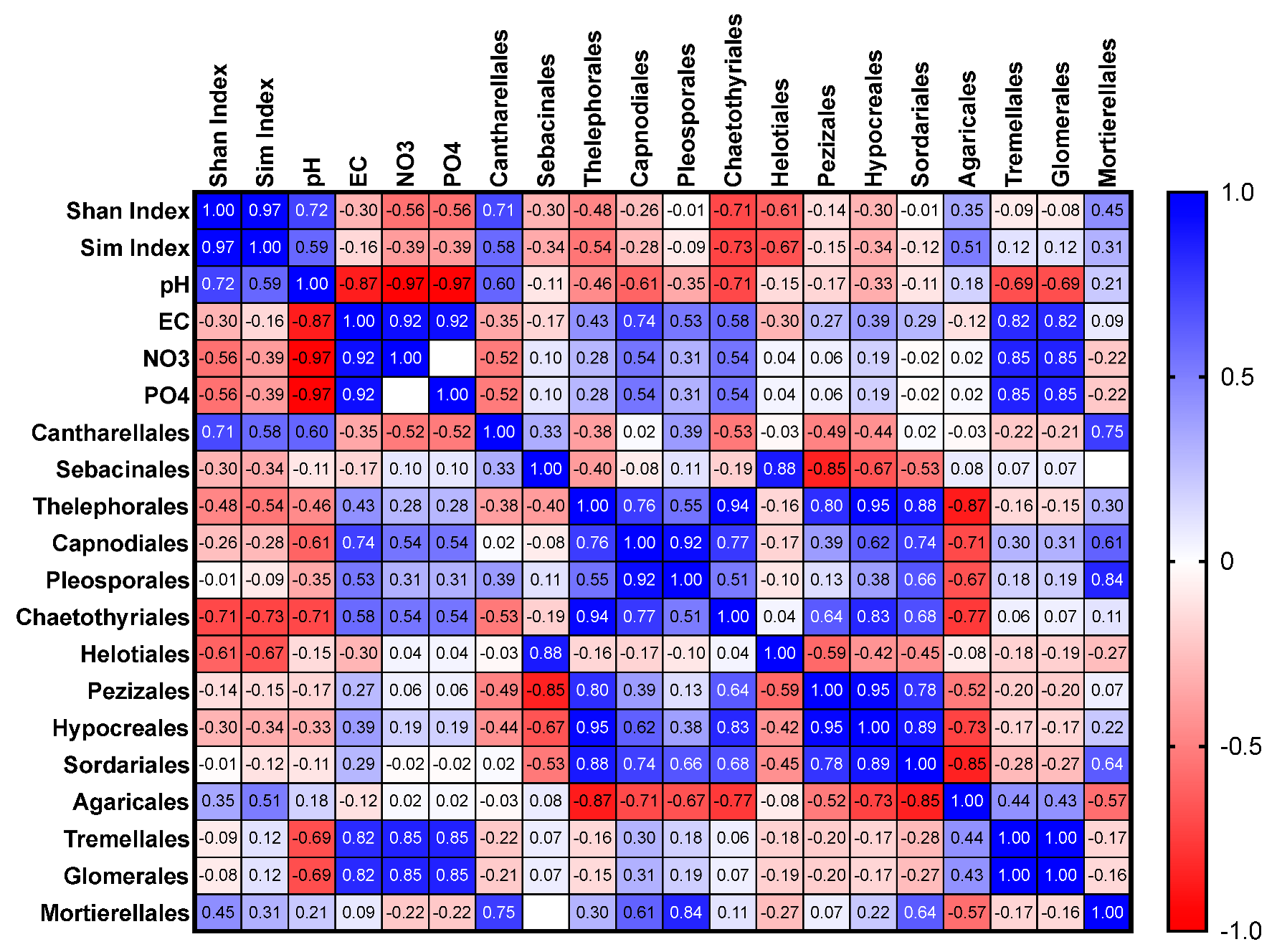
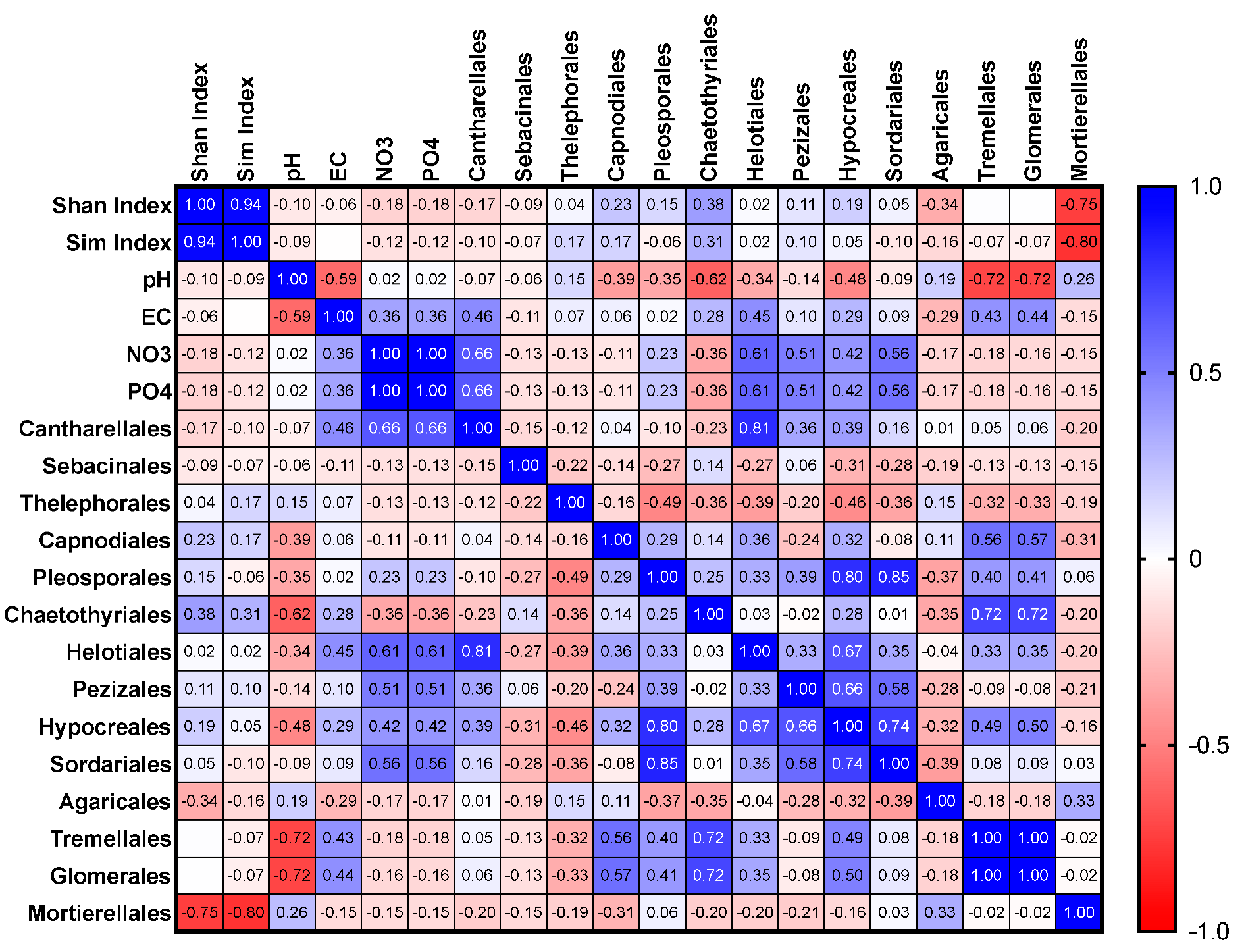
3.3. Relationship between Soil Characteristics and Fungal Distribution and Relative Abundance
3.3.1. All Plots
3.3.2. Orchid-Hosting Plots Only

3.3.3. Non-Orchid-Hosting Plots

4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Swarts, N.D.; Dixon, K.W. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Willmer, P. Climate Change: Bees and Orchids Lose Touch. Curr. Biol. 2014, 24, R1133–R1135. [Google Scholar] [CrossRef] [PubMed]
- Fay, M.F. British and Irish orchids in a changing world. Curtis’s Bot. Mag. 2015, 32, 3–23. [Google Scholar] [CrossRef]
- Yokoya, K.; Zettler, L.W.; Kendon, J.P.; Bidartondo, M.I.; Stice, A.L.; Skarha, S.; Corey, L.L.; Knight, A.C.; Sarasan, V. Preliminary findings on identification of mycorrhizal fungi from diverse orchids in the Central Highlands of Madagascar. Mycorrhiza 2015, 25, 611–625. [Google Scholar] [CrossRef]
- Liu, H.; Feng, C.-L.; Luo, Y.-B.; Chen, B.-S.; Wang, Z.-S.; Gu, H.-Y. Potential Challenges of Climate Change to Orchid Conservation in a Wild Orchid Hotspot in Southwestern China. Bot. Rev. 2010, 76, 174–192. [Google Scholar] [CrossRef]
- Phillips, R.D.; Reiter, N.; Peakall, R. Orchid conservation: From theory to practice. Ann. Bot. 2020, 126, 345–362. [Google Scholar] [CrossRef]
- McCormick, M.K.; Jacquemyn, H. What constrains the distribution of orchid populations? New Phytol. 2014, 202, 392–400. [Google Scholar] [CrossRef]
- Bidartondo, M.I.; Read, D.J. Fungal specificity bottlenecks during orchid germination and development. Mol. Ecol. 2008, 17, 3707–3716. [Google Scholar] [CrossRef]
- Kendon, J.P.; Yokoya, K.; Zettler, L.W.; Jacob, A.S.; McDiarmid, F.; Bidartondo, M.I.; Sarasan, V. Recovery of mycorrhizal fungi from wild collected protocorms of Madagascan endemic orchid Aerangis ellisii (B.S. Williams) Schltr. and their use in seed germination in vitro. Mycorrhiza 2020, 30, 567–576. [Google Scholar] [CrossRef]
- Vogt-Schilb, H.; Těšitelová, T.; Kotilínek, M.; Sucháček, P.; Kohout, P.; Jersáková, J. Altered rhizoctonia assemblages in grasslands on ex-arable land support germination of mycorrhizal generalist, not specialist orchids. New Phytol. 2020, 227, 1200–1212. [Google Scholar] [CrossRef]
- Rasmussen, H.N.; Rasmussen, F.N. Seedling mycorrhiza: A discussion of origin and evolution in Orchidaceae. Bot. J. Linn. Soc. 2014, 175, 313–327. [Google Scholar] [CrossRef]
- Fay, M.F. Orchid conservation: How can we meet the challenges in the twenty-first century? Bot. Stud. 2018, 59, 16. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, E.; Rillig, M.C.; Wehner, J.; Hegglin, D.; Wittwer, R.; Heijden, M.G.A. Sebacinales, but not total root associated fungal communities, are affected by land-use intensity. New Phytol. 2014, 203, 1036–1040. [Google Scholar] [CrossRef]
- Gerz, M.; Bueno, C.G.; Ozinga, W.A.; Zobel, M.; Moora, M. Responses of plant community mycorrhization to anthropogenic influence depend on the habitat and mycorrhizal type. Oikos 2019, 128, 1565–1575. [Google Scholar] [CrossRef]
- Lambers, H.; Martinoia, E.; Renton, M. Plant adaptations to severely phosphorus-impoverished soils. Curr. Opin. Plant Biol. 2015, 25, 23–31. [Google Scholar] [CrossRef]
- Kaur, J.; Phillips, C.; Sharma, J. Host population size is linked to orchid mycorrhizal fungal communities in roots and soil, which are shaped by microenvironment. Mycorrhiza 2020, 31, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Cavagnaro, T.R. Soil moisture legacy effects: Impacts on soil nutrients, plants and mycorrhizal responsiveness. Soil Biol. Biochem. 2016, 95, 173–179. [Google Scholar] [CrossRef]
- Mujica, M.I.; Saez, N.; Cisternas, M.; Manzano, M.; Armesto, J.J.; Pérez, F. Relationship between soil nutrients and mycorrhizal associations of two Bipinnula species (Orchidaceae) from central Chile. Ann. Bot. 2016, 118, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Bunch, W.D.; Cowden, C.C.; Wurzburger, N.; Shefferson, R.P. Geography and soil chemistry drive the distribution of fungal associations in lady’s slipper orchid, Cypripedium acaule. Botany 2013, 91, 850–856. [Google Scholar] [CrossRef]
- Bell, J.; Yokoya, K.; Kendon, J.P.; Sarasan, V. Diversity of root-associated culturable fungi of Cephalanthera rubra (Orchidaceae) in relation to soil characteristics. PeerJ 2020, 8, e8695. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Waud, M.; Merckx, V.S.F.T.; Brys, R.; Tyteca, D.; Hedrén, M.; Lievens, B. Habitat-driven variation in mycorrhizal communities in the terrestrial orchid genus Dactylorhiza. Sci. Rep. 2016, 6, 37182. [Google Scholar] [CrossRef]
- Mújica, E.B.; Mably, J.J.; Skarha, S.M.; Corey, L.L.; Richardson, L.W.; Danaher, M.W.; González, E.H.; Zettler, L.W. A comparision of ghost orchid (Dendrophylax lindenii) habitats in Florida and Cuba, with particular reference to seedling recruitment and mycorrhizal fungi. Bot. J. Linn. Soc. 2018, 186, 572–586. [Google Scholar] [CrossRef]
- Illyés, Z.; Halász, K.; Rudnóy, S.; Ouanphanivanh, N.; Garay, T.; Bratek, Z. Changes in the Diversity of the Mycorrhizal Fungi of Orchids as a Function of the Water Supply of the Habitat. J. Appl. Bot. Food Qual. 2010, 83, 28–36. [Google Scholar]
- Zettler, L.W.; Piskin, K.A. Mycorrhizal Fungi from Protocorms, Seedlings and Mature Plants of the Eastern Prairie Fringed Orchid, Platanthera leucophaea (Nutt.) Lindley: A Comprehensive List to Augment Conservation. Am. Midl. Nat. 2011, 166, 29–39. [Google Scholar] [CrossRef]
- Reiter, N.; Lawrie, A.C.; Linde, C.C. Matching symbiotic associations of an endangered orchid to habitat to improve conservation outcomes. Ann. Bot. 2018, 122, 947–959. [Google Scholar] [CrossRef]
- Downing, J.L.; Liu, H.; Shao, S.; Wang, X.; McCormick, M.; Deng, R.; Gao, J. Contrasting changes in biotic interactions of orchid populations subject to conservation introduction vs. conventional translocation in tropical China. Biol. Conserv. 2017, 212, 29–38. [Google Scholar] [CrossRef]
- Sarasan, V.; Pankhurst, T.; Yokoya, K.; Sriskandarajah, S.; McDiarmid, F. Preventing Extinction of the Critically Endangered Dactylorhiza incarnata subsp. ochroleuca in Britain Using Symbiotic Seedlings for Reintroduction. Microorganisms 2021, 9, 1421. [Google Scholar] [CrossRef]
- Lindahl, B.D.; Nilsson, R.H.; Tedersoo, L.; Abarenkov, K.; Carlsen, T.; Kjøller, R.; Kõljalg, U.; Pennanen, T.; Rosendahl, S.; Stenlid, J.; et al. Fungal community analysis by high-throughput sequencing of amplified markers—A user’s guide. New Phytol. 2013, 199, 288–299. [Google Scholar] [CrossRef]
- Hofmann, B.; Dreyling, L.; Grande, F.D.; Otte, J.; Schmitt, I. Habitat and tree species identity shape aboveground and belowground fungal communities in central European forests. Front. Microbiol. 2023, 14, 1067906. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee SJ, W.T.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Pro-Tocols: A Guide to Methods and Applications, 315th–322nd ed.; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; De Wit, P.; Sánchez-García, M.; Ebersberger, I.; de Sousa, F.; et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013, 4, 914–919. [Google Scholar] [CrossRef]
- Edgar, R.C. UNOISE2: Improved error-correction for Illumina 16S and ITS amplicon sequencing. bioRxiv 2016, 081257. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applica-tions. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- McCormick, M.K.; Taylor, D.L.; Juhaszova, K.; Burnett, R.K., Jr.; Whigham, D.F.; O’Neill, J.P. Limitations on orchid recruitment: Not a simple picture. Mol. Ecol. 2012, 21, 1511–1523. [Google Scholar] [CrossRef]
- Dickie, I.A.; Martínez-García, L.B.; Koele, N.; Grelet, G.-A.; Tylianakis, J.M.; Peltzer, D.A.; Richardson, S.J. Mycorrhizas and mycorrhizal fungal communities throughout ecosystem development. Plant Soil 2013, 367, 11–39. [Google Scholar] [CrossRef]
- Voyron, S.; Ercole, E.; Ghignone, S.; Perotto, S.; Girlanda, M. Fine-scale spatial distribution of orchid mycorrhizal fungi in the soil of host-rich grasslands. New Phytol. 2016, 213, 1428–1439. [Google Scholar] [CrossRef]
- Li, T.; Wu, S.; Yang, W.; Selosse, M.-A.; Gao, J. How Mycorrhizal Associations Influence Orchid Distribution and Population Dynamics. Front. Plant Sci. 2021, 12, 647114. [Google Scholar] [CrossRef]
- Janissen, B.; French, G.; Selby-Pham, J.; Lawrie, A.C.; Huynh, T. Differences in emergence and flowering in wild, re-introduced and translocated populations of an endangered terrestrial orchid and the influences of climate and orchid mycorrhizal abundance. Aust. J. Bot. 2021, 69, 9. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Deja, A.; De Hert, K.; Cachapa Bailarote, B.; Lievens, B. Variation in Mycorrhizal Associations with Tulasnelloid Fungi among Populations of Five Dactylorhiza Species. PLoS ONE 2012, 7, e42212. [Google Scholar] [CrossRef]
- Waud, M.; Busschaert, P.; Lievens, B.; Jacquemyn, H. Specificity and localised distribution of mycorrhizal fungi in the soil may contribute to co-existence of orchid species. Fungal Ecol. 2016, 20, 155–165. [Google Scholar] [CrossRef]
- Yokoya, K.; Zettler, L.W.; Bell, J.; Kendon, J.P.; Jacob, A.S.; Schofield, E.; Rajaovelona, L.; Sarasan, V. The Diverse Assemblage of Fungal Endophytes from Orchids in Madagascar Linked to Abiotic Factors and Seasonality. Diversity 2021, 13, 96. [Google Scholar] [CrossRef]
- Han, J.Y.; Xiao, H.; Gao, J. Seasonal dynamics of mycorrhizal fungi in Paphiopedilum spicerianum (Rchb. f) Pfitzer—A critically endangered orchid from China. Glob. Ecol. Conserv. 2016, 6, 327–338. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | pH | EC (µS cm−1) | PO4 (mg kg−1) | NO3 (mg kg−1) | D | H | DNA Yield (ng µL−1) |
|---|---|---|---|---|---|---|---|
| 1 | 7.65 | 573 | 15.1 | 75.51 | 0.90 | 2.62 | 84 |
| 2 | 7.98 | 312 | 36.18 | 180.91 | 0.50 | 0.93 | 70 |
| 3 | 7.87 | 470 | 95.97 | 479.86 | 0.87 | 2.27 | 78 |
| 4 | 7.5 | 479.33 | 74.82 | 374.1 | 0.38 | 0.96 | 90 |
| 5 | 7.81 | 791 | 71.47 | 357.33 | 0.83 | 2.09 | 99.1 |
| 6 | 7.82 | 830.67 | 70.48 | 352.4 | 0.86 | 2.18 | 48.8 |
| 7 | 7.54 | 1143.67 | 188.92 | 944.59 | 0.65 | 1.44 | 87 |
| 8 | 7.64 | 954.33 | 102.89 | 514.47 | 0.20 | 0.57 | 103 |
| 9 | 7.58 | 634.67 | 74.7 | 373.52 | 0.62 | 1.38 | 42.1 |
| 10 | 7.44 | 753.33 | 66.25 | 331.27 | 0.79 | 1.58 | 164 |
| 11 | 7.63 | 968.33 | 113.27 | 566.35 | 0.50 | 0.86 | 57 |
| 12 | 7.85 | 566.33 | 53.17 | 265.87 | 0.85 | 2.41 | 95 |
| 13 | 7.76 | 418 | 65.96 | 329.82 | 0.04 | 0.12 | 140 |
| 14 | 7.88 | 397.33 | 43.01 | 215.03 | 0.72 | 1.86 | 118 |
| 15 | 7.67 | 829.33 | 86.08 | 430.39 | 0.08 | 0.24 | 76 |
| 16 | 7.64 | 745 | 151.48 | 757.42 | 0.86 | 2.13 | 82 |
| 17 | 7.52 | 655.33 | 120.1 | 600.49 | 0.88 | 2.41 | 104 |
| 18 | 7.43 | 636.67 | 57.13 | 285.66 | 0.88 | 2.66 | 104 |
| 19 | 7.06 | 1271.33 | 44.59 | 222.95 | 0.80 | 1.89 | 71 |
| 20 | 7.53 | 659.33 | 48.88 | 244.4 | 0.92 | 2.80 | 91 |
| Sample ID | Class | Order | Family | Genus | Taxa (Species) |
|---|---|---|---|---|---|
| 1 | 12 | 33 | 49 | 49 | 173 (20) |
| 2 | 8 | 16 | 24 | 13 | 74 (4) |
| 3 | 11 | 21 | 31 | 25 | 103 (8) |
| 4 | 12 | 38 | 60 | 49 | 207 (26) |
| 5 | 7 | 19 | 22 | 17 | 48 (4) |
| 6 | 4 | 5 | 5 | 4 | 10 (0) |
| 7 | 11 | 36 | 67 | 59 | 236 (25) |
| 8 | 12 | 30 | 42 | 35 | 149 (10) |
| 9 | 10 | 23 | 29 | 23 | 74 (9) |
| 10 | 8 | 17 | 25 | 21 | 82 (10) |
| 11 | 12 | 29 | 39 | 34 | 151 (13) |
| 12 | 10 | 24 | 35 | 36 | 139 (14) |
| 13 | 8 | 19 | 31 | 22 | 99 (5) |
| 14 | 12 | 32 | 39 | 33 | 138 (13) |
| 15 | 12 | 29 | 43 | 35 | 164 (14) |
| 16 | 10 | 25 | 38 | 32 | 146 (12) |
| 17 | 12 | 29 | 43 | 38 | 151 (15) |
| 18 | 12 | 31 | 47 | 44 | 178 (16) |
| 19 | 11 | 25 | 41 | 42 | 171 (22) |
| 20 | 13 | 30 | 44 | 40 | 172 (18) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dove, A.; Charters, M.D.; Campbell, M.J.; Blake, H.; Menon, M.; Sarasan, V. Fungal Community Composition at the Last Remaining Wild Site of Yellow Early Marsh Orchid (Dactylorhiza incarnata ssp. ochroleuca). Microorganisms 2023, 11, 2124. https://doi.org/10.3390/microorganisms11082124
Dove A, Charters MD, Campbell MJ, Blake H, Menon M, Sarasan V. Fungal Community Composition at the Last Remaining Wild Site of Yellow Early Marsh Orchid (Dactylorhiza incarnata ssp. ochroleuca). Microorganisms. 2023; 11(8):2124. https://doi.org/10.3390/microorganisms11082124
Chicago/Turabian StyleDove, Andrea, Michael D. Charters, Matthew J. Campbell, Hanna Blake, Manoj Menon, and Viswambharan Sarasan. 2023. "Fungal Community Composition at the Last Remaining Wild Site of Yellow Early Marsh Orchid (Dactylorhiza incarnata ssp. ochroleuca)" Microorganisms 11, no. 8: 2124. https://doi.org/10.3390/microorganisms11082124