
CRISPR-Cas-Based Adaptive Immunity Mediates Phage Resistance in Periodontal Red Complex Pathogens
 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Neural Network
2.2. SVM
2.3. Random Forest
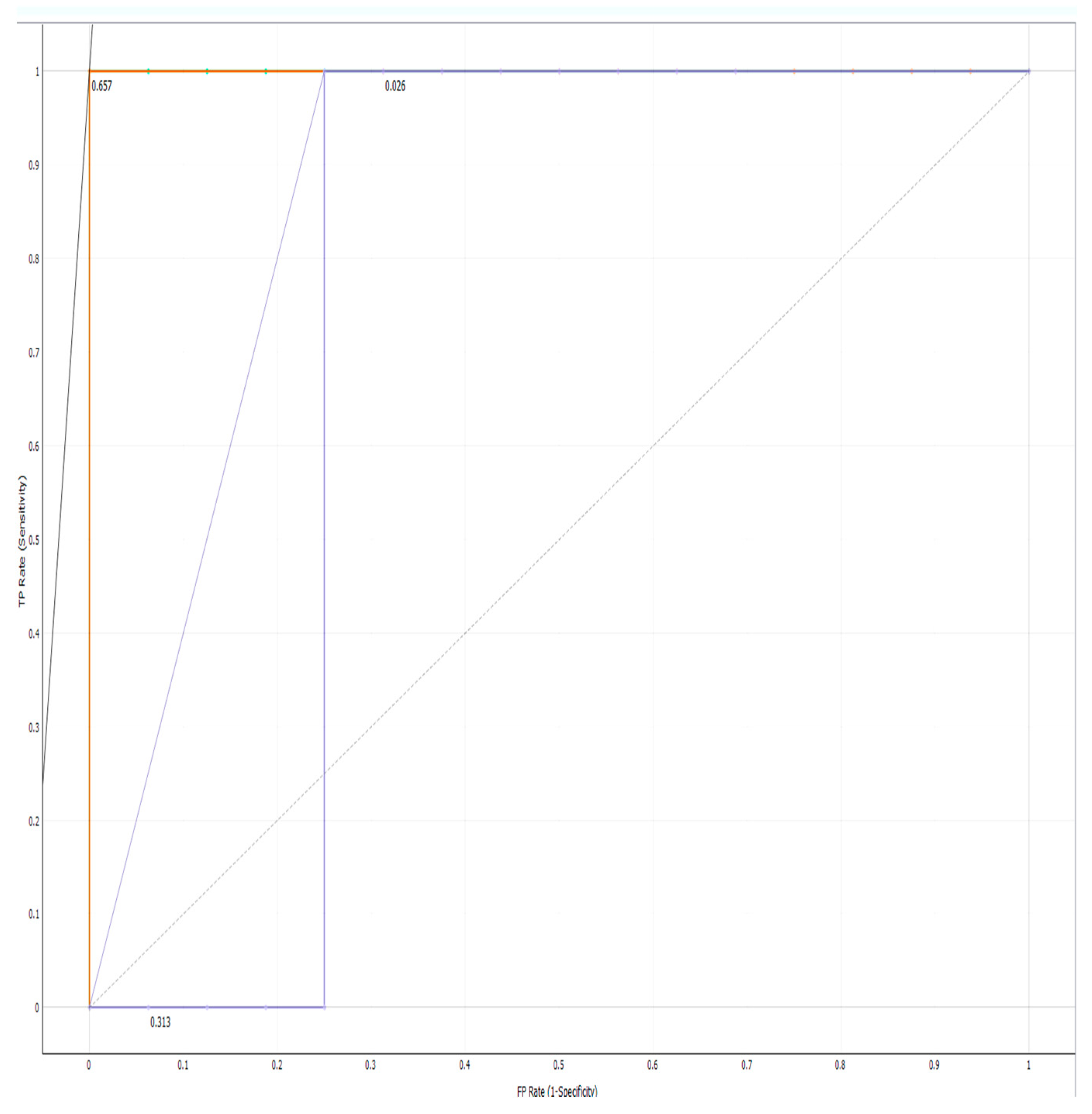
2.4. AUC-ROC Curve
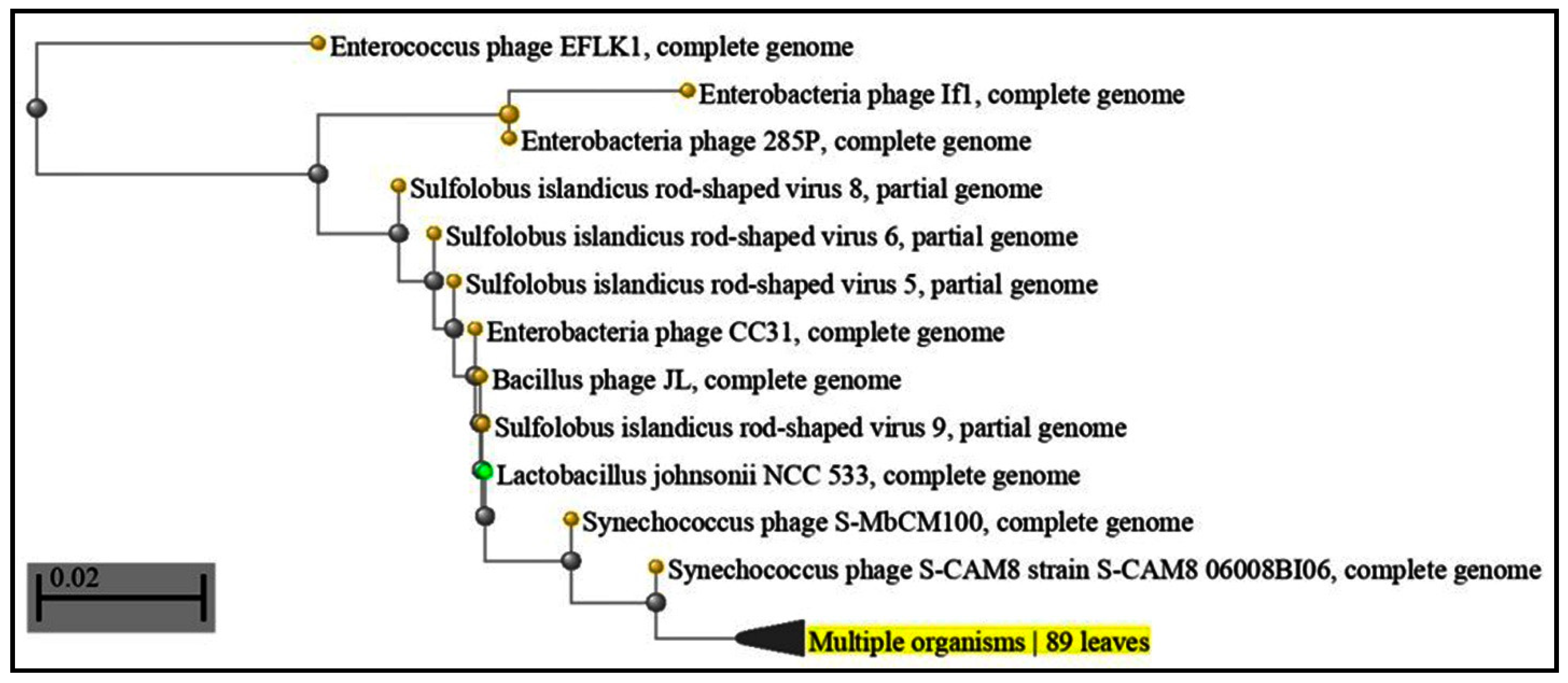
3. Results
3.1. Identification of CRISPR in P. gingivalis
3.2. Treponema Denticola CRISPR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sela, M.N. Role of Treponema denticola in periodontal diseases. Crit. Rev. Oral Biol. Med. 2001, 12, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Könönen, E.; Gursoy, M.; Gursoy, U.K. Periodontitis: A multifaceted disease of tooth-supporting tissues. J. Clin. Med. 2019, 8, 1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bibi, T.; Khurshid, Z.; Rehman, A.; Imran, E.; Srivastava, K.C.; Shrivastava, D. Gingival crevicular fluid (GCF): A diagnostic tool for the detection of periodontal health and diseases. Molecules 2021, 26, 1208. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, D.; Srivastava, K.C.; Ganji, K.K.; Alam, M.K.; Al Zoubi, I.; Sghaireen, M.G. Quantitative assessment of gingival inflammation in patients undergoing nonsurgical periodontal therapy using photometric CIELab analysis. BioMed Res. Int. 2021, 2021, 6615603. [Google Scholar] [CrossRef]
- Ximénez-Fyvie, L.A.; Haffajee, A.D.; Socransky, S.S. Microbial composition of supra-and subgingival plaque in subjects with adult periodontitis. J. Clin. Periodontol. 2000, 27, 722–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrivastava, D.; Srivastava, K.C.; Dayakara, J.K.; Sghaireen, M.G.; Gudipaneni, R.K.; Al-Johani, K.; Baig, M.N.; Khurshid, Z. BactericidalActivity of Crevicular Polymorphonuclear Neutrophils in Chronic Periodontitis Patients and Healthy Subjects under the Influence of Areca Nut Extract: An In Vitro Study. Appl. Sci. 2020, 10, 5008. [Google Scholar] [CrossRef]
- Assiri, K.I.; Sandeepa, N.C.; Asiri, R.S.; Mulawi, S.A.; Najmi, S.M.; Srivastava, K.C. Assessment of Oral–Systemic Disease Association amongst Dental Patients: A Retrospective Panoramic Radiographic Study. J. Contemp. Dent. Pract. 2020, 21, 748–755. [Google Scholar] [PubMed]
- Shrivastava, D.; Natoli, V.; Srivastava, K.C.; Alzoubi, I.A.; Nagy, A.I.; Hamza, M.O.; Al-Johani, K.; Alam, M.K.; Khurshid, Z. Novel approach to dental biofilm management through guided biofilm therapy (Gbt): A review. Microorganisms 2021, 9, 1966. [Google Scholar] [CrossRef]
- Lin, D.M.; Koskella, B.; Lin, H.C. Phage therapy: An alternative to antibiotics in the age of multi-drug resistance. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 162. [Google Scholar] [CrossRef]
- Gummalla, V.S.; Zhang, Y.; Liao, Y.-T.; Wu, V.C. The Role of Temperate Phages in Bacterial Pathogenicity. Microorganisms 2023, 11, 541. [Google Scholar] [CrossRef]
- Pinto, G.; Silva, M.D.; Peddey, M.; Sillankorva, S.; Azeredo, J. The role of bacteriophages in periodontal health and disease. Future Microbiol. 2016, 11, 1359–1369. [Google Scholar] [CrossRef]
- Egido, J.E.; Costa, A.R.; Aparicio-Maldonado, C.; Haas, P.-J.; Brouns, S.J. Mechanisms and clinical importance of bacteriophage resistance. FEMS Microbiol. Rev. 2022, 46, fuab048. [Google Scholar] [CrossRef]
- Yeh, T.-K.; Jean, S.-S.; Lee, Y.-L.; Lu, M.-C.; Ko, W.-C.; Lin, H.-J.; Liu, P.-Y.; Hsueh, P.-R. Bacteriophages and phage-delivered CRISPR-Cas system as antibacterial therapy. Int. J. Antimicrob. Agents 2022, 59, 106475. [Google Scholar] [CrossRef] [PubMed]
- Soto-Perez, P.; Bisanz, J.E.; Berry, J.D.; Lam, K.N.; Bondy-Denomy, J.; Turnbaugh, P.J. CRISPR-Cas system of a prevalent human gut bacterium reveals hyper-targeting against phages in a human virome catalog. Cell Host Microbe 2019, 26, 325–335.e325. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.M.; Grossman, A.D. Integrative and conjugative elements (ICEs): What they do and how they work. Annu. Rev. Genet. 2015, 49, 577–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alseth, E.O.; Pursey, E.; Luján, A.M.; McLeod, I.; Rollie, C.; Westra, E.R. Bacterial biodiversity drives the evolution of CRISPR-based phage resistance. Nature 2019, 574, 549–552. [Google Scholar] [CrossRef]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef]
- Jansen, R.; van Embden, J.D.A.; Gaastra, W.; Schouls, L.M. Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microbiol. 2002, 43, 1565–1575. [Google Scholar] [CrossRef]
- Santiago-Rodriguez, T.M.; Naidu, M.; Abeles, S.R.; Boehm, T.K.; Ly, M.; Pride, D.T. Transcriptome analysis of bacteriophage communities in periodontal health and disease. BMC Genom. 2015, 16, 549. [Google Scholar] [CrossRef] [Green Version]
- Jore, M.M.; Brouns, S.J.; van der Oost, J. RNA in defense: CRISPRs protect prokaryotes against mobile genetic elements. Cold Spring Harb. Perspect. Biol. 2012, 4, a003657. [Google Scholar] [CrossRef]
- Coordinators, N.R. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2012, 41, D8–D20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gussow, A.B.; Park, A.E.; Borges, A.L.; Shmakov, S.A.; Makarova, K.S.; Wolf, Y.I.; Bondy-Denomy, J.; Koonin, E.V. Machine-learning approach expands the repertoire of anti-CRISPR protein families. Nat. Commun. 2020, 11, 3784. [Google Scholar] [CrossRef] [PubMed]
- Konstantakos, V.; Nentidis, A.; Krithara, A.; Paliouras, G. CRISPR–Cas9 gRNA efficiency prediction: An overview of predictive tools and the role of deep learning. Nucleic Acids Res. 2022, 50, 3616–3637. [Google Scholar] [CrossRef]
- Mohanty, R.; Asopa, S.J.; Joseph, M.D.; Singh, B.; Rajguru, J.P.; Saidath, K.; Sharma, U. Red complex: Polymicrobial conglomerate in oral flora: A review. J. Fam. Med. Prim. Care 2019, 8, 3480. [Google Scholar]
- Butera, A.; Gallo, S.; Pascadopoli, M.; Taccardi, D.; Scribante, A. Home oral care of periodontal patients using antimicrobial gel with postbiotics, lactoferrin, and aloe barbadensis leaf juice powder vs. conventional chlorhexidine gel: A split-mouth randomized clinical trial. Antibiotics 2022, 11, 118. [Google Scholar] [CrossRef] [PubMed]
- Gallo, S.; Scribante, A. Ozone therapy in dentistry: From traditional applications towards innovative ones. A review of the literature. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Sanya, China, 28–30 October 2020; p. 012001. [Google Scholar]
- Kim, J.; Amar, S. Periodontal disease and systemic conditions: A bidirectional relationship. Odontology 2006, 94, 10–21. [Google Scholar] [CrossRef] [Green Version]
- Martínez-García, M.; Hernández-Lemus, E. Periodontal inflammation and systemic diseases: An overview. Front. Physiol. 2021, 12, 709438. [Google Scholar]
- Hajishengallis, G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar]
- Radaic, A.; Kapila, Y.L. The oralome and its dysbiosis: New insights into oral microbiome-host interactions. Comput. Struct. Biotechnol. J. 2021, 19, 1335–1360. [Google Scholar]
- Chen, T.; Olsen, I. Porphyromonas gingivalis and its CRISPR-Cas system. J. Oral Microbiol. 2019, 11, 1638196. [Google Scholar] [CrossRef] [Green Version]
- Richter, C.; Chang, J.T.; Fineran, P.C. Function and regulation of clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR associated (Cas) systems. Viruses 2012, 4, 2291–2311. [Google Scholar]
- Hille, F.; Charpentier, E. CRISPR-Cas: Biology, mechanisms and relevance. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150496. [Google Scholar] [CrossRef] [Green Version]
- Sorek, R.; Kunin, V.; Hugenholtz, P. CRISPR—A widespread system that provides acquired resistance against phages in bacteria and archaea. Nat. Rev. Microbiol. 2008, 6, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Ly, M.; Abeles, S.R.; Boehm, T.K.; Robles-Sikisaka, R.; Naidu, M.; Santiago-Rodriguez, T.; Pride, D.T. Altered oral viral ecology in association with periodontal disease. MBio 2014, 5, e01133-14. [Google Scholar] [CrossRef] [PubMed]
- Szafrański, S.P.; Slots, J.; Stiesch, M. The human oral phageome. Periodontol. 2000 2021, 86, 79–96. [Google Scholar] [PubMed]
- Shanmugasundaram, S.; Nayak, N.; Puzhankara, L.; Kedlaya, M.N.; Rajagopal, A.; Karmakar, S. Bacteriophages: The dawn of a new era in periodontal microbiology? Crit. Rev. Microbiol. 2023, 1–12. [Google Scholar] [CrossRef]
- Landsberger, M.; Gandon, S.; Meaden, S.; Rollie, C.; Chevallereau, A.; Chabas, H.; Buckling, A.; Westra, E.R.; van Houte, S. Anti-CRISPR phages cooperate to overcome CRISPR-Cas immunity. Cell 2018, 174, 908–916.e912. [Google Scholar]
- Marraffini, L.A. CRISPR-Cas immunity against phages: Its effects on the evolution and survival of bacterial pathogens. PLoS Pathog. 2013, 9, e1003765. [Google Scholar] [CrossRef] [Green Version]
- Mitrofanov, A.; Alkhnbashi, O.S.; Shmakov, S.A.; Makarova, K.S.; Koonin, E.V.; Backofen, R. CRISPRidentify: Identification of CRISPR arrays using machine learning approach. Nucleic Acids Res. 2021, 49, e20. [Google Scholar]
- Steier, L.; De Oliveira, S.D.; De Figueiredo, J.A.P. Bacteriophages in dentistry—State of the art and perspectives. Dent. J. 2019, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Abedon, S.T.; Danis-Wlodarczyk, K.M.; Wozniak, D.J. Phage cocktail development for bacteriophage therapy: Toward improving spectrum of activity breadth and depth. Pharmaceuticals 2021, 14, 1019. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Shen, W.; Zhong, Q.; Chen, Q.; He, X.; Baker, J.L.; Xiong, K.; Jin, X.; Wang, J.; Hu, F. Development of a bacteriophage cocktail to constrain the emergence of phage-resistant Pseudomonas aeruginosa. Front. Microbiol. 2020, 11, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spacer_Begin_Position | Spacer_Length | Spacer_Sequence |
|---|---|---|
| 2181514 | 36 | AAACGAAATGAAAAAGACAACAAACAGAAGACCCTC |
| 2181580 | 36 | GTGCCAGCTGCAGGGGGATGACATAGCCATTGACGA |
| 2181646 | 36 | GTGCCAGCTGCAGGGGGATGACATAGCCATTGACGA |
| 2181712 | 36 | TCCGCGCCGCGAGGTGGAGACCCTGCCGGAGGCGAA |
| 2181778 | 37 | CCTGAGAAAGAGGGGAGGGAGGAGCGATAGACGAAGT |
| 2181845 | 36 | CGAGAATCTATTGAGTAGCGAAGTCGTCACAAAGAT |
| 2181911 | 36 | AGCCATAGCTCTTCAATTTCAATTTCTTCTTTTAAT |
| 2181977 | 36 | CGAAAATAACAAAAATAGATATATTTATAAAAAAGA |
| 2182043 | 34 | ACTCTTATCATCTACTATCTCAAAAGCTCTTTTT |
| 2182107 | 38 | TCGCTATAACCCTATGTGATTCAGGAATCGGCTTGCTA |
| 2182175 | 36 | GAAACATTCGAGCCGTATTCAATTACGCCATCAATG |
| 2182241 | 38 | CACCCATTGTGCCGCCGTCCTGACCGAAAACTTCTTTA |
| 2182309 | 38 | TCGCTATAACCCTATGTGATTCAGGAATCGGCTTGCTA |
| 2182377 | 36 | GAAACATTCGAGCCGTATTCAATTACGCCATCAATG |
| 2182443 | 38 | CACCCATTGTGCCGCCGTCCTGACCGAAAACTTCTTTA |
| 2182511 | 38 | TCGCTATAACCCTATGTGATTCAGGAATCGGCTTGCTA |
| 2182579 | 36 | GAAACATTCGAGCCGTATTCAATTACGCCATCAATG |
| 2182645 | 38 | CACCCATTGTGCCGCCGTCCTGACCGAAAACTTCTTTA |
| 2182713 | 36 | ATTTCTTCATCTCGCGCTTGCTCAAAAGCGCGTTCA |
| 2182779 | 34 | TGCATCAAGTCACGAACTTTCTGCGAGATGGAAA |
| 2182843 | 36 | CCGTCTCGAAAAAATATCGGGACGTTTTTGTTTTCT |
| 2182909 | 35 | CTCCTTCACTTTGTCGACAATGTGCACTGTATTTG |
| 2182974 | 36 | AGGCGGAGTATCTCTTTGCCACCCAGTCCGCGCGCA |
| 2183040 | 36 | CGTAACCGCCTCGGTAGACCGCTCCGCACGGTCGTT |
| 2183106 | 37 | GTGCATTCCGGACAGCTTTCGCTTAAAAAGTTAGCGG |
| 2183173 | 37 | AAGAACGAACGCCTGCGCGATAAGCACCGCGAGCGTA |
| 2183240 | 36 | ATTTACCTGCAGACTTGTGCCCACCCACTTGATAGA |
| 2183306 | 36 | TGGTCACGGAGCGATACCATGAGTGTTTAGTAGATG |
| 2183372 | 36 | CGGAGGAGATCAGCTATGCGGATGATACCACCCGTG |
| 2183438 | 36 | TCAGAAAACTCGCGTCCATCTGATAGATGTACACGA |
| 2183504 | 36 | TAAACGATCGAGGCGCGGAGACCCTCCTTGCCAGTA |
| 2183570 | 37 | CCGTTCAGGAAAAAGTAACCGAGCTGAAGACCATGCT |
| 2183637 | 35 | AGGTGCTATCGCAGGACTGCAGGACATCCTCAATC |
| 2183702 | 36 | AGCCGCTCGACTTGACGCCATGCAAAAACAGATAGA |
| 2183768 | 36 | TCTGCGAGTTGTGAGAGGCAATAAACTGCTGGGAGC |
| 2183834 | 35 | ATTGCTGTTAATTCTGTCATCTCTTATTTCTTTTA |
| 2183899 | 35 | TCTATCTGCGTATAGCCCTCGCCGTCCACGCCCTG |
| 2183964 | 37 | GGGAAGCTGCTTTGCTCGCTGAGATAGCAGCACTTCA |
| 2184031 | 36 | TTTATTGACGCCACCCCGCCGACGAAAAAAAATCAT |
| 2184097 | 36 | TTTCGGTCTTTACGTTTGTCGCCACGGATACATGCT |
| 2184163 | 37 | ATTGTAAATAAATTACATGGCTATTGAAAAACAAATT |
| 2184230 | 36 | GATTGTACGACTTTGCTATAAAGTCTGAGTTATTAA |
| 2184296 | 36 | TATCAACAATCACCGTCATATGTGTAATATACTTGA |
| 2184362 | 36 | GGGCAGGCGTATTGCCCCACTTCTCCCCGAATGCAT |
| 2184428 | 36 | TGAGGAATCATATCAGTGTTTATTTTTTCATCGATT |
| 2184494 | 36 | TGGAAGTGGGTAGAAGAAAGCCCCAACGTGTTTAAG |
| 2184560 | 36 | CATGACAAAGAGACGGTTATCGGTAGAGGACAGGCA |
| 2184626 | 35 | CGCGTGGAAGGGGCATGTACACTTGTAGTTCGCCC |
| 2184691 | 34 | TGACAGGCCCTGCAGCGTGTGAGAGCGGAAATGC |
| 2184755 | 37 | ATTTTCAATCATGATATTTTATTTTTCCTGCAAACGC |
| 2184822 | 36 | TCGCATGTGGGAGCGCGGCGGTCTCTGCTTTACGAA |
| 2184888 | 36 | TCAGCGTGATGAGCGCTTGAGGCTCCTCCATCGAGT |
| 2184954 | 36 | AAGGATGATTTGGAAAATTTAGTAAGATAGTTGATA |
| 2185020 | 37 | TTCTTGGAGAAAGCGAAGACCATGAACCTAAGCGTCG |
| 2185087 | 36 | GGAAATATAGTTATTGTATCTACTAAAAGACATAGT |
| 2185153 | 34 | ACCCCATCTTGCAGAGTATATGCGAGCAAAATTT |
| 2185217 | 34 | TAGTTAGCACAGTTGCTACTATCGTAGTAGCTGT |
| 2185281 | 35 | AGATAAACTTTCTTCTCGAATTAAGAAAATCGAGA |
| 2185346 | 36 | CCGCGGCCATCGAGGCCACCGCCTCCGTCCTCCGCG |
| 2185412 | 37 | TATGAAAACAGAAAAGAACTTCTCAGCCCTGAGTTTG |
| 2185479 | 37 | CAAACTCAATGATTATCTGTCAAGAAGCAAGAAACGA |
| 2185546 | 37 | AGTAAAAATTACCCTAGATGCCGAAACGGACGGCCTT |
| 2185613 | 36 | CAGAATTTGCACGAACAGTATGATGTTCGTGTTCTT |
| 2185679 | 36 | AAGCGCGAGACAGGCCGAGCCGGCACAGCTTAGTGC |
| 2185745 | 36 | AAAACGGCGATAAAATAGCGTTCGAGATTTTCCGCA |
| 2185811 | 37 | TAGTTGTAGCGATTGTCTCAGTTGCATTACTCCTTAC |
| 2185878 | 37 | AAATAACGAGAAAAAGAATGCTTAAATTGTTCTTCGT |
| 2185945 | 36 | AGAAGAGGGTAAACTATTTGCTAATCTTGAATGCTT |
| 2186011 | 36 | TCTCAATATCTTTCATAGCTACTAAAAATTTACGAA |
| 2186077 | 35 | ATTGCTGTTAATTCTGTCATCTCTTATTTCTTTTA |
| 2186142 | 35 | TCTATCTGCGTATAGCCCTCGCCGTCCACGCCCTG |
| 2186207 | 36 | TGCCCCCTCACCCATCTAACCTCGAGCCGTTGAGCC |
| 2186273 | 36 | GATTACATAATGATAGACGACAGAGATTGTGCAGAA |
| 2186339 | 35 | GTACTGATAATTACGCTGCAAGGTCAGACGGTGAT |
| 2186404 | 36 | TTGCCAGGGCTTGCTGATGCGCGCGCTCCAGCTGCT |
| 2186470 | 36 | ACAGAACCAGCTCCGTCAAATCTCCCGCTTTTTGTC |
| 2186536 | 36 | TGCCCCCTCACCCATCTAATCTCGAGCCGCTGAGCC |
| 2186602 | 36 | TATTCATTCGCTCAAGCGAGGGCATCCTGCTGCAGC |
| 2186668 | 36 | GCTCGCGGAGGGAGAGGGTGCCGGTATCCTGCCTCC |
| 2186734 | 36 | AGAGTCTGTATGTGAAAGTGTAGTTCGAAACATTAT |
| 2186800 | 34 | AGAATACTTTGAAGTTGTATTCAATTACTTTGAA |
| 2186864 | 36 | CCGTTGTGACAGAGCTGCGCCGTCGTGGCTATGATG |
| 2186930 | 36 | TGAAAGCCCGAAGATAATCTACACGCAAGATTGTTA |
| 2186996 | 35 | CGTCATCAGGTGGATATTCTTACTGCTATCCACGA |
| 2187061 | 36 | TCGTCTGCGACGTATCGCAGCTCTGCCAGCTCCGCG |
| 2187127 | 36 | GCATCTTCGCAGGAAAGAAGAAGGCTCCGTCCTCGA |
| 2187193 | 36 | AACAGTCAAGGGGGAGCCGACCTCTCCGGCGGATTT |
| 2187259 | 36 | GCAGTGCGGCCGACAAGGCTAACCTCGCTCAGCTGA |
| 2187325 | 36 | AATCCATGTTCATGAGGAGAGGATACGAGTTCTATT |
| 2187391 | 35 | CCTTTTGCTATTGCAGCAAACATATACAATAATAA |
| 2187456 | 36 | ACTTCATGGATTTAGCGAGATACTCATTATAATTAA |
| 2187522 | 36 | TCTTCAAAGATAGTTGTTATCAAATATCGCGCTGAA |
| 2187588 | 37 | ATCTTCAAAGATAGTTGTTATCAAATATCGCGCTGAA |
| 2187655 | 36 | GCTCAAGCGTCCAAGAAAGTTCAAGAAATTGTACTA |
| 2187721 | 37 | TACTATGGTGTCTGCATTTGAGATACGCAAATAGCAA |
| 2187788 | 36 | CCTCAATATCTTTCATTGCGACGAGAAATTTGCGAA |
| 2187854 | 36 | AATCTATGATGATATAGAAGAAGACGACTTGTTGTT |
| 2187920 | 36 | ATTAATGATTTTCTGAAATAAAGAAGCAGTTGCATA |
| 2187986 | 35 | CTCAATATCTTTCATTGCGACGAGGAACTTGCGAA |
| 2188051 | 36 | CCCCGTGGTAACTCATACCACCGACTATTCCACCGT |
| 2188117 | 36 | TTTGAGTAATCATCGAATAATTATCGATTAATCATT |
| 2188183 | 36 | TTATATGCATCATATTCTTAAAGTATTTTATTTGAA |
| 2188249 | 36 | GACGAATACGGCGTTCATCATCGATAGTCGCGATGC |
| 2188315 | 36 | CGGGCGGCCAACCGGTCACAACAAGAATAGACCGAT |
| 2188381 | 35 | TTCCTTCCAGTCGCAGCTTAAGATACTGCGACTAC |
| 2188446 | 36 | AATTTTCATCAGAGCATAAAAAAGGGCAAACTTTTT |
| 2188512 | 35 | GATACGAGCACCAAGGCTGCGATACCGATTGCGTA |
| 2188577 | 36 | CCTCGAGCAGATCTTCCTGCTCTTTGATGAGTGAGG |
| 2188643 | 35 | GCAGGTTGATTATAATGTTGAAGATGCTTTAAGGG |
| 2188708 | 36 | CTCGTATCGACTTTCAAGCAGGCTGGAGTGCAGCCT |
| 2188774 | 37 | AATATGAGATCGGAAACAATTATAGTTGCGTCGATAT |
| 2188841 | 37 | TCACAATAACCGACAAAATGTCTCGCGTAACGTACAA |
| 2188908 | 35 | GTCGCCTGTTTTCTTGAACTCCTCACTGATTCGTA |
| 2188973 | 35 | GATGGTGTCGACATCATACGACAATAGATCGTCGA |
| 2189038 | 35 | GATGGTGTCGACATCATACGACAATAGATCGTCGA |
| 2189103 | 36 | AATATTTTTTCAAAAATTGTAAAACTTATTAAGTCA |
| 2189169 | 36 | AAGTTGCTGATTGTCTTAGAATGAAAGGTTATGCTC |
| 2189235 | 35 | ATAATATTACATAATGGCACACTGATGGTAAACTT |
| 2189300 | 34 | CATGATGTACAAATATATCATGATCGTATACTAC |
| Spacer_Begin_Position | Spacer_Length | Spacer_Sequence |
|---|---|---|
| 367189 | 30 | TATAGGAGGTTTCAAAATGGAAAAATCGAA |
| 367255 | 30 | TATCAAGTTGAGCCTTCTTTAAAGCTCCGC |
| 367321 | 30 | TATAGGAGTTCCAGACCCAGCACCATCACC |
| 367387 | 30 | AAAATCGAATGTATCGCAAGATTCAAACCA |
| 367453 | 30 | TACAAAATCGAAGCAGAAGAAAGGAACTTC |
| 367519 | 30 | GGTTCCAATCTTTTTGGAATGATTAACAAT |
| 367585 | 30 | GATTCTGTATTTCAACGCGATGTTGCTAAT |
| 367651 | 30 | CTAACAAAAGGTGGAATTTTACCGAACAAT |
| 367717 | 29 | AATTAGTTGTCATTGAAGGTGAAGCCGGA |
| 367782 | 30 | GCGGAAAAACTATATCGTAATCTTCATAGA |
| 367848 | 30 | GCTGGAACGCCTATAGCGACGCAAGCTCCT |
| 367914 | 30 | CGCTGGAACGCCTATAGCGACGCAAGCTCC |
| 367980 | 30 | GGTTCCAATCTTTTTGGAATGATTAACAAT |
| 368046 | 30 | CATCTAGAATCCTATAAGGCACGAAGTAAT |
| 368112 | 30 | CCTTTTTTGTAACTCCTATTTGCAGCTATG |
| 368178 | 30 | ATTACTTTTCGAAAAAAAGCCGTATTATAG |
| 368244 | 30 | TCTTTGTATTATAAAGTTAGCAGAGGAAAA |
| 368310 | 30 | GAATCTACCACCCTCAATACTCCGCCTATT |
| 368376 | 30 | GTCAACATCACCGCGATCACTACAAACAGC |
| 368442 | 30 | GAATGAAAAGGACAAGGAAAAAGCTGCCCT |
| 368508 | 30 | TGATTATTTGGAAGGCATGAGTAAATGCTG |
| 368574 | 30 | GCAGTAACTCACAAGCCACTTTGAGAGTTG |
| 368640 | 30 | TTCGACGCTTGTCGAAAAGGCAATCAAGGC |
| 368706 | 30 | CGAGAAGTTATTATTCTGAACTTCACATCG |
| 368772 | 30 | CTTTGGTATCAATTAGGATTTCCTAAAGTC |
| 368838 | 30 | TACAATGATTGCTTGTTGTTCTGATGGAAC |
| 368904 | 30 | TAGCCTCACCATTATAAAGCAATTCGCATG |
| 368970 | 30 | TGTTACGTCAAAAAATCCAATAAGTTGAAG |
| 369036 | 30 | CCTGATAAGGAAGATTGGCGAAAGAAGGTA |
| 369102 | 30 | TGCTACATCAAATAACCCTACAAGTTGAAG |
| 369168 | 30 | CCAAAAGTTCACAGTCATCCGAGTAGACGT |
| 369234 | 30 | CTATCTACTTTTGGGAACCCTAATTGGTAC |
| 369300 | 30 | TTTCTTCTGTTTGTCCATGTCCAAACCTCC |
| 369366 | 30 | AACAATGTGTGATTTTTCGGACTTAGTCCC |
| 369432 | 30 | AAGGGAATAACCTTACCATTCTGTCTTATG |
| 369498 | 30 | TTCCCAAAAGTTGATGCTGATACGATTGGT |
| 369564 | 30 | AACAATCAGCCGTGAGGGAATACGCCGCGT |
| 369630 | 30 | AGGTTAATGATGAAAAAAATAATAACTACT |
| 369696 | 30 | GGGCATATTATGCAGATATGCAACGAAACG |
| 369762 | 30 | CTTGGAAAAGAATTTATAAAATGCGAAGTT |
| 369828 | 30 | GAACATATGCTCGCTCTTTCTCGAGTACTC |
| 369894 | 30 | AAACTTTGAGGTACTAAATAAAACAAGTCA |
| 369960 | 30 | ACCTTTCAATAGTAGCATCGGGCAAACCAG |
| 370026 | 30 | GTCTCTAGTTACTTTACGTATAAACTCTAT |
| 370092 | 30 | GGGCATATTATGCAGATATGCAACGAAACG |
| 370158 | 30 | CTTGGAAAAGAATTTATAAAATGCGAAGTT |
| 370224 | 30 | ATGCGATATATCTATGACTTTACCTATTCT |
| 370290 | 30 | AAACTTTGAGGTACTAAATAAAACAAGTCA |
| 370356 | 30 | ATATCTTTTGTCGTTAAAGTTAGTAAAAAA |
| 370422 | 30 | TTTGAAATTCCCCAAATGTCAATTGTTTTC |
| 370488 | 30 | GAAAATGCAGGCGGTTCCACTGGAGAGGTT |
| 370554 | 30 | TAATTCAAAAAAAGGTCTTGGTTTGAAAGG |
| 370620 | 30 | AGCCCGCCCTGCGGAATTGCACGGCCCGTT |
| 370686 | 30 | ATTGAGCGTCAAGCACCCGGTAAGCCCACC |
| 370752 | 30 | TTGGTTATTCGACTTTTGATTTGAGCTATC |
| 370818 | 30 | CTCGCTCGAGCACAACAGGTGGCTGTCCAC |
| 370884 | 30 | TTTCCAGCTAGAGCATCAAAGTTTATAGGG |
| SPACER ID | POSITION | SEQUENCE |
|---|---|---|
| 2508368 | 36 | ACAGAAACTTCTTTTCCTGCAAGATTAAATAATACA |
| 2508434 | 37 | GGTATGTATAAATCTACACGTCTTGGGTTTTCTAATA |
| 2508501 | 40 | AAAATTATCTTTGATAACTTTAAGAATCTTTTTGTCTTCT |
| 2508571 | 40 | CCATTACTGCGCGGGCGGCGATGCAGGAGAACCCGGACCG |
| 2508641 | 39 | TAAAAGGTTAAAAGTTAAATGAAGAAAGACATAAAACGA |
| 2508710 | 36 | GTCTTTGGAGGCCTTTACTCTTTTAAAAATGCCCGA |
| 2508776 | 34 | TCTCTTGTGTAGTTATAACACACAATTGAGTCAT |
| 2508840 | 35 | CCTCTTCGTAATACGGCTCTATATCGAGCTCTCTG |
| 2508905 | 38 | CCCCGAAGGCGCGGCCGTTCCACTCTAAGGTGTCGACT |
| 2508973 | 36 | GTTAGATGATAACTTCCGTCTTCTCCGAGCATCATC |
| 2509039 | 36 | TTCACGGGGGTAAAGCCCGCCCCTTACGGGGAACTA |
| 2509105 | 37 | AAAAGATTTGCTATATTGTGAAAAATTTAAAAAAAAG |
| 2509172 | 39 | TTCACGGGGGTAAAGCCCGCCCCTTACGGGGAACTACTC |
| 2509241 | 38 | TGCAGCCGTTGTCTCGCAAAAAGCAGCGCCTTCTAAAA |
| 2509309 | 36 | CCTGCCCTGCGTGGGGGCTTTGGTGTGCGCAGCCTG |
| 2509375 | 37 | CTTCAAAAGCGGCAGCGCGCTTTTTGAGAAGGCGCTG |
| 2509442 | 37 | TTTGTGTTATACGGAGATTATACACGGTGGGTGTGGG |
| 2509509 | 37 | TGCGCGCATACGTTCTTCTACGTACGCCTTTTTGGTT |
| 2509576 | 38 | AAAAGATTTGCTATTTTGTAATAGCAAAGAACTATGAA |
| 2509644 | 35 | TAAGAGGTTATCTCCGTCCGCACGAGTTCGGACGT |
| 2509709 | 36 | CATACGTTGCACGAATGTCCGCATCAAAAGCGGCAG |
| 2509775 | 36 | GTTATTCTCCCAATCACCGAACCTGTCAACAAAAGA |
| 2509841 | 38 | CTATAACTGCAGCCCGCACGGCTGCACGGTTGGCGTAC |
| 2509909 | 37 | GTCTGGTCGTTTTTTCTGTATTGGGGGGGGCGGCATT |
| 2509976 | 36 | ACGTTAACACGTGCACCGCCAACGCGGGCAGAAAAG |
| 2510042 | 35 | CGTATCTGAAACGTCCGAATTGGTGCGGACGGAGG |
| 2510107 | 36 | GCCACCACCGACCCTGCACGGGCGGTTTGGCTCGCG |
| 2510173 | 37 | ACATAGCAAACCTTGTTGGATAATATTCTTCGTCTAT |
| 2510240 | 36 | GAACAATAACAAACAGGATTGGGCGATCATACTTTT |
| 2510306 | 36 | CTCGGATTTATCCATTACGTCGGCCGCATCGATTTG |
| 2510372 | 38 | CCAACGCGGGGAAGAAGGCGGATGAGGTTTCGGAACAA |
| 2510440 | 37 | CGCCGAGAAGGTCTCCGTCCTCGTCAAAAACAGACTC |
| 2510507 | 37 | GCTCGGGGATCGATATAATAGAATGGTAGAGGAGTGG |
| 2510574 | 38 | TACGGGGTCACCTCCGTTCGCATGAGTTTGGACGTTTC |
| 2510642 | 36 | CATATAACAGTGCGACGAGACAGGCTGCGCACACCA |
| 2510708 | 37 | GGGGTAAATCATGTAAAACGAACAATTTTAGAATATA |
| 2510775 | 36 | CGTAGCGCTATTATGCGCGGTATGGCGCGGCTCTGC |
| 2510841 | 36 | CTTGCAGTAGGGGCAGTGAAAATTACACGACCCGAA |
| 2510907 | 39 | CTATTACCGCCGCCCGCACGGGCGCACGGCTGGCGTACG |
| 2510976 | 38 | GAATAAAAATCAGCGTAAAAACATATATGCTTACACGA |
| 2511044 | 38 | TTTTTTGTGGCCGTTTCGGCCTTATCTGTATCTACTGT |
| 2511112 | 36 | AGCCGACGCGATATATTCGTGCCCGGCAAACAAATC |
| 2511178 | 36 | CGCCGTACACGTGGTGATATTTATGAAACAAATTTT |
| 2511244 | 36 | GATATATTTTTCTAAACATTTTCATGACTTTATCGG |
| 2511310 | 36 | TGATAACTTTTAAAGTCTTTTTATCCTCTGCATTCA |
| 2511376 | 37 | ATTGTACACCTTGTCTACTTGCTCATCAGGGAAATTC |
| 2511443 | 37 | GTAATCCTCACACTGCCGATTATGGCTGAGTAATCTA |
| 2511510 | 36 | AGTTGATGCAAAAGGTTTATCGAAACACCTTCCTTC |
| 2511576 | 37 | CCTTATCTCAATGGTTTAAGGAAGACGTTGCGCGCAT |
| 2511643 | 36 | AGCCTATCTAAACGCCAAATAACTAATTTGTCTCCT |
| 2511709 | 37 | TCGGTTTGGTGATAGTTGGTAACAAGATTTTAAAACA |
| 2511776 | 34 | ATTACATCAGGATAGATGGTGTGGCGTATGAAAA |
| 2511840 | 37 | GAAGCTGGTAACCGCGGATGTCGCCTTCGGCCTCGAA |
| 2511907 | 38 | CCGCTACCCAAAAAACCCTCGACAAAATGCCCGAAATC |
| 2511975 | 37 | CACTGGGGCACATAGTCCTGATCTCCTCGGCGAGTAT |
| 2512042 | 37 | GAAAAGTGCTAATGGTAAAGTTAAACTTTCTCCTTTA |
| 2512109 | 35 | TTAGATGATCTTGCGGACTACAGTGCCGAATTTGA |
| 2512174 | 36 | GCAAACGGGGCCTTTTGTGCCATGTCACCAGTGCGG |
| 2512240 | 37 | AGAGACGTTCCTCCTGCGTTGTACAAGATACTCTGTA |
| 2512307 | 34 | AATTTAAGTGATGATGTAAAAACTGGTATTAATA |
| 2512371 | 35 | TTAGATGATCTTGCGGACTACAGTGCCGAATTTGA |
| 2512436 | 36 | TTCGCAATCGCTTCGACGGCAAAAGTGCCGCCCAGG |
| 2512502 | 35 | TGTTTATGTACGACTCTTTCAGTTTTAACTGCTTT |
| 2512567 | 39 | GATATAATGCTTACAATCGAGGCATTTAATAACATTAAA |
| 2512636 | 39 | CAATGTGTACGTTGTCATTGCCGGTATATTAGGATTCTT |
| 2512705 | 35 | AAGTGCCTTTGCCGTTAATTTTGTCAATGAGTTTC |
| 2512770 | 38 | GCGAAATCCTGCTCGTGCAGGGTGGAAGCAAGAATATC |
| 2512838 | 38 | AGCGTCATAGCATTCACACCGGCAGCACCAGTTACGAA |
| 2512906 | 37 | GTCGGACACGATGGGCAGAAATTCTTCTTCGACCATG |
| 2512973 | 35 | CCGTTAAATTGTCTGGCAAGGACGTGACGCCGGTA |
| 2513038 | 36 | CGTTGTAGAAATCGTCTTCGTTGATAACAAGTGTTA |
| 2513104 | 37 | GCTTCTTCGATTCTTCTTTTACCTCTCCGGTTTCCGT |
| Model | AUC | CA | F1 | Precision | Recall | LogLoss | Specificity |
|---|---|---|---|---|---|---|---|
| Random Forest | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 0.097 | 1.000 |
| Neural Network | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 0.021 | 1.000 |
| SVM | 0.938 | 1.000 | 1.000 | 1.000 | 1.000 | 0.387 | 1.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yadalam, P.K.; Arumuganainar, D.; Anegundi, R.V.; Shrivastava, D.; Alftaikhah, S.A.A.; Almutairi, H.A.; Alobaida, M.A.; Alkaberi, A.A.; Srivastava, K.C. CRISPR-Cas-Based Adaptive Immunity Mediates Phage Resistance in Periodontal Red Complex Pathogens. Microorganisms 2023, 11, 2060. https://doi.org/10.3390/microorganisms11082060
Yadalam PK, Arumuganainar D, Anegundi RV, Shrivastava D, Alftaikhah SAA, Almutairi HA, Alobaida MA, Alkaberi AA, Srivastava KC. CRISPR-Cas-Based Adaptive Immunity Mediates Phage Resistance in Periodontal Red Complex Pathogens. Microorganisms. 2023; 11(8):2060. https://doi.org/10.3390/microorganisms11082060
Chicago/Turabian StyleYadalam, Pradeep Kumar, Deepavalli Arumuganainar, Raghavendra Vamsi Anegundi, Deepti Shrivastava, Sultan Abdulkareem Ali Alftaikhah, Haifa Ali Almutairi, Muhanad Ali Alobaida, Abdullah Ahmed Alkaberi, and Kumar Chandan Srivastava. 2023. "CRISPR-Cas-Based Adaptive Immunity Mediates Phage Resistance in Periodontal Red Complex Pathogens" Microorganisms 11, no. 8: 2060. https://doi.org/10.3390/microorganisms11082060