

The Intestinal Microbiome after Traumatic Injury
 , , ,
, , ,  and
and
Abstract
:1. Microbiome
1.1. Maintenance of the Intestinal Barrier
1.2. Role in the Immune System and Inflammation
1.3. Microbiome Characterization
2. Trauma
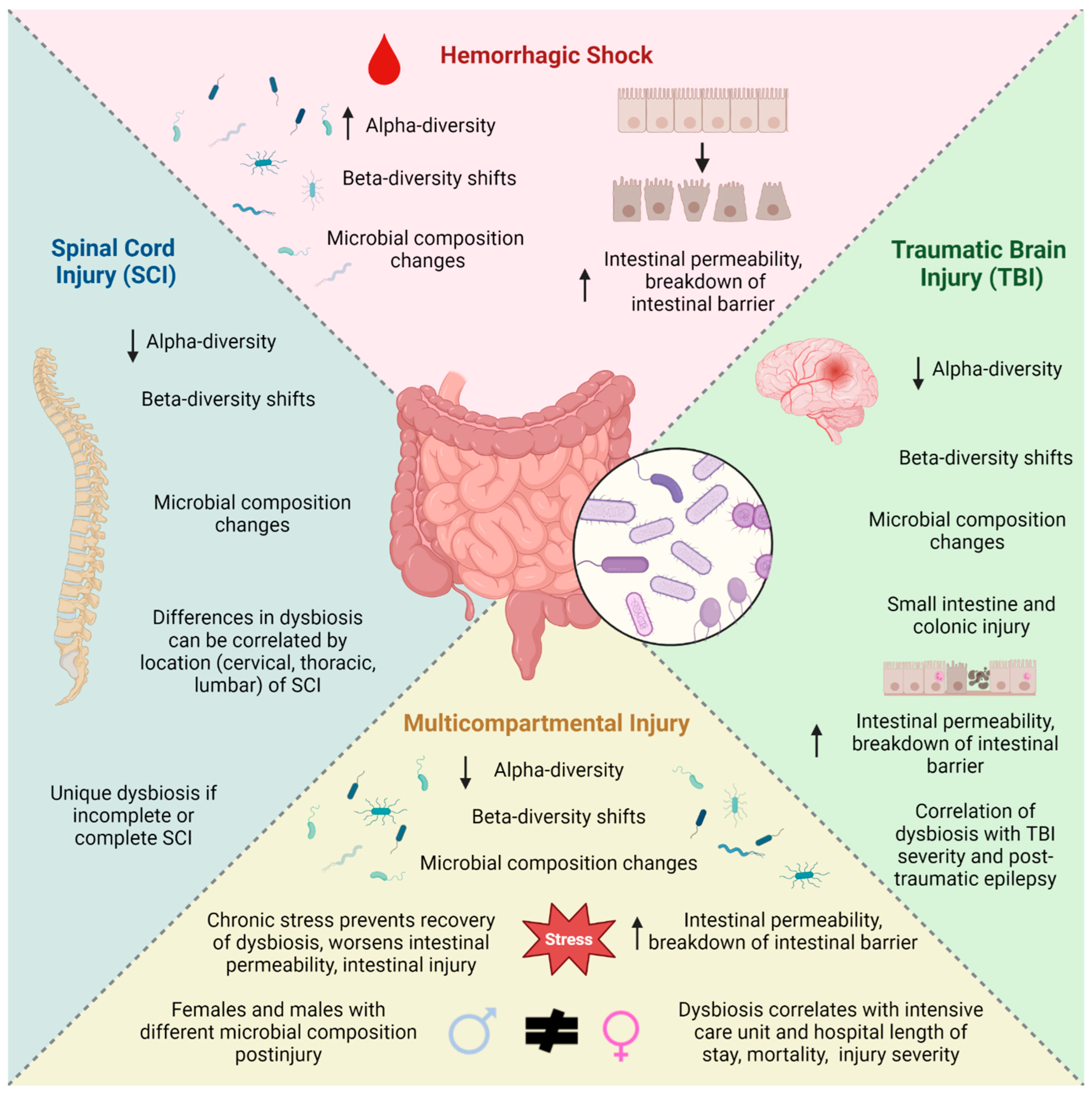
2.1. Hemorrhagic Shock
2.2. Traumatic Brain Injury

{kind=link}
| Author and Year | Species | Trauma Model | Time Point(s) Studied Postinjury | Fecal Microbiome Diversity and Composition Changes | Intestinal Barrier Effects | |
|---|---|---|---|---|---|---|
| Age matters: microbiome depletion prior to repeat mild traumatic brain injury differentially alters microbial composition and function in adolescent and adult rats | Sgro et al., 2022 [33] | Male and Female Sprague-Dawley Rats | TBI§ (repetitive) | 17 and 30 days | ↔ Alpha-diversity (30 d) ↔ Beta-diversity (30 d) ↔ Microbial composition (30 d) | ↔ Tight-junction protein expression in small intestine (30 d) ↓ Occludin expression in small intestine (30 d) |
| Traumatic brain injury induces gastrointestinal dysfunction and dysbiosis of gut microbiota accompanied by alterations in bile acid profile | You et al., 2022 [19] | Male C57BL/6J Mice | TBI† | 1 h, 6 h, 1, 3, and 7 days | ↓ Alpha-diversity (all time points) Beta-diversity differences (1 d, 3 d, 7 d) ↓ Firmicutes (6 h) ↑ Bacteroidetes, Proteobacteria (6 h) Changes in genus composition over time | ↔ ZO-1 expression in colon and small intestine (1 d, 3 d) ↓ Occludin expression in colon (3 d) ↓ Claudin-1 expression in small intestine (1 d, 3 d), colon (1 d) ↓ Claudin-2 expression in small intestine (3 d), colon (1 d) |
| Translocation and dissemination of gut bacteria after severe traumatic brain injury | Yang et al., 2022 [20] | Male C57BL/6 Mice | TBI† | 3 h, 1, 3, 7, and 14 days | ↑ Alpha-diversity (3 d, 7 d) Beta-diversity differences ↓ Firmicutes, Lactobacillus, Akkermansia, Roseburia (7 d) ↑ Bacteroidetes, Streptococcus, Lachnoclostridium (7 d) | ↑ Villous atrophy in small intestine (1 d, 3 d, 7 d, 14 d) ↓ Lysozyme from Paneth cells in small intestine (3 d) ↑ Caspase-3 in small intestine (1 d, 3 d, 7 d, 14 d) |
| Oral administration of brain protein combined with probiotics induces immune tolerance through the tryptophan pathway | Hou et al., 2021 [21] | Male Sprague-Dawley Rats | TBI† | 3 and 7 days | ↓ Alpha-diversity (3 d) Beta-diversity differences ↑ Akkermansia (3 d) ↓ Lactobacillus (3 d, 7 d), Parabacteroides (7 d) | ↓ Claudin in colon, small intestine (3 d, 7 d) ↓ Occludin in colon, small intestine (3 d, 7 d) ↑ Plasma fluorescein isothiocyanate-dextran (3 d, 7 d) |
| Sustained dysbiosis and decreased fecal short-chain fatty acids after traumatic brain injury and impact on neurologic outcome | Opevemi et al., 2021 [41] | Male C57BL6/J Mice | TBI† | 3 h, 1, 3, 7, 14, and 28 days | ↓ Alpha-diversity (7 d, 14 d, 28 d) Beta-diversity differences ↑ Verrucomicrobiaceae, Erysipelotrichaceae ↓ Lachnospiraceae, Ruminococcaceae, Bacteroidaceae | NS |
| Effects of traumatic brain injury on the gut microbiota composition and serum amino acid profile in rats | Taraskina et al., 2022 [24] | Male Wistar Rats | TBI† | 7 days | ↓ Alpha-diversity Beta-diversity differences ↓ Bacteroidetes ↑ Rikenellaceae, Prevotellaceae, Lactobacillus, Turicibacter, Helicobacter | NS |
| Traumatic brain injury in mice induces acute bacterial dysbiosis within the fecal microbiome | Treangen et al., 2018 [18] | Male C57BL/6J Mice | TBI† | 24 h | ↓ Lactobacillus ↑ Marvinbryantia, Clostridiales | NS |
| Moderate traumatic brain injury alters the gastrointestinal microbiome in a time-dependent manner | Nicholson et al., 2019 [23] | Male Sprague-Dawley Rats | TBI† | 2 h, 1, 3, and 7 days | ↓ Alpha-diversity (3 d) Beta-diversity differences (1 d, 3 d) ↓ Firmicutes (1 d, 3 d), Lachnospiraceae (2 h, 3 d), Mogibacteriaceaei (1 d, 3 d) ↑ Bacteroidaceae (1 d), Verrucomicrobia (1 d), Enterobacteriaceae (3 d) | NS |
| Differential fecal microbiome dysbiosis after equivalent traumatic brain injury in aged versus young adult mice | Davis 4th et al., 2021 [32] | Male C57BL/6 Mice | TBI† | 1, 7, and 28 days | ↔ Beta-diversity over time ↑ Clostridium clocleatum, Anaerostipes, Lactobacillus, Coprococcus | NS |
| Susceptibility to epilepsy after traumatic brain injury is associated with preexistent gut microbiome profile | Medel-Matus et al., 2022 [26] | Male and Female Sprague-Dawley Rats | TBI† | 7 days, 1 month, and 7 months | ↔ Alpha-diversity over time Beta-diversity differences over time ↓ Lachnospiraceae (7 d, 1 mo, 7 mo), Lactobaccillus (1 mo) ↑ Ruminiclostridium, Pseudomonas (7 d), Bacteroides (1 mo), Parabacteroides (7 mo) | NS |
| Traumatic brain injury alters the gut-derived serotonergic system and associated peripheral organs | Mercado et al., 2022 [38] | Male C57BL/6J Mice | TBI† | 1, 3, and 7 days | ↓ Clostridium leptum (7 d) ↑ Clostridium scindens (7 d) | NS |
| Acute gut microbiome changes after traumatic brain injury are associated with chronic deficits in decision making and impulsivity in male rats | Frankot et al., 2023 [25] | Male Long-Evans Rats | TBI† | 3, 30, and 60 days | ↓ Alpha-diversity (3 d v 30 d) Beta-diversity differences over time ↓ Firmicutes, Bacteroides | NS |
| An integrated analysis of gut microbiota and the brain transcriptome reveals host–gut microbiota interactions following traumatic brain injury | Bao et al., 2023 [22] | Male C57BL/6 Mice | TBI† | 7 days | ↔ Alpha-diversity ↔ Beta-diversity ↑ Actinobacteria, Bifidobacteriales, Bifidobacteriaceae, Bifidobacterium | NS |
| Bidirectional brain–gut interactions and chronic pathological changes after traumatic brain injury in mice | Ma et al., 2017 [34] | Male C57BL/6 Mice | TBI† | 24 h, 28 days | NS | ↑ Colonic injury (28 d) ↔ Jejunal injury (24 h, 28 d) ↓ Transepithelial electrical resistance in jejunum (24 h) ↑ Paracellular flux in colon (28 d) ↔ ZO-1 in colon (28 d) ↔ Occludin expression in colon (28 d) ↓ Claudin-1 in colon (28 d) ↔ Claudin-2 in colon (28 d) |
| CORM-3 exerts a neuroprotective effect in a rodent model of traumatic brain injury via the bidirectional gut–brain interactions | Zhang et al., 2021 [35] | Male Sprague-Dawley Rats | TBI† & HS | 24 h, 30 days | NS | Jejunal histological changes at 24 h postinjury characterized by cleaved caspase-1-positive cells, pyroptotic cells |
| Disruption of the intestinal barrier and endotoxemia after traumatic brain injury: implications for post-traumatic epilepsy | Mazarati et al., 2021 [36] | Male Sprague-Dawley Rats | TBI† | 1 week, 7 months | NS | ↑ Plasma lipopolysaccharide (1 week, 7 month) ↑ Plasma fluorescein isothiocyanate-labeled dextran (1 week, 7 month) |
2.3. Spinal Cord Injury
2.4. Multicompartmental Injury
3. Therapeutics
3.1. Probiotics, Prebiotics, and other Medications
3.2. Fecal Microbiota Transplantation
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Miniet, A.A.; Grunwell, J.R.; Coopersmith, C.M. The microbiome and the immune system in critical illness. Curr. Opin. Crit. Care 2021, 27, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.; Mendez-Garcia, C.; Rojo, D.; Barbas, C.; Moya, A. Antibiotic use and microbiome function. Biochem. Pharmacol. 2017, 134, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Le Roy, C.I. Effect of Diet on the Gut Microbiota: Rethinking Intervention Duration. Nutrients 2019, 11, 2862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martino, C.; Dilmore, A.H.; Burcham, Z.M.; Metcalf, J.L.; Jeste, D.; Knight, R. Microbiota succession throughout life from the cradle to the grave. Nat. Rev. Microbiol. 2022, 20, 707–720. [Google Scholar] [CrossRef]
- Alverdy, J.C.; Krezalek, M.A. Collapse of the Microbiome, Emergence of the Pathobiome, and the Immunopathology of Sepsis. Crit. Care Med. 2017, 45, 337–347. [Google Scholar] [CrossRef]
- Rao, J.N.; Wang, J.Y. Regulation of Gastrointestinal Mucosal Growth. In Integrated Systems Physiology: From Molecule to Function to Disease; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2010. [Google Scholar]
- Camilleri, M. Leaky gut: Mechanisms, measurement and clinical implications in humans. Gut 2019, 68, 1516–1526. [Google Scholar] [CrossRef]
- Lueschow, S.R.; McElroy, S.J. The Paneth Cell: The Curator and Defender of the Immature Small Intestine. Front. Immunol. 2020, 11, 587. [Google Scholar] [CrossRef] [Green Version]
- Pelaseyed, T.; Bergstrom, J.H.; Gustafsson, J.K.; Ermund, A.; Birchenough, G.M.; Schutte, A.; van der Post, S.; Svensson, F.; Rodriguez-Pineiro, A.M.; Nystrom, E.E.; et al. The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunol. Rev. 2014, 260, 8–20. [Google Scholar] [CrossRef] [Green Version]
- Negi, S.; Das, D.K.; Pahari, S.; Nadeem, S.; Agrewala, J.N. Potential Role of Gut Microbiota in Induction and Regulation of Innate Immune Memory. Front. Immunol. 2019, 10, 2441. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhu, L.; Qin, S. Gut Microbiota Modulation on Intestinal Mucosal Adaptive Immunity. J. Immunol. Res. 2019, 2019, 4735040. [Google Scholar] [CrossRef] [Green Version]
- Takiishi, T.; Fenero, C.I.M.; Camara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, A.D.; Reid, J.N.; Macklaim, J.M.; McMurrough, T.A.; Edgell, D.R.; Gloor, G.B. Unifying the analysis of high-throughput sequencing datasets: Characterizing RNA-seq, 16S rRNA gene sequencing and selective growth experiments by compositional data analysis. Microbiome 2014, 2, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Chen, J. A comprehensive evaluation of microbial differential abundance analysis methods: Current status and potential solutions. Microbiome 2022, 10, 130. [Google Scholar] [CrossRef] [PubMed]
- Laudadio, I.; Fulci, V.; Palone, F.; Stronati, L.; Cucchiara, S.; Carissimi, C. Quantitative Assessment of Shotgun Metagenomics and 16S rDNA Amplicon Sequencing in the Study of Human Gut Microbiome. OMICS 2018, 22, 248–254. [Google Scholar] [CrossRef]
- Liu, Y.X.; Qin, Y.; Chen, T.; Lu, M.; Qian, X.; Guo, X.; Bai, Y. A practical guide to amplicon and metagenomic analysis of microbiome data. Protein Cell 2021, 12, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J.; Zhao, C.; Gai, Z.; Mu, X.; Wang, Y.; Zhang, C.; Su, Z.; Gao, L.; Zhu, D.; et al. Blood Loss Leads to Increase in Relative Abundance of Opportunistic Pathogens in the Gut Microbiome of Rabbits. Curr. Microbiol. 2020, 77, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.; Kuang, L.; Xiang, X.; Zhang, J.; Zhu, Y.; Wu, Y.; Yan, Q.; Liu, L.; Li, T. Activated Drp1-mediated mitochondrial ROS influence the gut microbiome and intestinal barrier after hemorrhagic shock. Aging 2020, 12, 1397–1416. [Google Scholar] [CrossRef]
- Yang, R.; Gallo, D.J.; Baust, J.J.; Watkins, S.K.; Delude, R.L.; Fink, M.P. Effect of hemorrhagic shock on gut barrier function and expression of stress-related genes in normal and gnotobiotic mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, R1263–R1274. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Xu, T.; Zhang, C.; Yu, W.Q.; Wei, T.; Zhang, B.; Chen, Q.; Qiu, J.Y.; Li, H.J.; et al. Role of TLR5 in the Translocation and Dissemination of Commensal Bacteria in the Intestine after Traumatic Hemorrhagic Shock. J. Immunol. Res. 2021, 2021, 6417658. [Google Scholar] [CrossRef]
- Treangen, T.J.; Wagner, J.; Burns, M.P.; Villapol, S. Traumatic Brain Injury in Mice Induces Acute Bacterial Dysbiosis Within the Fecal Microbiome. Front. Immunol. 2018, 9, 2757. [Google Scholar] [CrossRef] [Green Version]
- You, W.; Zhu, Y.; Wei, A.; Du, J.; Wang, Y.; Zheng, P.; Tu, M.; Wang, H.; Wen, L.; Yang, X. Traumatic Brain Injury Induces Gastrointestinal Dysfunction and Dysbiosis of Gut Microbiota Accompanied by Alterations of Bile Acid Profile. J. Neurotrauma 2022, 39, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Yuan, Q.; Li, Z.; Du, Z.; Wu, G.; Yu, J.; Hu, J. Translocation and Dissemination of Gut Bacteria after Severe Traumatic Brain Injury. Microorganisms 2022, 10, 2082. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Xu, L.; Song, S.; Fan, W.; Wu, Q.; Tong, X.; Yan, H. Oral Administration of Brain Protein Combined With Probiotics Induces Immune Tolerance Through the Tryptophan Pathway. Front. Mol. Neurosci. 2021, 14, 634631. [Google Scholar] [CrossRef]
- Bao, W.; Sun, Y.; Lin, Y.; Yang, X.; Chen, Z. An integrated analysis of gut microbiota and the brain transcriptome reveals host-gut microbiota interactions following traumatic brain injury. Brain Res. 2023, 1799, 148149. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, S.E.; Watts, L.T.; Burmeister, D.M.; Merrill, D.; Scroggins, S.; Zou, Y.; Lai, Z.; Grandhi, R.; Lewis, A.M.; Newton, L.M.; et al. Moderate Traumatic Brain Injury Alters the Gastrointestinal Microbiome in a Time-Dependent Manner. Shock 2019, 52, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Taraskina, A.; Ignatyeva, O.; Lisovaya, D.; Ivanov, M.; Ivanova, L.; Golovicheva, V.; Baydakova, G.; Silachev, D.; Popkov, V.; Ivanets, T.; et al. Effects of Traumatic Brain Injury on the Gut Microbiota Composition and Serum Amino Acid Profile in Rats. Cells 2022, 11, 1409. [Google Scholar] [CrossRef]
- Frankot, M.A.; O’Hearn, C.M.; Blancke, A.M.; Rodriguez, B.; Pechacek, K.M.; Gandhi, J.; Hu, G.; Martens, K.M.; Vonder Haar, C. Acute gut microbiome changes after traumatic brain injury are associated with chronic deficits in decision-making and impulsivity in male rats. Behav. Neurosci. 2023, 137, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Medel-Matus, J.S.; Lagishetty, V.; Santana-Gomez, C.; Shin, D.; Mowrey, W.; Staba, R.J.; Galanopoulou, A.S.; Sankar, R.; Jacobs, J.P.; Mazarati, A.M. Susceptibility to epilepsy after traumatic brain injury is associated with preexistent gut microbiome profile. Epilepsia 2022, 63, 1835–1848. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, K.; Hou, X.; Hou, L. The association of traumatic brain injury, gut microbiota and the corresponding metabolites in mice. Brain Res. 2021, 1762, 147450. [Google Scholar] [CrossRef]
- Wen, L.; You, W.; Wang, Y.; Zhu, Y.; Wang, H.; Yang, X. Investigating Alterations in Caecum Microbiota After Traumatic Brain Injury in Mice. J. Vis. Exp. 2019, 151, e59410. [Google Scholar] [CrossRef]
- Matharu, D.; Dhotre, D.; Balasubramanian, N.; Pawar, N.; Sagarkar, S.; Sakharkar, A. Repeated mild traumatic brain injury affects microbial diversity in rat jejunum. J. Biosci. 2019, 44, 120. [Google Scholar] [CrossRef]
- Angoa-Perez, M.; Zagorac, B.; Anneken, J.H.; Briggs, D.I.; Winters, A.D.; Greenberg, J.M.; Ahmad, M.; Theis, K.R.; Kuhn, D.M. Repetitive, mild traumatic brain injury results in a progressive white matter pathology, cognitive deterioration, and a transient gut microbiota dysbiosis. Sci. Rep. 2020, 10, 8949. [Google Scholar] [CrossRef] [PubMed]
- Houlden, A.; Goldrick, M.; Brough, D.; Vizi, E.S.; Lenart, N.; Martinecz, B.; Roberts, I.S.; Denes, A. Brain injury induces specific changes in the caecal microbiota of mice via altered autonomic activity and mucoprotein production. Brain Behav. Immun. 2016, 57, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.T.T.; Islam, M.; Das, P.; Gilbert, J.A.; Ho, K.J.; Schwulst, S.J. Differential Fecal Microbiome Dysbiosis after Equivalent Traumatic Brain Injury in Aged Versus Young Adult Mice. J. Exp. Neurol. 2021, 2, 120–130. [Google Scholar] [CrossRef]
- Sgro, M.; Iacono, G.; Yamakawa, G.R.; Kodila, Z.N.; Marsland, B.J.; Mychasiuk, R. Age matters: Microbiome depletion prior to repeat mild traumatic brain injury differentially alters microbial composition and function in adolescent and adult rats. PLoS ONE 2022, 17, e0278259. [Google Scholar] [CrossRef]
- Ma, E.L.; Smith, A.D.; Desai, N.; Cheung, L.; Hanscom, M.; Stoica, B.A.; Loane, D.J.; Shea-Donohue, T.; Faden, A.I. Bidirectional brain-gut interactions and chronic pathological changes after traumatic brain injury in mice. Brain Behav. Immun. 2017, 66, 56–69. [Google Scholar] [CrossRef]
- Zhang, L.M.; Zhang, D.X.; Zheng, W.C.; Hu, J.S.; Fu, L.; Li, Y.; Xin, Y.; Wang, X.P. CORM-3 exerts a neuroprotective effect in a rodent model of traumatic brain injury via the bidirectional gut-brain interactions. Exp. Neurol. 2021, 341, 113683. [Google Scholar] [CrossRef] [PubMed]
- Mazarati, A.; Medel-Matus, J.S.; Shin, D.; Jacobs, J.P.; Sankar, R. Disruption of intestinal barrier and endotoxemia after traumatic brain injury: Implications for post-traumatic epilepsy. Epilepsia 2021, 62, 1472–1481. [Google Scholar] [CrossRef]
- Rice, M.W.; Pandya, J.D.; Shear, D.A. Gut Microbiota as a Therapeutic Target to Ameliorate the Biochemical, Neuroanatomical, and Behavioral Effects of Traumatic Brain Injuries. Front. Neurol. 2019, 10, 875. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.R.; Sgro, M.; Yamakawa, G.R.; Li, C.; McDonald, S.J.; Sun, M.; Shultz, S.R.; Brady, R.D.; Mychasiuk, R. Gut microbiome depletion and repetitive mild traumatic brain injury differentially modify bone development in male and female adolescent rats. Bone Rep. 2021, 15, 101123. [Google Scholar] [CrossRef] [PubMed]
- Mercado, N.M.; Zhang, G.; Ying, Z.; Gomez-Pinilla, F. Traumatic brain injury alters the gut-derived serotonergic system and associated peripheral organs. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166491. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.W.; Rogers, M.B.; Gao, Y.; Vincent, G.; Firek, B.A.; Janesko-Feldman, K.; Vagni, V.; Kochanek, P.M.; Ozolek, J.A.; Mollen, K.P.; et al. Depletion of gut microbiota is associated with improved neurologic outcome following traumatic brain injury. Brain Res. 2020, 1747, 147056. [Google Scholar] [CrossRef]
- Celorrio, M.; Abellanas, M.A.; Rhodes, J.; Goodwin, V.; Moritz, J.; Vadivelu, S.; Wang, L.; Rodgers, R.; Xiao, S.; Anabayan, I.; et al. Gut microbial dysbiosis after traumatic brain injury modulates the immune response and impairs neurogenesis. Acta Neuropathol. Commun. 2021, 9, 40. [Google Scholar] [CrossRef]
- Opeyemi, O.M.; Rogers, M.B.; Firek, B.A.; Janesko-Feldman, K.; Vagni, V.; Mullett, S.J.; Wendell, S.G.; Nelson, B.P.; New, L.A.; Marino, E.; et al. Sustained Dysbiosis and Decreased Fecal Short-Chain Fatty Acids after Traumatic Brain Injury and Impact on Neurologic Outcome. J. Neurotrauma 2021, 38, 2610–2621. [Google Scholar] [CrossRef]
- Kaplan, G.B.; Leite-Morris, K.A.; Wang, L.; Rumbika, K.K.; Heinrichs, S.C.; Zeng, X.; Wu, L.; Arena, D.T.; Teng, Y.D. Pathophysiological Bases of Comorbidity: Traumatic Brain Injury and Post-Traumatic Stress Disorder. J. Neurotrauma 2018, 35, 210–225. [Google Scholar] [CrossRef]
- Laudani, S.; Torrisi, S.A.; Alboni, S.; Bastiaanssen, T.F.S.; Benatti, C.; Rivi, V.; Moloney, R.D.; Fuochi, V.; Furneri, P.M.; Drago, F.; et al. Gut microbiota alterations promote traumatic stress susceptibility associated with p-cresol-induced dopaminergic dysfunctions. Brain Behav. Immun. 2023, 107, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Tanelian, A.; Nankova, B.; Miari, M.; Nahvi, R.J.; Sabban, E.L. Resilience or susceptibility to traumatic stress: Potential influence of the microbiome. Neurobiol. Stress 2022, 19, 100461. [Google Scholar] [CrossRef]
- Tanelian, A.; Nankova, B.; Cheriyan, A.; Arens, C.; Hu, F.; Sabban, E.L. Differences in gut microbiota associated with stress resilience and susceptibility to single prolonged stress in female rodents. Neurobiol. Stress 2023, 24, 100533. [Google Scholar] [CrossRef]
- Hanscom, M.; Loane, D.J.; Shea-Donohue, T. Brain-gut axis dysfunction in the pathogenesis of traumatic brain injury. J. Clin. Investig. 2021, 131, e143777. [Google Scholar] [CrossRef]
- George, A.K.; Behera, J.; Homme, R.P.; Tyagi, N.; Tyagi, S.C.; Singh, M. Rebuilding Microbiome for Mitigating Traumatic Brain Injury: Importance of Restructuring the Gut-Microbiome-Brain Axis. Mol. Neurobiol. 2021, 58, 3614–3627. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.S.; Grandhi, R.; Patterson, T.T.; Nicholson, S.E. A Review of Traumatic Brain Injury and the Gut Microbiome: Insights into Novel Mechanisms of Secondary Brain Injury and Promising Targets for Neuroprotection. Brain Sci. 2018, 8, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krakovski, M.A.; Arora, N.; Jain, S.; Glover, J.; Dombrowski, K.; Hernandez, B.; Yadav, H.; Sarma, A.K. Diet-microbiome-gut-brain nexus in acute and chronic brain injury. Front. Neurosci. 2022, 16, 1002266. [Google Scholar] [CrossRef]
- Chiu, L.S.; Anderton, R.S. The role of the microbiota-gut-brain axis in long-term neurodegenerative processes following traumatic brain injury. Eur. J. Neurosci. 2023, 57, 400–418. [Google Scholar] [CrossRef]
- Mahajan, C.; Khurana, S.; Kapoor, I.; Sokhal, S.; Kumar, S.; Prabhakar, H.; Mathur, P.; Mani, K. Characteristics of Gut Microbiome After Traumatic Brain Injury. J. Neurosurg. Anesthesiol. 2023, 35, 86–90. [Google Scholar] [CrossRef]
- Urban, R.J.; Pyles, R.B.; Stewart, C.J.; Ajami, N.; Randolph, K.M.; Durham, W.J.; Danesi, C.P.; Dillon, E.L.; Summons, J.R.; Singh, C.K.; et al. Altered Fecal Microbiome Years after Traumatic Brain Injury. J. Neurotrauma 2020, 37, 1037–1051. [Google Scholar] [CrossRef] [Green Version]
- Brenner, L.A.; Stamper, C.E.; Hoisington, A.J.; Stearns-Yoder, K.A.; Stanislawksi, M.A.; Brostow, D.P.; Hoffmire, C.A.; Forster, J.E.; Schneider, A.L.; Postolache, T.T.; et al. Microbial Diversity and Community Structures Among Those With Moderate to Severe TBI: A United States-Veteran Microbiome Project Study. J. Head Trauma Rehabil. 2020, 35, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zayed, A.A.; Kigerl, K.A.; Zane, K.; Sullivan, M.B.; Popovich, P.G. Spinal Cord Injury Changes the Structure and Functional Potential of Gut Bacterial and Viral Communities. mSystems 2021, 6, e01356-20. [Google Scholar] [CrossRef]
- Kigerl, K.A.; Hall, J.C.; Wang, L.; Mo, X.; Yu, Z.; Popovich, P.G. Gut dysbiosis impairs recovery after spinal cord injury. J. Exp. Med. 2016, 213, 2603–2620. [Google Scholar] [CrossRef]
- Kang, J.N.; Sun, Z.F.; Li, X.Y.; Zhang, X.D.; Jin, Z.X.; Zhang, C.; Zhang, Y.; Wang, H.Y.; Huang, N.N.; Jiang, J.H.; et al. Alterations in gut microbiota are related to metabolite profiles in spinal cord injury. Neural. Regen. Res. 2023, 18, 1076–1083. [Google Scholar] [CrossRef]
- O’Connor, G.; Jeffrey, E.; Madorma, D.; Marcillo, A.; Abreu, M.T.; Deo, S.K.; Dietrich, W.D.; Daunert, S. Investigation of Microbiota Alterations and Intestinal Inflammation Post-Spinal Cord Injury in Rat Model. J. Neurotrauma 2018, 35, 2159–2166. [Google Scholar] [CrossRef] [PubMed]
- Doelman, A.; Tigchelaar, S.; McConeghy, B.; Sinha, S.; Keung, M.S.; Manouchehri, N.; Webster, M.; Fisk, S.; Morrison, C.; Streijger, F.; et al. Characterization of the gut microbiome in a porcine model of thoracic spinal cord injury. BMC Genom. 2021, 22, 775. [Google Scholar] [CrossRef]
- Rong, Z.; Huang, Y.; Cai, H.; Chen, M.; Wang, H.; Liu, G.; Zhang, Z.; Wu, J. Gut Microbiota Disorders Promote Inflammation and Aggravate Spinal Cord Injury Through the TLR4/MyD88 Signaling Pathway. Front. Nutr. 2021, 8, 702659. [Google Scholar] [CrossRef]
- Lee, J.; Lee, G.; Ko, G.; Joong Lee, S. Nerve injury-induced gut dysbiosis contributes to spinal cord TNF-alpha expression and nociceptive sensitization. Brain Behav. Immun. 2023, 110, 155–161. [Google Scholar] [CrossRef]
- Myers, S.A.; Gobejishvili, L.; Saraswat Ohri, S.; Garrett Wilson, C.; Andres, K.R.; Riegler, A.S.; Donde, H.; Joshi-Barve, S.; Barve, S.; Whittemore, S.R. Following spinal cord injury, PDE4B drives an acute, local inflammatory response and a chronic, systemic response exacerbated by gut dysbiosis and endotoxemia. Neurobiol. Dis. 2019, 124, 353–363. [Google Scholar] [CrossRef]
- Raue, K.D.; David, B.T.; Fessler, R.G. Spinal Cord-Gut-Immune Axis and Its Implications Regarding Therapeutic Development for Spinal Cord Injury. J. Neurotrauma 2023, 40, 793–806. [Google Scholar] [CrossRef]
- Pang, R.; Wang, J.; Xiong, Y.; Liu, J.; Ma, X.; Gou, X.; He, X.; Cheng, C.; Wang, W.; Zheng, J.; et al. Relationship between gut microbiota and lymphocyte subsets in Chinese Han patients with spinal cord injury. Front. Microbiol. 2022, 13, 986480. [Google Scholar] [CrossRef]
- Bazzocchi, G.; Turroni, S.; Bulzamini, M.C.; D’Amico, F.; Bava, A.; Castiglioni, M.; Cagnetta, V.; Losavio, E.; Cazzaniga, M.; Terenghi, L.; et al. Changes in gut microbiota in the acute phase after spinal cord injury correlate with severity of the lesion. Sci. Rep. 2021, 11, 12743. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, W.; Zhang, J.; Jing, Y.; Yang, M.; Du, L.; Gao, F.; Gong, H.; Chen, L.; Li, J.; et al. Gut microbiota dysbiosis in male patients with chronic traumatic complete spinal cord injury. J. Transl. Med. 2018, 16, 353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Jing, Y.; Zhang, W.; Zhang, J.; Yang, M.; Du, L.; Jia, Y.; Chen, L.; Gong, H.; Li, J.; et al. Dysbiosis of gut microbiota is associated with serum lipid profiles in male patients with chronic traumatic cervical spinal cord injury. Am. J. Transl. Res. 2019, 11, 4817–4834. [Google Scholar] [PubMed]
- Kong, G.; Zhang, W.; Zhang, S.; Chen, J.; He, K.; Zhang, C.; Yuan, X.; Xie, B. The gut microbiota and metabolite profiles are altered in patients with spinal cord injury. Mol. Brain 2023, 16, 26. [Google Scholar] [CrossRef] [PubMed]
- Gungor, B.; Adiguzel, E.; Gursel, I.; Yilmaz, B.; Gursel, M. Intestinal Microbiota in Patients with Spinal Cord Injury. PLoS ONE 2016, 11, e0145878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.; Xu, J.; Ma, Q.; Chen, M.; Wang, L.; Wen, S.; Yang, C.; Ma, C.; Wang, Y.; Luo, Q.; et al. Alterations in the fecal microbiota of patients with spinal cord injury. PLoS ONE 2020, 15, e0236470. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Van Der Pol, W.; Eraslan, M.; McLain, A.; Cetin, H.; Cetin, B.; Morrow, C.; Carson, T.; Yarar-Fisher, C. Comparison of the gut microbiome composition among individuals with acute or long-standing spinal cord injury vs. able-bodied controls. J. Spinal Cord Med. 2022, 45, 91–99. [Google Scholar] [CrossRef]
- Yu, B.; Qiu, H.; Cheng, S.; Ye, F.; Li, J.; Chen, S.; Zhou, L.; Yang, Y.; Zhong, C.; Li, J. Profile of gut microbiota in patients with traumatic thoracic spinal cord injury and its clinical implications: A case-control study in a rehabilitation setting. Bioengineered 2021, 12, 4489–4499. [Google Scholar] [CrossRef] [PubMed]
- Pattanakuhar, S.; Kaewchur, T.; Saiyasit, N.; Chattipakorn, N.; Chattipakorn, S.C. Level of injury is an independent determining factor of gut dysbiosis in people with chronic spinal cord injury: A cross-sectional study. Spinal Cord 2022, 60, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Kelly, L.S.; Apple, C.G.; Gharaibeh, R.; Pons, E.E.; Thompson, C.W.; Kannan, K.B.; Darden, D.B.; Efron, P.A.; Thomas, R.M.; Mohr, A.M. Stress-related changes in the gut microbiome after trauma. J. Trauma Acute Care Surg. 2021, 91, 192–199. [Google Scholar] [CrossRef]
- Nicholson, S.E.; Merrill, D.; Zhu, C.; Burmeister, D.M.; Zou, Y.; Lai, Z.; Darlington, D.N.; Lewis, A.M.; Newton, L.; Scroggins, S.; et al. Polytrauma independent of therapeutic intervention alters the gastrointestinal microbiome. Am. J. Surg. 2018, 216, 699–705. [Google Scholar] [CrossRef]
- Appiah, S.A.; Foxx, C.L.; Langgartner, D.; Palmer, A.; Zambrano, C.A.; Braumuller, S.; Schaefer, E.J.; Wachter, U.; Elam, B.L.; Radermacher, P.; et al. Evaluation of the gut microbiome in association with biological signatures of inflammation in murine polytrauma and shock. Sci. Rep. 2021, 11, 6665. [Google Scholar] [CrossRef]
- Yracheta, J.; Muraoka, W.; Wu, X.; Burmeister, D.; Darlington, D.; Zhao, D.; Lai, Z.; Sayyadioskoie, S.; Cap, A.P.; Bynum, J.; et al. Whole blood resuscitation restores intestinal perfusion and influences gut microbiome diversity. J. Trauma Acute Care Surg. 2021, 91, 1002–1009. [Google Scholar] [CrossRef]
- Munley, J.A.; Kelly, L.S.; Pons, E.E.; Kannan, K.B.; Coldwell, P.S.; Whitley, E.M.; Gillies, G.S.; Efron, P.A.; Nagpal, R.; Mohr, A.M. Multicompartmental traumatic injury and the microbiome: Shift to a pathobiome. J. Trauma Acute Care Surg. 2023, 94, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Munley, J.A.; Kelly, L.S.; Park, G.; Gillies, G.S.; Pons, E.E.; Kannan, K.B.; Whitley, E.M.; Bible, L.E.; Efron, P.A.; Nagpal, R.; et al. Multicompartmental Traumatic Injury Induces Sex-Specific Alterations in the Gut Microbiome. J. Trauma Acute Care Surg. 2023, 95, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Rupani, B.; Caputo, F.J.; Watkins, A.C.; Vega, D.; Magnotti, L.J.; Lu, Q.; Xu, D.Z.; Deitch, E.A. Relationship between disruption of the unstirred mucus layer and intestinal restitution in loss of gut barrier function after trauma hemorrhagic shock. Surgery 2007, 141, 481–489. [Google Scholar] [CrossRef]
- Wrba, L.; Ohmann, J.J.; Eisele, P.; Chakraborty, S.; Braumuller, S.; Braun, C.K.; Klohs, B.; Schultze, A.; von Baum, H.; Palmer, A.; et al. Remote Intestinal Injury Early After Experimental Polytrauma and Hemorrhagic Shock. Shock 2019, 52, e45–e51. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, D.M.; Johnson, T.R.; Lai, Z.; Scroggins, S.R.; DeRosa, M.; Jonas, R.B.; Zhu, C.; Scherer, E.; Stewart, R.M.; Schwacha, M.G.; et al. The gut microbiome distinguishes mortality in trauma patients upon admission to the emergency department. J. Trauma Acute Care Surg. 2020, 88, 579–587. [Google Scholar] [CrossRef]
- Howard, B.M.; Kornblith, L.Z.; Christie, S.A.; Conroy, A.S.; Nelson, M.F.; Campion, E.M.; Callcut, R.A.; Calfee, C.S.; Lamere, B.J.; Fadrosh, D.W.; et al. Characterizing the gut microbiome in trauma: Significant changes in microbial diversity occur early after severe injury. Trauma Surg. Acute Care Open 2017, 2, e000108. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, S.E.; Burmeister, D.M.; Johnson, T.R.; Zou, Y.; Lai, Z.; Scroggins, S.; DeRosa, M.; Jonas, R.B.; Merrill, D.R.; Zhu, C.; et al. A prospective study in severely injured patients reveals an altered gut microbiome is associated with transfusion volume. J. Trauma Acute Care Surg. 2019, 86, 573–582. [Google Scholar] [CrossRef]
- Balasubramanian, R.; Bazaz, M.R.; Pasam, T.; Sharief, N.; Velip, L.; Samanthula, G.; Dandekar, M.P. Involvement of Microbiome Gut-Brain Axis in Neuroprotective Effect of Quercetin in Mouse Model of Repeated Mild Traumatic Brain Injury. NeuroMolecular Med. 2023, 25, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Yanckello, L.M.; Fanelli, B.; McCulloch, S.; Xing, X.; Sun, M.; Hammond, T.C.; Colwell, R.; Gu, Z.; Ericsson, A.C.; Chang, Y.H.; et al. Inulin Supplementation Mitigates Gut Dysbiosis and Brain Impairment Induced by Mild Traumatic Brain Injury during Chronic Phase. J. Cell. Immunol. 2022, 4, 50–64. [Google Scholar] [CrossRef]
- Yanckello, L.M.; Chang, Y.H.; Sun, M.; Chlipala, G.; Green, S.J.; Lei, Z.; Ericsson, A.C.; Xing, X.; Hammond, T.C.; Bachstetter, A.D.; et al. Inulin supplementation prior to mild traumatic brain injury mitigates gut dysbiosis, and brain vascular and white matter deficits in mice. Front. Microbiomes 2022, 1, 986951. [Google Scholar] [CrossRef]
- Li, H.; Sun, J.; Du, J.; Wang, F.; Fang, R.; Yu, C.; Xiong, J.; Chen, W.; Lu, Z.; Liu, J. Clostridium butyricum exerts a neuroprotective effect in a mouse model of traumatic brain injury via the gut-brain axis. Neurogastroenterol. Motil 2018, 30, e13260. [Google Scholar] [CrossRef]
- Ma, Y.; Liu, T.; Fu, J.; Fu, S.; Hu, C.; Sun, B.; Fan, X.; Zhu, J. Lactobacillus acidophilus Exerts Neuroprotective Effects in Mice with Traumatic Brain Injury. J. Nutr. 2019, 149, 1543–1552. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Yang, D.; Bai, F.; Zhang, C.; Qin, C.; Li, D.; Wang, L.; Yang, M.; Chen, Z.; Li, J. Melatonin Treatment Alleviates Spinal Cord Injury-Induced Gut Dysbiosis in Mice. J. Neurotrauma 2019, 36, 2646–2664. [Google Scholar] [CrossRef] [PubMed]
- Rong, Z.J.; Cai, H.H.; Wang, H.; Liu, G.H.; Zhang, Z.W.; Chen, M.; Huang, Y.L. Ursolic Acid Ameliorates Spinal Cord Injury in Mice by Regulating Gut Microbiota and Metabolic Changes. Front. Cell. Neurosci. 2022, 16, 872935. [Google Scholar] [CrossRef]
- He, N.; Shen, G.; Jin, X.; Li, H.; Wang, J.; Xu, L.; Chen, J.; Cao, X.; Fu, C.; Shi, D.; et al. Resveratrol suppresses microglial activation and promotes functional recovery of traumatic spinal cord via improving intestinal microbiota. Pharmacol. Res. 2022, 183, 106377. [Google Scholar] [CrossRef]
- Tian, F.; Gao, X.; Zhang, L.; Wang, X.; Wan, X.; Jiang, T.; Wu, C.; Bi, J.; Lei, Q. Effects of n-3 PUFAs on Intestinal Mucosa Innate Immunity and Intestinal Microbiota in Mice after Hemorrhagic Shock Resuscitation. Nutrients 2016, 8, 609. [Google Scholar] [CrossRef] [Green Version]
- Brenner, L.A.; Forster, J.E.; Stearns-Yoder, K.A.; Stamper, C.E.; Hoisington, A.J.; Brostow, D.P.; Mealer, M.; Wortzel, H.S.; Postolache, T.T.; Lowry, C.A. Evaluation of an Immunomodulatory Probiotic Intervention for Veterans With Co-occurring Mild Traumatic Brain Injury and Posttraumatic Stress Disorder: A Pilot Study. Front. Neurol. 2020, 11, 1015. [Google Scholar] [CrossRef]
- Davis, B.T.T.; Chen, Z.; Islam, M.; Timken, M.E.; Procissi, D.; Schwulst, S.J. Fecal Microbiota Transfer Attenuates Gut Dysbiosis and Functional Deficits After Traumatic Brain Injury. Shock 2022, 57, 251–259. [Google Scholar] [CrossRef]
- Davis, B.T.T.; Chen, Z.; Islam, M.; Timken, M.E.; Procissi, D.; Schwulst, S.J. Postinjury Fecal Microbiome Transplant Decreases Lesion Size and Neuroinflammation in Traumatic Brain Injury. Shock 2022, 58, 287–294. [Google Scholar] [CrossRef]
- Du, D.; Tang, W.; Zhou, C.; Sun, X.; Wei, Z.; Zhong, J.; Huang, Z. Fecal Microbiota Transplantation Is a Promising Method to Restore Gut Microbiota Dysbiosis and Relieve Neurological Deficits after Traumatic Brain Injury. Oxid. Med. Cell. Longev. 2021, 2021, 5816837. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Yu, Y.; Bai, F.; Wang, L.; Yang, D.; Zhang, C.; Qin, C.; Yang, M.; Zhang, D.; Zhu, Y.; et al. Effect of fecal microbiota transplantation on neurological restoration in a spinal cord injury mouse model: Involvement of brain-gut axis. Microbiome 2021, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Bai, F.; Wang, L.; Yang, D.; Yan, Y.; Wang, Q.; Zhu, Y.; Yu, Y.; Chen, Z. Fecal Microbiota Transplantation Exerts Neuroprotective Effects in a Mouse Spinal Cord Injury Model by Modulating the Microenvironment at the Lesion Site. Microbiol. Spectr. 2022, 10, e0017722. [Google Scholar] [CrossRef] [PubMed]

| Title | Author and Year | Species | Trauma Model | Time Point(s) Studied Postinjury | Fecal Microbiome Diversity and Composition Changes | Intestinal Barrier Effects |
|---|---|---|---|---|---|---|
| Multicompartmental traumatic injury and the microbiome: shift to a pathobiome | Munley et al., 2023 [73] | Male Sprague-Dawley Rats | Lung contusion, hemorrhagic shock, laparotomy with cecectomy, and bifemoral pseudofracture (PT), or PT with CS (PT/CS) | 3, 7 days | ↓ Alpha-diversity (PT, 3 d) ↓ Alpha-diversity (PT/CS, 3 d, 7 d) Beta-diversity differences (3 d, 7 d) ↑ Enterococcus, Bacteroides, Parabacteroides (PT, 3 d) ↑ Bacteroides (PT/CS, 7 d) | ↑ Plasma occludin (PT/CS, 7 d) ↑ Colonic injury (inflammatory cell infiltrates, lamina propria edema) (PT and PT/CS, 7 d) |
| Multicompartmental Traumatic Injury Induces Sex-Specific Alterations in the Gut Microbiome | Munley et al., 2023 [74] | Male and Female Sprague-Dawley Rats | Lung contusion, hemorrhagic shock, laparotomy with cecectomy, and bifemoral pseudofracture (PT), or PT with CS | 2 days | ↓ Alpha-diversity (PT, PT/CS both sexes) Beta-diversity differences (PT, PT/CS both sexes) ↑ Blautia, Bacteroides (PT, both sexes) ↑ Parasutterella, Frisingicoccus (PT/CS, both sexes) | ↑ Plasma occludin (PT, PT/CS both sexes) ↑ Plasma lipopolysaccharide-binding protein (PT, PT/CS both sexes) ↑ Ileum injury (PT/CS, males) ↑ Colon injury (PT/CS, females) |
| Polytrauma independent of therapeutic intervention alters the gut microbiome | Nicholson et al., 2018 [70] | Male Sprague-Dawley Rats | Laparotomy with small intestine and liver crush injuries, femur fracture, skeletal muscle crush, HS (PT) | 2 h | ↔ Alpha-diversity Beta-diversity differences ↑ Deferribacteres, Lachnospiraceae, Mogibacteriaceae | NS |
| Whole blood resuscitation restores intestinal perfusion and influences gut microbiome diversity | Yracheta et al., 2021 [72] | Male Sprague-Dawley Rats | Laparotomy with liver crush injury, skeletal muscle crush injury, femur fracture (PT), HS | 2 h | ↔ Alpha-diversity Beta-diversity differences ↑ Roseburia, Enterobacteriaceae ↓ Rothia, Streptococcus | NS |
| Stress-related changes in the gut microbiome after trauma | Kelly et al., 2021 [69] | Male Sprague-Dawley Rats | LCHS, LCHS and 7 d CS (LCHS/CS 7/7), or LCHS and 14 d CS (LCHS/CS 14) | 3, 7, 14 days | ↔ Alpha-diversity (LCHS, all time points) ↑ Alpha-diversity (LCHS/CS 7/7, 3 d, 7 d) ↑ Alpha-diversity (LCHS/CS 14, 7 d, 14 d) ↔ Beta-diversity (7 d, 14 d) ↑ Ruminococcaceae (3 d), Bacteroides (7 d), Lactobacillus (14 d) (LCHS/CS 7/7) ↑ Clostridium (3 d), Bacteroides (7 d), Lachnospiraceae (14 d) (LCHS/CS 14) | NS |
| Relationship between disruption of the unstirred mucus layer and intestinal restitution in loss of gut barrier function after traumatic hemorrhagic shock | Rupani et al., 2007 [75] | Male Sprague-Dawley Rats | Laparotomy and sham shock (T/SS), or laparotomy and HS (T/HS) | 0, 30, 60, 180 min | NS | ↑ Villous injury in ileum (T/HS, 60 min) ↓ Villous height in ileum (T/HS, all time points) ↑ Enterocyte apoptosis in ileum (T/HS, 30, 60 min) ↓ Mucus layer in ileum (T/HS) ↑ Serum fluorescein isothiocyanate-dextran 4 (T/HS, 0, 60 and 180 min) |
| Remote intestinal injury early after experimental polytrauma and hemorrhagic shock | Wrba et al., 2019 [76] | Male C57BL/6 Mice | Hemorrhagic shock (HS), or bilateral chest trauma, closed head injury, femur fracture with soft tissue injury (PT), or PT and HS (PT/HS) | 2 h | NS | ↑ Plasma intestinal fatty-acid-binding protein (HS, PT, PT/HS) ↓ Zonula occludens protein 1 ileum, colon (PT/HS) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munley, J.A.; Kirkpatrick, S.L.; Gillies, G.S.; Bible, L.E.; Efron, P.A.; Nagpal, R.; Mohr, A.M. The Intestinal Microbiome after Traumatic Injury. Microorganisms 2023, 11, 1990. https://doi.org/10.3390/microorganisms11081990
Munley JA, Kirkpatrick SL, Gillies GS, Bible LE, Efron PA, Nagpal R, Mohr AM. The Intestinal Microbiome after Traumatic Injury. Microorganisms. 2023; 11(8):1990. https://doi.org/10.3390/microorganisms11081990
Chicago/Turabian StyleMunley, Jennifer A., Stacey L. Kirkpatrick, Gwendolyn S. Gillies, Letitia E. Bible, Philip A. Efron, Ravinder Nagpal, and Alicia M. Mohr. 2023. "The Intestinal Microbiome after Traumatic Injury" Microorganisms 11, no. 8: 1990. https://doi.org/10.3390/microorganisms11081990