
An Overview of the Use and Applications of Limosilactobacillus fermentum in Broiler Chickens

 , and
, and
Abstract
:1. Introduction
2. Properties of Limosilactobacillus fermentum

{kind=link}
| Strain | Origin | Functional Properties | References |
|---|---|---|---|
| L. fermentum YLF016 | Yak gut | High survival rate in the gut; strong adherence to intestinal cells; antibacterial and antioxidant effects; non-hemolytic activity | [53] |
| L. fermentum PC-10 | Poultry gut | Inhibition of S. Gallinarum growth | [56] |
| L. fermentum PG1 | Poultry digesta | Adhesion to the epithelial cells; survival at low pH; tolerance to bile salts; antibacterial activity | [57] |
| L. fermentum Y57 | Artisanal yogurt | Reduction of hypercholesterolemia in rats | [62] |
| L. fermentum GR-3 | Fermented food | Ameliorates human hyperuricemia via degrading and promoting excretion of uric acid | [63] |
| L. fermentum MBD93 | – | Adhesion to gastrointestinal mucin; exclusion of enteropathogenic bacteria | [71] |
| L. fermentum 10 | Human feces | Strong adhesion to * HT29 epithelial cells; high tolerance to bile salt; autoaggregation activity; reduction of E. coli adhesion; antibacterial and antioxidant activity | [75] |
| L. fermentum J23 | Cheese | Antimicrobial activity of bacteriocin-containing fractions; growth inhibition of E. coli, S. aureus, L. innocua, and S. Typhimurium | [82] |
| L. fermentum SD11 | Human oral cavity | Production of fermencin SD11; antibacterial activity against oral pathogens | [83] |
| L. fermentum BZ532 | Cereal beverage | Production of bacteriocin LF-BZ532 with a broad antimicrobial spectrum, including anti-listerial and anti-pseudomonas activity | [84] |
| L. fermentum LBM97 | Fermented vegetable | Production of bacteriocins LBM97-4 and LBM97-5 with antibacterial activity against S. aureus and E. coli | [85] |
| L. fermentum ME-3 | Human feces | Complete glutathione system; protection against oxidative stress | [90] |
| L. fermentum JX306 | Fermented vegetable | High scavenging activity of free and hydrogen radicals; improving glutathione peroxidase activity; effective inhibition of oxidative damage in liver and kidney | [92] |
| L. fermentum UCO-979C | Human gut | Inhibition of H. pylori growth and urease activity | [77,96] |
| L. fermentum DLBSA204 | Human breast milk | Macrophages activation; induction of nitric oxide synthesis; virus inactivation; downregulation of pro-inflammatory cytokines | [97] |
| L. fermentum IM12 | Human gut | Inhibition of NF-κB-STAT3 signaling pathway | [98] |
| L. fermentum AGR1487 | Human oral cavity | Capacity to activate TLR signaling pathway, immunomodulatory effects | [99] |
| L. fermentum CECT5716 | Human breast milk | High production of mucins; intestinal anti-inflammatory effects; immunomodulatory effects; alleviation of colitis-associated dysbiosis; glutathione-associated complex; mastitis prevention | [60,67,76,91,100,101] |
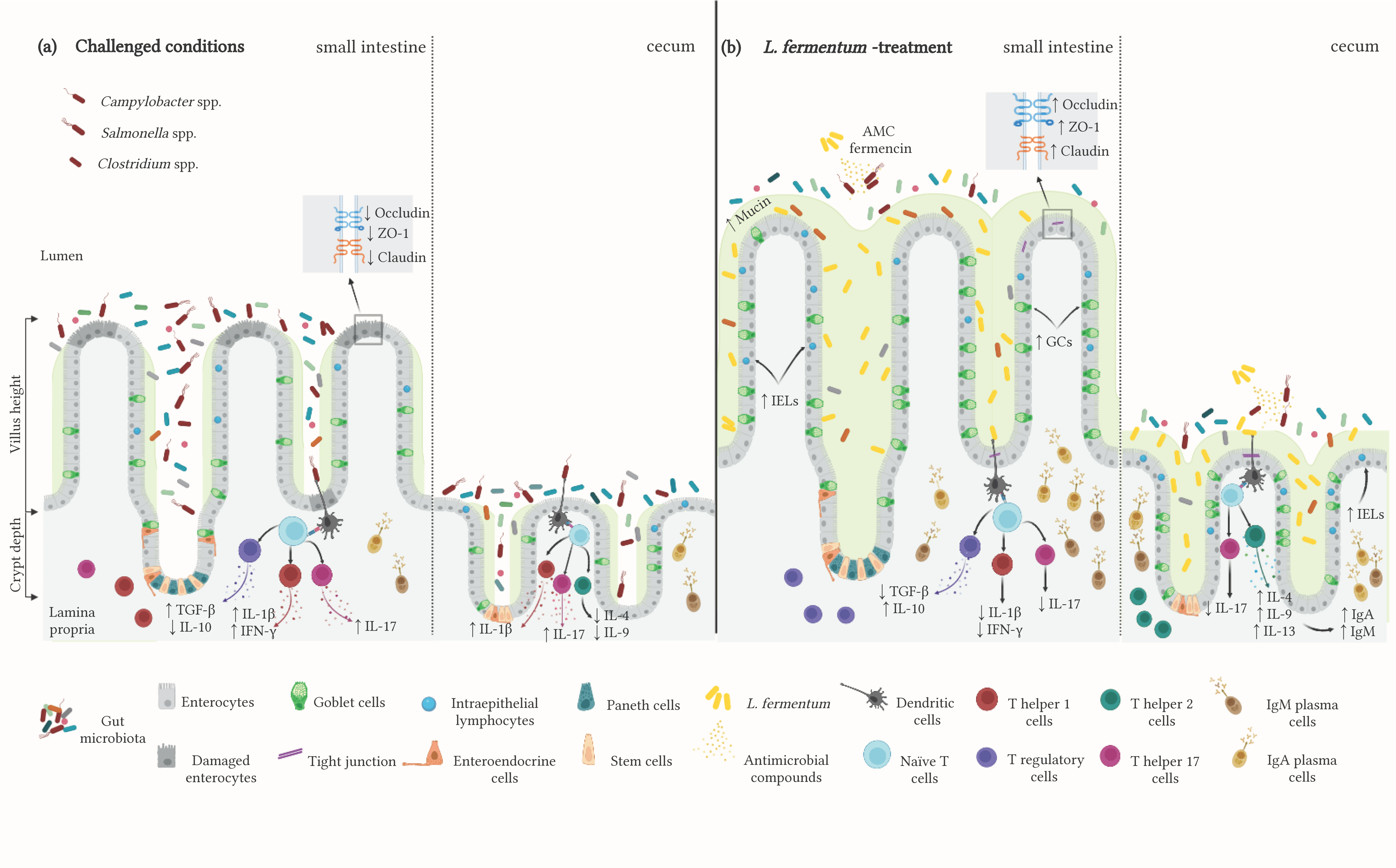
3. Applications of L. fermentum in Broiler Chickens
3.1. Gut Health, Microbiota, and Homeostasis
3.2. Modulation of Immune Reaction
3.3. Antagonism against Potentially Harmful Bacteria
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lin, J. Antibiotic growth promoters enhance animal production by targeting intestinal bile salt hydrolase and its producers. Front. Microbiol. 2014, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Cabello, F.C.; Godfrey, H.P. Even therapeutic antimicrobial use in animal husbandry may generate environmental hazards to human health. Environ. Microbiol. 2016, 18, 311–313. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Xu, S.; Tang, Z.; Li, Z.; Zhang, L. Use of antimicrobials in food animals and impact of transmission of antimicrobial resistance on humans. Biosaf. Health 2021, 3, 32–38. [Google Scholar] [CrossRef]
- European Parliament and the Council of the European Union. Regulation (EU) 2019/6 of the European Parliament and of the Council of 11 December 2018 on veterinary medicinal products and repealing Directive 2001/82/EC. OJEU 2019, 276, 43–167. [Google Scholar]
- Hicks, M.H. Results of new FDA rules on antibiotic use in US food-producing animals. J. Public Health 2020, 42, E573–E574. [Google Scholar] [CrossRef] [PubMed]
- Founou, L.L.; Founou, R.C.; Essack, S.Y. Antibiotic resistance in the food chain: A developing country-perspective. Front. Microbiol. 2016, 7, 1881. [Google Scholar] [CrossRef]
- Sugden, R.; Kelly, R.; Davies, S. Combatting antimicrobial resistance globally. Nat. Microbiol. 2016, 1, 16187. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and inputs from lactic acid bacteria and their bacteriocins as alternatives to antibiotic growth promoters during food-animal production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callaway, T.R.; Lillehoj, H.; Chuanchuen, R.; Gay, C.G. Alternatives to antibiotics: A symposium on the challenges and solutions for animal health and production. Antibiotics 2021, 10, 471. [Google Scholar] [CrossRef]
- Morais, T.; Inácio, A.; Coutinho, T.; Ministro, M.; Cotas, J.; Pereira, L.; Bahcevandziev, K. Seaweed potential in the animal feed: A review. J. Mar. Sci. Eng. 2020, 8, 559. [Google Scholar] [CrossRef]
- FAO/WHO. Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria. 2001. Available online: http://www.fao.org/3/a-a0512e.pdf (accessed on 10 April 2023).
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Fijan, S. Microorganisms with claimed probiotic properties: An overview of recent literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar] [CrossRef] [Green Version]
- Heeney, D.D.; Gareau, M.G.; Marco, M.L. Intestinal Lactobacillus in health and disease, a driver or just along for the ride? Curr. Opin. Biotechnol. 2018, 49, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Pessione, E. Lactic acid bacteria contribution to gut microbiota complexity: Lights and shadows. Front. Cell. Infect. Microbiol. 2012, 2, 86. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Qian, K.; Wang, C.; Wu, Y. Roles of probiotic lactobacilli inclusion in helping piglets establish healthy intestinal inter-environment for pathogen defense. Probiotics Antimicrob. Proteins 2018, 10, 243–250. [Google Scholar] [CrossRef]
- Stanley, D.; Hughes, R.J.; Moore, R.J. Microbiota of the chicken gastrointestinal tract: Influence on health, productivity and disease. Appl. Microbiol. Biotechnol. 2014, 98, 4301–4310. [Google Scholar] [CrossRef] [PubMed]
- Haakensen, M.; Dobson, C.M.; Hill, J.E.; Ziola, B. Reclassification of Pediococcus dextrinicus (Coster and White 1964) back 1978 (Approved Lists 1980) as Lactobacillus dextrinicus comb. nov., and emended description of the genus Lactobacillus. Int. J. Syst. Evol. Microbiol. 2009, 59, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Alp, D.; Kuleaşan, H. Adhesion mechanisms of lactic acid bacteria: Conventional and novel approaches for testing. World J. Microbiol. Biotechnol. 2019, 3, 156. [Google Scholar] [CrossRef]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [Green Version]
- Tojo, R.; Suarez, A.; Clemente, M.G.; de los Reyes-Gavilan, C.G.; Margolles, A.; Gueimonde, M.; Ruas-Madiedo, P. Intestinal microbiota in health and disease: Role of bifidobacteria in gut homeostasis. World J. Gastroenterol. 2014, 20, 15163–15176. [Google Scholar] [CrossRef]
- Ohland, C.L.; Macnaughton, W.K. Probiotic bacteria and intestinal epithelial barrier function. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G807–G819. [Google Scholar] [CrossRef] [Green Version]
- Woo, J.; Ahn, J. Probiotic–mediated competition, exclusion and displacement in biofilm formation by foodborne pathogens. Lett. Appl. Microbiol. 2013, 56, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Stone, W.; Tolmay, J.; Tucker, K.; Wolfaardt, G.M. Disinfectant, soap or probiotic cleaning? Surface microbiome diversity and biofilm competitive exclusion. Microorganisms 2020, 8, 1726. [Google Scholar]
- Dobson, A.; Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocin production: A probiotic trait? Appl. Environ. Microbiol. 2012, 78, 1–6. [Google Scholar]
- Atassi, F.; Servin, A.L. Individual and co-operative roles of lactic acid and hydrogen peroxide in the killing activity of enteric strain Lactobacillus johnsonii NCC933 and vaginal strain Lactobacillus gasseri KS120. 1 against enteric, uropathogenic and vaginosis-associated pathogens. FEMS Microbiol. Lett. 2010, 304, 29–38. [Google Scholar] [PubMed]
- Raheem, A.; Liang, L.; Zhang, G.; Cui, S. Modulatory effects of probiotics during pathogenic infections with emphasis on immune regulation. Front. Immunol. 2021, 12, 616713. [Google Scholar] [CrossRef]
- Habil, N.; Al-Murrani, W.; Beal, J.; Foey, A. Probiotic bacterial strains differentially modulate macrophage cytokine production in a strain-dependent and cell subset-specific manner. Benef. Microbes 2011, 2, 283–293. [Google Scholar]
- Feng, S.; Wang, S.; Qu, D.; Li, J.; Tian, F.; Yu, L.; Zhang, H.; Zhao, J.; Chen, W.; Zhai, Q. Species-or genus-dependent immunostimulatory effects of gut-derived potential probiotics. J. Genet. Genom 2022, in press.
- Evrard, B.; Coudeyras, S.; Dosgilbert, A.; Charbonnel, N.; Alamé, J.; Tridon, A.; Forestier, C. Dose-dependent immunomodulation of human dendritic cells by the probiotic Lactobacillus rhamnosus Lcr35. PLoS ONE 2011, 6, e18735. [Google Scholar]
- Ouwehand, A.C. A review of dose-responses of probiotics in human studies. Benef. Microbes 2017, 8, 143–151. [Google Scholar]
- Xu, Y.; Tian, Y.; Cao, Y.; Li, J.; Guo, H.; Su, Y.; Tian, Y.; Wang, C.; Wang, T.; Zhang, L. Probiotic properties of Lactobacillus paracasei subsp. Paracasei L1 and its growth performance-promotion in chicken by improving the intestinal microflora. Front. Physiol. 2019, 10, 937. [Google Scholar] [CrossRef]
- Zhao, Y.; Zeng, D.; Wang, H.; Qing, X.; Sun, N.; Xin, J.; Luo, M.; Khalique, A.; Pan, K.; Shu, G.; et al. Dietary probiotic Bacillus licheniformis H2 enhanced growth performance, morphology of small intestine and liver, and antioxidant capacity of broiler chickens against Clostridium perfringens–induced subclinical necrotic enteritis. Probiotics Antimicrob. Proteins 2020, 12, 883–895. [Google Scholar] [CrossRef]
- Amer, S.A.; Mohamed, W.A.M.; Gharib, H.S.A.; Al-Gabri, N.A.; Gouda, A.; Elabbasy, M.T.; Abd El-Rahman, G.I.; Omar, A.E. Changes in the growth, ileal digestibility, intestinal histology, behavior, fatty acid composition of the breast muscles, and blood biochemical parameters of broiler chickens by dietary inclusion of safflower oil and vitamin C. BMC Vet. Res. 2021, 17, 68. [Google Scholar] [CrossRef]
- Coudert, E.; Baéza, E.; Berri, C. Use of algae in poultry production: A review. Worlds Poult. Sci. J. 2020, 76, 767–786. [Google Scholar] [CrossRef]
- Šefcová, M.A.; Santacruz, F.; Larrea-Álvarez, C.M.; Vinueza-Burgos, C.; Ortega-Paredes, D.; Molina-Cuasapaz, G.; Rodríguez, J.; Calero-Cáceres, W.; Revajová, V.; Fernández-Moreira, E.; et al. Administration of dietary microalgae ameliorates intestinal parameters, improves body weight, and reduces thawing loss of fillets in broiler chickens: A pilot study. Animals 2021, 11, 3601. [Google Scholar] [CrossRef] [PubMed]
- Giang, H.H.; Viet, T.Q.; Ogle, B.; Lindberg, J.E. Growth performance, digestibility, gut environment and health status in weaned piglets fed a diet supplemented with a complex of lactic acid bacteria alone or in combination with Bacillus subtilis and Saccharomyces boulardii. Livest. Sci. 2012, 143, 132–141. [Google Scholar] [CrossRef]
- Wang, S.; Yao, B.; Gao, H.; Zang, J.; Tao, S.; Zhang, S.; Huang, S.; He, B.; Wang, J. Combined supplementation of Lactobacillus fermentum and Pediococcus acidilactici promoted growth performance, alleviated inflammation, and modulated intestinal microbiota in weaned pigs. BMC Vet. Res. 2019, 15, 239. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zijlstra, R.T.; Gänzle, M.G. Feeding Limosilactobacillus fermentum K9-2 and Lacticaseibacillus casei K9-1, or Limosilactobacillus reuteri tmw1. 656 reduces pathogen load in weanling pigs. Front. Microbiol. 2020, 11, 608293. [Google Scholar] [CrossRef]
- Nimalan, N.; Sørensen, S.L.; Fečkaninová, A.; Koščová, J.; Mudroňová, D.; Gancarčíková, S.; Vatsos, I.N.; Bisa, S.; Kiron, V.; Sørensen, M. Supplementation of lactic acid bacteria has positive effects on the mucosal health of Atlantic salmon (Salmo salar) fed soybean meal. Aquac. Rep. 2023, 28, 101461. [Google Scholar] [CrossRef]
- Ahmadifar, E.; Moghadam, M.S.; Dawood, M.A.; Hoseinifar, S.H. Lactobacillus fermentum and/or ferulic acid improved the immune responses, antioxidative defence and resistance against Aeromonas hydrophila in common carp (Cyprinus carpio) fingerlings. Fish Shellfish Immunol. 2019, 94, 916–923. [Google Scholar] [CrossRef]
- Krishnaveni, G.; Vignesh, S.; Vidhyalakshmi, N.; Vijay, V.; Ramesh, U. Effects of dietary supplementation of Lactobacillus fermentum URLP18 on growth, innate immunity and survival against Aeromonas hydrophila ATCC 7966 challenge in freshwater fish Cyprinus carpio (common carp). Aquac. Rep. 2021, 52, 1160–1176. [Google Scholar] [CrossRef]
- Madreseh, S.; Ghaisari, H.R.; Hosseinzadeh, S. Effect of lyophilized, encapsulated Lactobacillus fermentum and lactulose feeding on growth performance, heavy metals, and trace element residues in rainbow trout (Oncorhynchus mykiss) tissues. Probiotics Antimicrob. Proteins 2019, 11, 1257–1263. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Khan, A.; Ling, Z.; Wu, Y.; Feng, P.; Zhou, T.; Salama, E.; El-Dalatony, M.M.; Tian, X.; Liu, P.; et al. Feed-additive Limosilactobacillus fermentum GR-3 reduces arsenic accumulation in Procambarus clarkii. Ecotoxicol. Environ. Saf. 2022, 231, 113216. [Google Scholar] [CrossRef]
- Nguyen Thi Truc, L.; Trinh Ngoc, A.; Tran Thi Hong, T.; Nguyen Thanh, T.; Huynh Kim, H.; Pham Kim, L.; Huynh Truong, G.; Truong Quoc, P.; Nguyen Thi Ngoc, T. Selection of lactic acid bacteria (LAB) antagonizing Vibrio Parahaemolyticus: The pathogen of acute hepatopancreatic necrosis disease (AHPND) in whiteleg shrimp (Penaeus vannamei). Biology 2019, 8, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen Thi Truc, L.; Nguyen Thanh, T.; Tran Thi Hong, T.; Pham Van, D.; Vo Thi Tuyet, M.; Nguyen Trong, N.; Minh Phan, C.; Ngoc, D.C.; Truong Quoc, P. Effects of feed mixed with lactic acid bacteria and carbon, nitrogen, phosphorus supplied to the water on the growth and survival rate of white leg shrimp (Penaeus vannamei) infected with acute hepatopancreatic necrosis disease caused by Vibrio parahaemolyticus. Biology 2021, 10, 280. [Google Scholar] [PubMed]
- Lee, B.H.; Hsu, W.H.; Chen, Y.Z.; Hsu, K.T.; Pan, T.M. Limosilactobacillus fermentum SWP-AFFS02 improves the growth and survival rate of white shrimp via regulating immunity and intestinal microbiota. Fermentation 2021, 7, 179. [Google Scholar] [CrossRef]
- El Jeni, R.; Dittoe, D.K.; Olson, E.G.; Lourenco, J.; Corcionivoschi, N.; Ricke, S.C.; Callaway, T.R. Probiotics and potential applications for alternative poultry production systems. Poult. Sci. 2021, 100, 101156. [Google Scholar] [CrossRef]
- El-Hack, A.; Mohamed, E.; El-Saadony, M.T.; Shafi, M.E.; Qattan, S.Y.A.; Batiha, G.E.; Khafaga, A.F.; Abdel-Moneim, A.E.; Alagawany, M. Probiotics in poultry feed: A comprehensive review. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1835–1850. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Ksiezarek, M.; Grosso, F.; Gonçalves Ribeiro, T.; Peixe, L. Genomic diversity of genus Limosilactobacillus. Microb. Genom. 2022, 8, 000847. [Google Scholar] [CrossRef]
- Salvetti, E.; Harris, H.M.B.; Felis, G.E.; O’Toole, P.W. Comparative genomics of the genus Lactobacillus reveals robust phylogroups that provide the basis for reclassification. Appl. Environ. Microbiol. 2018, 84, e00993-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Ma, H.; Fakhar-e-Alam Kulyar, M.; Pan, H.; Li, K.; Li, A.; Mo, Q.; Wang, Y.; Dong, H.; Bao, Y.; et al. Complete genome analysis of Lactobacillus fermentum YLF016 and its probiotic characteristics. Microb. Pathog. 2022, 162, 105212. [Google Scholar] [CrossRef] [PubMed]
- Bintsis, T. Lactic acid bacteria: Their applications in foods. J. Bacteriol. Mycol. 2018, 6, 89–94. [Google Scholar]
- Duar, R.M.; Lin, X.B.; Zheng, J.; Martino, M.E.; Grenier, T.; Pérez-Muñoz, M.E.; Leulier, F.; Gänzle, M.; Walter, J. Lifestyles in transition: Evolution and natural history of the genus Lactobacillus. FEMS Microbiol. Rev. 2017, 41, S27–S48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adnan, M.; Nawaz, M.; Rabbani, M.; Mushtaq, M.H. In vitro characterization of probiotic potential of Limosilactobacillus fermentum against Salmonella Gallinarum causing fowl typhoid. Animals 2023, 13, 1284. [Google Scholar]
- Lin, W.H.; Yu, B.; Jang, S.H.; Tsen, H.Y. Different probiotic properties for Lactobacillus fermentum strains isolated from swine and poultry. Anaerobe 2007, 13, 107–113. [Google Scholar]
- Archer, A.C.; Halami, P.M. Probiotic attributes of Lactobacillus fermentum isolated from human feces and dairy products. Appl. Microbiol. Biotechnol. 2015, 99, 8113–8123. [Google Scholar] [CrossRef]
- López-Huertas, E. Safety and efficacy of human breast milk Lactobacillus fermentum CECT 5716. A mini-review of studies with infant formulae. Benef. Microbes 2015, 6, 219–224. [Google Scholar] [CrossRef]
- Hurtado, J.A.; Maldonado-Lobón, J.A.; Paz Díaz-Ropero, M.; Flores-Rojas, K.; Uberos, J.; Leante, J.L.; Affumicato, L.; Luz Couce, M.; Garrido, J.M.; Olivares, M.; et al. Oral administration to nursing women of Lactobacillus fermentum CECT5716 prevents lactational mastitis development: A randomized controlled trial. Breastfeeding Med. 2017, 12, 202–209. [Google Scholar]
- Mikelsaar, M.; Zilmer, M. Lactobacillus fermentum ME-3–an antimicrobial and antioxidative probiotic. Microb. Ecol. Health Dis. 2009, 21, 1–27. [Google Scholar]
- Zafar, H.; Ain, N.U.; Alshammari, A.; Alghamdi, S.; Raja, H.; Ali, A.; Siddique, A.; Tahir, S.D.; Akbar, S.; Arif, M.; et al. Lacticaseibacillus rhamnosus FM9 and Limosilactobacillus fermentum Y57 are as effective as statins at improving blood lipid profile in high cholesterol, high-fat diet model in male wistar rats. Nutrients 2022, 14, 1654. [Google Scholar] [CrossRef]
- Zhao, S.; Feng, P.; Hu, X.; Cao, W.; Liu, P.; Han, H.; Jin, W.; Li, X. Probiotic Limosilactobacillus fermentum GR-3 ameliorates human hyperuricemia via degrading and promoting excretion of uric acid. iScience 2022, 25, 105198. [Google Scholar] [CrossRef] [PubMed]
- FAO. Evaluation of the Generally Recognized as Safe (GRAS). Notice Inventory; 2020. Available online: https://www.fda.gov/media/154201/download (accessed on 14 May 2023).
- Zhao, Y.; Hong, K.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Lactobacillus fermentum and its potential immunomodulatory properties. J. Funct. Foods 2019, 56, 21–32. [Google Scholar] [CrossRef]
- Leuschner, R.G.K.; Robinson, T.P.; Hugas, M.; Cocconcelli, P.S.; Richard-Forget, F.; Klein, G.; Richardson, M. Qualified presumption of safety (QPS): A generic risk assessment approach for biological agents notified to the European Food Safety Authority (EFSA). Trends Food Sci. Technol. 2010, 21, 425–435. [Google Scholar]
- Bond, D.M.; Morris, J.M.; Nassar, N. Study protocol: Evaluation of the probiotic Lactobacillus fermentum CECT5716 for the prevention of mastitis in breastfeeding women: A randomised controlled trial. BMC Pregnancy Childbirth 2017, 17, 148. [Google Scholar]
- Panicker, A.S.; Ali, S.A.; Anand, S.; Panjagari, N.R.; Kumar, S.; Mohanty, A.K.; Behare, P.V. Evaluation of some in vitro probiotic properties of Lactobacillus fermentum strains. J. Food Sci. Technol. 2018, 55, 2801–2807. [Google Scholar] [CrossRef]
- Ali, S.A.; Singh, P.; Tomar, S.K.; Mohanty, A.K.; Behare, P. Proteomics fingerprints of systemic mechanisms of adaptation to bile in Lactobacillus fermentum. J. Proteom. 2020, 213, 103600. [Google Scholar] [CrossRef]
- Das, S.; Vishakha, K.; Banerjee, S.; Bera, T.; Mondal, S.; Ganguli, A. A novel probiotic strain of Lactobacillus fermentum TIU19 isolated from Haria beer showing both in vitro antibacterial and antibiofilm properties upon two multi resistant uro-pathogen strains. Curr. Res. Microb. Sci. 2022, 3, 100150. [Google Scholar] [CrossRef]
- Chatterjee, M.; Pushkaran, A.C.; Vasudevan, A.K.; Menon, K.K.N.; Biswas, R.; Mohan, C.G. Understanding the adhesion mechanism of a mucin binding domain from Lactobacillus fermentum and its role in enteropathogen exclusion. Int. J. Biol. Macromol. 2018, 110, 598–607. [Google Scholar] [CrossRef]
- Archer, A.C.; Kurrey, N.K.; Halami, P.M. In vitro adhesion and anti–inflammatory properties of native Lactobacillus fermentum and Lactobacillus delbrueckii spp. J. Appl. Microbiol. 2018, 125, 243–256. [Google Scholar] [CrossRef]
- Vélez, M.P.; de Keersmaecker, S.C.J.; Vanderleyden, J. Adherence factors of Lactobacillus in the human gastrointestinal tract. FEMS Microbiol. Lett. 2007, 276, 140–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falah, F.; Vasiee, A.; Behbahani, B.A.; Yazdi, F.T.; Moradi, S.; Mortazavi, S.A.; Roshanak, S. Evaluation of adherence and anti-infective properties of probiotic Lactobacillus fermentum strain 4-17 against Escherichia coli causing urinary tract infection in humans. Microb. Pathog. 2019, 131, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Gharbi, Y.; Fhoula, I.; Ruas-Madiedo, P.; Afef, N.; Boudabous, A.; Gueimonde, M.; Ouzari, H.I. In-vitro characterization of potentially probiotic Lactobacillus strains isolated from human microbiota: Interaction with pathogenic bacteria and the enteric cell line HT29. Ann. Microbiol. 2019, 69, 61–72. [Google Scholar] [CrossRef]
- Algieri, F.; Garrido-Mesa, J.; Vezza, T.; Rodriguez-Sojo, M.J.; Rodriguez-Cabezas, M.E.; Olivares, M.; Garcia, F.; Galvez, J.; Moron, R.; Rodriguez-Nogales, A. Intestinal anti-inflammatory effects of probiotics in DNBS-colitis via modulation of gut microbiota and microRNAs. Eur. J. Nutr. 2021, 60, 2537–2551. [Google Scholar] [CrossRef]
- García, A.; Navarro, K.; Sanhueza, E.; Pineda, S.; Pastene, E.; Quezada, M.; Henríquez, K.; Karlyshev, A.; Villena, J.; González, C. Characterization of Lactobacillus fermentum UCO-979C, a probiotic strain with a potent anti-Helicobacter pylori activity. Electron. J. Biotechnol. 2017, 25, 75–83. [Google Scholar] [CrossRef]
- Lehri, B.; Seddon, A.M.; Karlyshev, A.V. Lactobacillus fermentum 3872 as a potential tool for combatting Campylobacter jejuni infections. Virulence 2017, 8, 1753–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayashree, S.; Karthikeyan, R.; Nithyalakshmi, S.; Ranjani, J.; Gunasekaran, P.; Rajendhran, J. Anti-adhesion property of the potential probiotic strain Lactobacillus fermentum 8711 against methicillin-resistant Staphylococcus aureus (MRSA). Front. Microbiol. 2018, 9, 411. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Ramos, A.; Madi-Moussa, D.; Coucheney, F.; Drider, D. Current knowledge of the mode of action and immunity mechanisms of LAB-bacteriocins. Microorganisms 2021, 9, 2107. [Google Scholar] [CrossRef]
- Kumariya, R.; Garsa, A.K.; Rajput, Y.S.; Sood, S.K.; Akhtar, N.; Patel, S. Bacteriocins: Classification, synthesis, mechanism of action and resistance development in food spoilage causing bacteria. Microb. Pathog. 2019, 128, 171–177. [Google Scholar] [CrossRef]
- Heredia-Castro, P.Y.; Reyes-Díaz, R.; Rendón-Rosales, M.Á.; Beltrán-Barrientos, L.M.; Torres-Llanez, M.J.; Estrada-Montoya, M.C.; Hernández-Mendoza, A.; González-Córdova, A.F.; Vallejo-Cordoba, B. Novel bacteriocins produced by Lactobacillus fermentum strains with bacteriostatic effects in milk against selected indicator microorganisms. J. Dairy Sci. 2021, 104, 4033–4043. [Google Scholar] [CrossRef]
- Wannun, P.; Piwat, S.; Teanpaisan, R. Purification, characterization, and optimum conditions of fermencin SD11, a bacteriocin produced by human orally Lactobacillus fermentum SD11. Appl. Biochem. Biotechnol. 2016, 179, 572–582. [Google Scholar] [CrossRef]
- Rasheed, H.A.; Tuoheti, T.; Zhang, Y.; Azi, F.; Tekliye, M.; Dong, M. Purification and partial characterization of a novel bacteriocin produced by bacteriocinogenic Lactobacillus fermentum BZ532 isolated from Chinese fermented cereal beverage (Bozai). Lebensm. Wiss. Technol. 2020, 124, 109113. [Google Scholar] [CrossRef]
- Gu, Y.; Ismael, M.; Wang, X.; Liu, B.; Shan, Y.; Chen, Y.; Zhou, Y.; Yi, Y.; Lü, X. Mining and heterologous expression of bacteriocins from Limosilactobacillus fermentum LBM97. Food Biosci. 2021, 44, 101389. [Google Scholar] [CrossRef]
- Sánchez–Maldonado, A.F.; Schieber, A.; Gänzle, M.G. Structure–function relationships of the antibacterial activity of phenolic acids and their metabolism by lactic acid bacteria. J. Appl. Microbiol. 2011, 111, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, K.; Vadivoo, V.S.; Raj, G.D. Rapid determination of hydrogen peroxide produced by Lactobacillus using enzyme coupled rhodamine isocyanide/calcium phosphate nanoparticles. Biosens. Bioelectron. 2014, 61, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Bauer, G. Helicobacter pylori and reactive oxygen species. In Gastrointestinal Tissue, 1st ed.; Gracia-Sancho, J., Salvadó, J., Eds.; Academic Press: Cambridge, MA, USA, 2017; Chapter 6; pp. 81–97. [Google Scholar]
- Marciano, F.; Vajro, P. Oxidative stress and gut microbiota. In Gastrointestinal Tissue, 1st ed.; Gracia-Sancho, J., Salvadó, J., Eds.; Academic Press: Cambridge, MA, USA, 2017; Chapter 8; pp. 113–123. [Google Scholar]
- Kullisaar, T.; Songisepp, E.; Aunapuu, M.; Kilk, K.; Arend, A.; Mikelsaar, M.; Zilmer, M. Complete glutathione system in probiotic Lactobacillus fermentum ME-3. Appl. Biochem. Microbiol. 2010, 46, 481–486. [Google Scholar] [CrossRef]
- Surya, A.; Liu, X.; Miller, M.J. Glutathione utilization in Lactobacillus fermentum CECT 5716. J. Agric. Food Chem. 2018, 66, 12651–12656. [Google Scholar] [CrossRef]
- Zhang, D.I.; Li, C.; Shi, R.; Zhao, F.; Yang, Z. Lactobacillus fermentum JX306 restrain D-galactose-induced oxidative stress of mice through its antioxidant activity. Pol. J. Microbiol. 2020, 69, 205–215. [Google Scholar] [CrossRef]
- Masella, R.; Di Benedetto, R.; Varì, R.; Filesi, C.; Giovannini, C. Novel mechanisms of natural antioxidant compounds in biological systems: Involvement of glutathione and glutathione-related enzymes. J. Nutr. Biochem. 2005, 16, 577–586. [Google Scholar] [CrossRef]
- Appenzeller-Herzog, C. Glutathione- and non-glutathionebased oxidant control in the endoplasmic reticulum. J. Cell Sci. 2011, 124, 847–855. [Google Scholar] [CrossRef] [Green Version]
- Paulino do Nascimento, L.C.; Lacerda, D.C.; Ferreira, D.J.S.; de Souza, E.L.; de Brito Alves, J.L. Limosilactobacillus fermentum, current evidence on the antioxidant properties and opportunities to be exploited as a probiotic microorganism. Probiotics Antimicrob. Proteins 2022, 14, 960–979. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Castillo, V.; Zelaya, H.; Ilabaca, A.; Espinoza-Monje, M.; Komatsu, R.; Albarracín, L.; Kitazawa, H.; Garcia-Cancino, A.; Villena, J. Lactobacillus fermentum UCO-979C beneficially modulates the innate immune response triggered by Helicobacter pylori infection in vitro. Benef. Microbes 2018, 9, 829–841. [Google Scholar] [CrossRef] [PubMed]
- Wulandari, A.; Tandrasasmita, O.; Tjandrawinata, R. Immunomodulatory and macrophage activating activity of Lactobacillus fermentum DLBSA204 in response to respiratory infection in a cellular model. Biosci. Biotechnol. Res. Asia 2016, 13, 1291–1302. [Google Scholar] [CrossRef]
- Lim, S.M.; Jang, H.M.; Jang, S.E.; Han, M.J.; Kim, D.H. Lactobacillus fermentum IM12 attenuates inflammation in mice by inhibiting NF-κB-STAT3 signalling pathway. Benef. Microbes 2017, 8, 407–419. [Google Scholar] [CrossRef]
- Anderson, R.C.; Ulluwishewa, D.; Young, W.; Ryan, L.J.; Henderson, G.; Meijerink, M.; Maier, E.; Wells, J.M.; Roy, N.C. Human oral isolate Lactobacillus fermentum AGR1487 induces a pro-inflammatory response in germ-free rat colons. Sci. Rep. 2016, 6, 20318. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Abad, B.; Garrote, J.A.; Bernardo, D.; Montalvillo, E.; Escudero-Hernández, C.; Vázquez, E.; Arranz, E. Differential immunomodulatory effects of Lactobacillus rhamnosus DR20, Lactobacillus fermentum CECT 5716 and Bifidobacterium animalis subsp. lactis on monocyte-derived dendritic cells. J. Funct. Foods 2016, 22, 300–312. [Google Scholar] [CrossRef]
- Perez-Cano, F.J.; Dong, H.; Yaqoob, P. In vitro immunomodulatory activity of Lactobacillus fermentum CECT5716 and Lactobacillus salivarius CECT5713: Two probiotic strains isolated from human breast milk. Immunobiol. 2010, 215, 996–1004. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C.J. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar]
- Vissers, Y.M.; Snel, J.; Zuurendonk, P.F.; Kleerebezem, M.; Wichers, H.J.; Savelkoul, H.F. Lactobacillus strains differentially modulate cytokine production by hPBMC from pollen-allergic patients. FEMS Immunol. Med. Microbiol. 2011, 61, 28–40. [Google Scholar] [CrossRef]
- Mehdi, Y.; Létourneau-Montminy, M.P.; Gaucher, M.L.; Chorfi, Y.; Suresh, G.; Rouissi, T.; Kaur Brar, S.; Côté, C.; Avalos Ramirez, A.; Godbout, S. Use of antibiotics in broiler production: Global impacts and alternatives. Anim. Nutr. 2018, 4, 170–178. [Google Scholar] [CrossRef]
- Chapman, H.D.; Jeffers, T.K. Vaccination of chickens against coccidiosis ameliorates drug resistance in commercial poultry production. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 214–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Salem, H.M.; El-Tahan, A.M.; Soliman, M.M.; Youssef, G.B.; Taha, A.E.; Soliman, S.M.; Ahmed, A.E.; El-kott, A.F.; et al. Alternatives to antibiotics for organic poultry production: Types, modes of action and impacts on bird’s health and production. Poult. Sci. 2022, 101, 101696. [Google Scholar] [CrossRef] [PubMed]
- Šefcová, M.A.; Larrea-Álvarez, M.; Larrea-Álvarez, C.M.; Karaffová, V.; Ortega-Paredes, D.; Vinueza-Burgos, C.; Ševčíková, Z.; Levkut, M.; Herich, R.; Revajová, V. The probiotic Lactobacillus fermentum Biocenol CCM 7514 moderates Campylobacter jejuni-induced body weight impairment by improving gut morphometry and regulating cecal cytokine abundance in broiler chickens. Animals 2021, 11, 235. [Google Scholar] [CrossRef]
- Šefcová, M.A.; Ortega-Paredes, D.; Larrea-Álvarez, C.M.; Mina, I.; Guapás, V.; Ayala-Velasteguí, D.; Leoro-Garzón, P.; Molina-Cuasapaz, G.; Vinueza-Burgos, C.; Revajová, V.; et al. Effects of Lactobacillus fermentum administration on intestinal morphometry and antibody serum levels in Salmonella Infantis-challenged chickens. Microorganisms 2023, 11, 256. [Google Scholar] [CrossRef]
- Šefcová, M.; Larrea-Álvarez, M.; Larrea-Álvarez, C.; Karaffová, V.; Revajová, V.; Gancarčíková, S.; Ševčíková, Z.; Herich, R. Lactobacillus fermentum administration modulates cytokine expression and lymphocyte subpopulation levels in broiler chickens challenged with Campylobacter coli. Foodborne Pathog. Dis. 2020, 17, 485–493. [Google Scholar] [CrossRef]
- Šefcová, M.; Larrea-Álvarez, M.; Larrea-Álvarez, C.; Revajová, V.; Karaffová, V.; Koščová, J.; Nemcová, R.; Ortega-Paredes, D.; Vinueza-Burgos, C.; Levkut, M.; et al. Effects of Lactobacillus fermentum supplementation on body weight and pro-inflammatory cytokine expression in Campylobacter jejuni-challenged chickens. Vet. Sci. 2020, 7, 121. [Google Scholar] [CrossRef]
- Cao, L.; Yang, X.; Sun, F.; Liu, C.; Yao, J. Lactobacillus strain with high adhesion stimulates intestinal mucin expression in broiler. J. Poult. Sci. 2012, 49, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Yang, X.J.; Li, Z.J.; Sun, F.F.; Wu, X.H.; Yao, J.H. Reduced lesions in chickens with Clostridium perfringens-induced necrotic enteritis by Lactobacillus fermentum 1.2029. Poult. Sci. 2012, 91, 3065–3071. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Xi, Y.; Xia, Y.; Wu, T.; Zhao, D.; Zhang, Z.; Ding, B. Dietary Lactobacillus fermentum and Bacillus coagulans supplementation modulates intestinal immunity and microbiota of broiler chickens challenged by Clostridium perfringens. Front. Vet. Sci. 2021, 8, 680742. [Google Scholar] [CrossRef]
- Hati, S.; Mishra, B.K.; Patel, M.; Prajapati, J.B.; Bhagora, N.J.; Savaliya, F.P.; Pathan, M.; Purnima, B.J.; Ghodasara, D.J. Indigenous Lactobacillus strains improve growth performance and high-density cholesterol levels in broilers. Indian J. Exp. Biol 2021, 59, 556–563. [Google Scholar]
- Geeta, A.S.Y.; Pradhan, S.; Rajoria, R.; Kumar, A.; Gopi, M.; Navani, N.K.; Pathania, R. Probiotic attributes of Lactobacillus fermentum NKN51 isolated from yak cottage cheese and the impact of its feeding on growth, immunity, caecal microbiology and jejunal histology in the starter phase of broiler birds. Indian J. Anim. Res. 2021, 55, 451–456. [Google Scholar]
- Wang, M.; Hu, J.; Yu, H.; Li, W.; He, G.; Dong, J.; Liu, Y.; Shi, S. Lactobacillus fermentum 1.2133 display probiotic potential in vitro and protect against Salmonella pullorum in chicken of infection. Lett. Appl. Microbiol. 2023, 76, ovac041. [Google Scholar] [PubMed]
- Reuben, R.C.; Sarkar, S.L.; Ibnat, H.; Setu, M.A.A.; Roy, P.C.; Jahid, I.K. Novel multi-strain probiotics reduces Pasteurella multocida induced fowl cholera mortality in broilers. Sci. Rep. 2021, 11, 8885. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.Z.; Zeng, X.G.; Ding, X.L. Effects of replacing soybean meal with fermented rapeseed meal on performance, serum biochemical variables and intestinal morphology of broilers. Asian. Australas. J. Anim. Sci. 2012, 25, 1734–1741. [Google Scholar] [CrossRef] [PubMed]
- Chiang, G.; Lu, W.Q.; Piao, X.S.; Hu, J.K.; Gong, L.M.; Thacker, P.A. Effects of feeding solid-state fermented rapeseed meal on performance, nutrient digestibility, intestinal ecology and intestinal morphology of broiler chickens. Asian. Australas. J. Anim. Sci. 2009, 23, 263–271. [Google Scholar] [CrossRef]
- Capcarova, M.; Weiss, J.; Hrncar, C.; Kolesarova, A.; Pal, G. Effect of Lactobacillus fermentum and Enterococcus faecium strains on internal milieu, antioxidant status and body weight of broiler chickens. J. Anim. Physiol. Anim. Nutr. 2010, 94, e215–e224. [Google Scholar] [CrossRef]
- Khan, M.; Raoult, D.; Richet, H.; Lepidi, H.; La Scola, B. Growth-promoting effects of single-dose intragastrically administered probiotics in chickens. Br. Poult. Sci. 2007, 48, 732–735. [Google Scholar] [CrossRef]
- Bai, S.P.; Wu, A.M.; Ding, X.M.; Lei, Y.; Bai, J.; Zhang, K.Y.; Chio, J.S. Effects of probiotic-supplemented diets on growth performance and intestinal immune characteristics of broiler chickens. Poult. Sci. 2013, 92, 663–670. [Google Scholar] [CrossRef]
- Penha Filho, R.A.C.; Díaz, S.J.A.; Fernando, F.S.; Chang, Y.F.; Andreatti Filho, R.L.; Junior, A.B. Immunomodulatory activity and control of Salmonella Enteritidis colonization in the intestinal tract of chickens by Lactobacillus based probiotic. Vet. Immunol. Immunopathol. 2015, 167, 64–69. [Google Scholar] [CrossRef]
- Li, P.; Zheng, L.; Qi, Y.; Liu, Z.; Du, E.; Wei, J.; Zhang, Z.; Guo, S.; Ding, B. Dietary Lactobacillus fermentum and Lactobacillus paracasei improve the intestinal health of broilers challenged with coccidia and Clostridium perfringens. Front. Vet. Sci. 2022, 9, 1025677. [Google Scholar] [CrossRef]
- Collett, S.R. Nutrition and wet litter problems in poultry. Anim. Feed Sci. Technol. 2012, 173, 65–75. [Google Scholar] [CrossRef]
- Chen, J.; Tellez, G.; Richards, J.D.; Escobar, J. Identification of potential biomarkers for gut barrier failure in broiler chickens. Front. Vet. Sci. 2015, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Biasato, I.; Ferrocino, I.; Biasibetti, E.; Grego, E.; Dabbou, S.; Sereno, A.; Gai, F.; Gasco, L.; Schiavone, A.; Cocolin, L.; et al. Modulation of intestinal microbiota, morphology and mucin composition by dietary insect meal inclusion in free-range chickens. BMC Vet. Res. 2018, 14, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swaggerty, C.L.; Callaway, T.R.; Kogut, M.H.; Piva, A.; Grilli, E. Modulation of the immune response to improve health and reduce foodborne pathogens in poultry. Microorganisms 2019, 7, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio, L.A. Possibilities of early life programming in broiler chickens via intestinal microbiota modulation. Poult. Sci. 2019, 98, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kogut, M.H.; Lee, A.; Santin, E. Microbiome and pathogen interaction with the immune system. Poult. Sci. 2020, 99, 1906–1913. [Google Scholar] [CrossRef]
- Celi, P.; Gabai, G. Oxidant/antioxidant balance in animal nutrition and health: The role of protein oxidation. Front. Vet. Sci. 2015, 2, 48. [Google Scholar] [CrossRef] [Green Version]
- Sato, K. Molecular nutrition: Interaction of nutrients, gene regulations and performances. Anim. Sci. J. 2016, 87, 857–862. [Google Scholar] [CrossRef]
- Mishra, B.; Jha, R. Oxidative stress in the poultry gut: Potential challenges and interventions. Front. Vet. Sci. 2019, 6, 60. [Google Scholar] [CrossRef] [Green Version]
- Kogut, M.H.; Genovese, K.J.; Swaggerty, C.L.; He, H.; Broom, L. Inflammatory phenotypes in the intestine of poultry: Not all inflammation is created equal. Poult. Sci. 2018, 97, 2339–2346. [Google Scholar] [CrossRef]
- Barone, R.; Rappa, F.; Macaluso, F.; Caruso Bavisotto, C.; Sangiorgi, C.; Di Paola, G.; Marino Gammazza, A. Alcoholic liver disease: A mouse model reveals protection by Lactobacillus fermentum. Clin. Transl. Gastroenterol. 2016, 7, e138. [Google Scholar]
- Chen, X.; Zhao, X.; Wang, H.; Yang, Z.; Li, J.; Suo, H. Prevent effects of Lactobacillus fermentum HY01 on dextran sulfate sodium-induced colitis in mice. Nutrients 2017, 9, 545. [Google Scholar] [PubMed] [Green Version]
- Garcia-Castillo, V.; Komatsu, R.; Clua, P.; Indo, Y.; Takagi, M.; Salva, S.; Islam, M.A.; Alvarez, S.; Takahashi, H.; Garcia-Cancino, A.; et al. Evaluation of the immunomodulatory activities of the probiotic strain Lactobacillus fermentum UCO-979C. Front. Immunol. 2019, 10, 1376. [Google Scholar] [PubMed] [Green Version]
- Awad, W.A.; Hess, C.; Hess, M. Re-Thinking the chicken–Campylobacter jejuni interaction: A review. Avian Pathol. 2018, 47, 352–363. [Google Scholar] [PubMed]
- Connerton, P.L.; Richards, P.J.; Lafontaine, G.M.; O’kane, P.M.; Ghaffar, N.; Cummings, N.J.; Smith, D.L.; Fish, N.M.; Connerton, I.F. The effect of the timing of exposure to Campylobacter jejuni on the gut microbiome and inflammatory responses of broiler chickens. Microbiome 2018, 6, 88. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yan, X.; Han, D.; Liu, Y.; Song, W.; Tong, T.; Ma, Y. Lactobacillus casei DBN023 protects against jejunal mucosal injury in chicks infected with Salmonella pullorum CMCC-533. Res. Vet. Sci. 2019, 127, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Guo, Y.; Wang, Z. β-1, 3/1, 6-Glucan alleviated intestinal mucosal barrier impairment of broiler chickens challenged with Salmonella enterica serovar Typhimurium. Poult. Sci. 2013, 92, 1764–1773. [Google Scholar] [PubMed]
- Daneshmand, A.; Kermanshahi, H.; Mohammed, J.; Sekhavati, M.H.; Javadmanesh, A.; Ahmadian, M.; Alizadeh, M.; Razmyar, J.; Kulkarni, R.R. Intestinal changes and immune responses during Clostridium perfringens-induced necrotic enteritis in broiler chickens. Poult. Sci. 2022, 101, 101652. [Google Scholar]
- Jayaraman, S.; Thangavel, G.; Kurian, H.; Mani, R.; Mukkalil, R.; Chirakkal, H. Bacillus subtilis PB6 improves intestinal health of broiler chickens challenged with Clostridium perfringens-induced necrotic enteritis. Poult Sci. 2013, 92, 370–374. [Google Scholar]
- Singh, R.; Remington, B.; Blackall, P.; Turni, C. Epidemiology of fowl cholera in free range broilers. Avian Dis. 2014, 58, 124–128. [Google Scholar] [CrossRef] [PubMed]

| Strain | Dose | Administration | Main Results | References |
|---|---|---|---|---|
| L. fermentum Biocenol CCM 7514 | 1 × 109 CFU/0.2 mL | Orally | ↑ VH and ↑ VH:CD ratio in the small intestine; ↑ GC count in the duodenum and jejunum; positive correlation between gut architecture and BW in early stages ↑ mRNA expression of IL-4, IL-18, IL-13; ↓ mRNA expression of IL-15, IL-16, IL-17RA, IL-9, IL-6RA and CXCL-12; ↑ percentages of IgM and CD8 cells in the cecum of young chickens Antagonistic effects against C. jejuni, C. coli, and S. Infantis; attenuation of intestinal impairments and regulation of cecal inflammatory response | [107,108,109,110] |
| L. fermentum 1.2029 | 1 × 108 CFU/0.5 mL | Orogastrically | ↑ jejunal GC density; ↑ mRNA expression of muc2 in the jejunum and ileum of 21-d-old chickens | [111] |
| 1 × 108 CFU/mL | Orally | Lessening of C. perfringens-induced conditions; intestinal necrotic lesions not observed after treatment; ↑ mRNA expression of IL-10; ↓ mRNA expression of IFN-γ and TLR2 in 28-d-old chickens | [112] | |
| 1 × 109 CFU/kG | Dietary | Modulation of C. perfringens-stimulated expression of pro-inflammatory cytokines in the jejunum in 28-d-old chickens | [113] | |
| L. fermentum KGL4 and L. plantarum KGL3A complex | 1 × 108 CFU/mL | Dietary | ↓ fecal coliform and enterococci count; ↑ fecal lactobacilli count during initial growth phase; well-organized intestinal epithelial lining and villi structure; ↑ BW; ↓ LDL and ↑ HDL content in serum of 42-d-old chickens | [114] |
| L. fermentum NKN51 | 1 × 107 CFU/gM | Dietary | ↓ total count of E. coli; ↑ count of lactobacilli; ↑ VH, VW, VH:CD ratio and surface area in the jejunum; ↑ BW and ↓ FCR of 28-d-old chickens | [115] |
| L. fermentum 1.2133 | 2.5 × 108 CFU | Dietary | ↑ number of lactobacilli in the ileum and cecum; ↓ Salmonella counts in the cecum of 15-d-old chickens Lessening of intestinal lesions inflicted by S. Pullorum | [116] |
| L. fermentum (strain unspecified) | 1 × 108 CFU/kG | 1 Dietary | ↓ enterobacteria counts, ↑ lactobacilli counts in ileum and cecum; ↑ BW and ↓ FCR of 28-d-old chickens Reduced effects of P. multocida on intestinal microbiota; regulation of anti-inflammatory genes | [117] |
| L. fermentum CICC 20176 | approx. 5 log CFU/mL | 2 RSM fermentation | ↑ VH:CD ratio in the jejunum; ↑ concentration of serum IgG and IgM; no differences in growth performance of 21- and 42-d-old chickens | [118] |
| L. fermentum CGMCC 0843 | approx. 5 log CFU/mL | 3 RSM fermentation | ↑ percentages of dry matter digestibility in 42-d-old chickens; ↑ VH:CD ratio in the jejunum and ileum of 21- and 42-d-old chickens; ↑ lactobacilli count in the ceca and colon of 21- and 42-d-old chickens | [119] |
| L. fermentum CCM 7158 | 1 × 109 CFU | In drinking water | ↓ total antioxidant status; ↓ content of serum triglycerides; ↑ BW in 42-d-old chickens | [120] |
| L. fermentum CIP 102980 | 1 × 107 CFU/mL | Intragastrically | ↑ BW and ↓ FCR in 36-d-old chickens | [121] |
| L. fermentum JS and S. cerevisiae product | 1 × 107 CFU/g | Dietary | ↑ percentages of CD3, CD4, CD8 cells and ↑ mRNA expression of TLR2 and TLR4 in the jejunum of 21- and 42-d-old chickens; ↑ BW, ↓ FCR ratio during starter period | [122] |
| L. fermentum (strain unspecified) | 1 × 105 CFU | 4 Orally | Protective effects against S. Enteritidis infection; ↑ percentages of macrophages and CD4 cells; minimized lesions in the cecal tonsils in 11-d-old chickens | [123] |
| L. fermentum (strain unspecified) | 1 × 109 CFU/g | Dietary | C. perfringens-induced downregulation of ZO-1, Mucin-2, and Occludin in the jejunum of 13-d-old chickens relieved by probiotic administration | [124] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Racines, M.P.; Solis, M.N.; Šefcová, M.A.; Herich, R.; Larrea-Álvarez, M.; Revajová, V. An Overview of the Use and Applications of Limosilactobacillus fermentum in Broiler Chickens. Microorganisms 2023, 11, 1944. https://doi.org/10.3390/microorganisms11081944
Racines MP, Solis MN, Šefcová MA, Herich R, Larrea-Álvarez M, Revajová V. An Overview of the Use and Applications of Limosilactobacillus fermentum in Broiler Chickens. Microorganisms. 2023; 11(8):1944. https://doi.org/10.3390/microorganisms11081944
Chicago/Turabian StyleRacines, Maria Paula, Maria Nicole Solis, Miroslava Anna Šefcová, Róbert Herich, Marco Larrea-Álvarez, and Viera Revajová. 2023. "An Overview of the Use and Applications of Limosilactobacillus fermentum in Broiler Chickens" Microorganisms 11, no. 8: 1944. https://doi.org/10.3390/microorganisms11081944