
Acaricidal Biominerals and Mode-of-Action Studies against Adult Blacklegged Ticks, Ixodes scapularis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Mechanical Insecticides and Ticks
2.2. Dipping Assay
2.3. Tick Movement
2.4. Choice-Mortality Assay
2.5. Scanning Electron Microscopy (SEM)
2.6. Statistical Analysis
3. Results
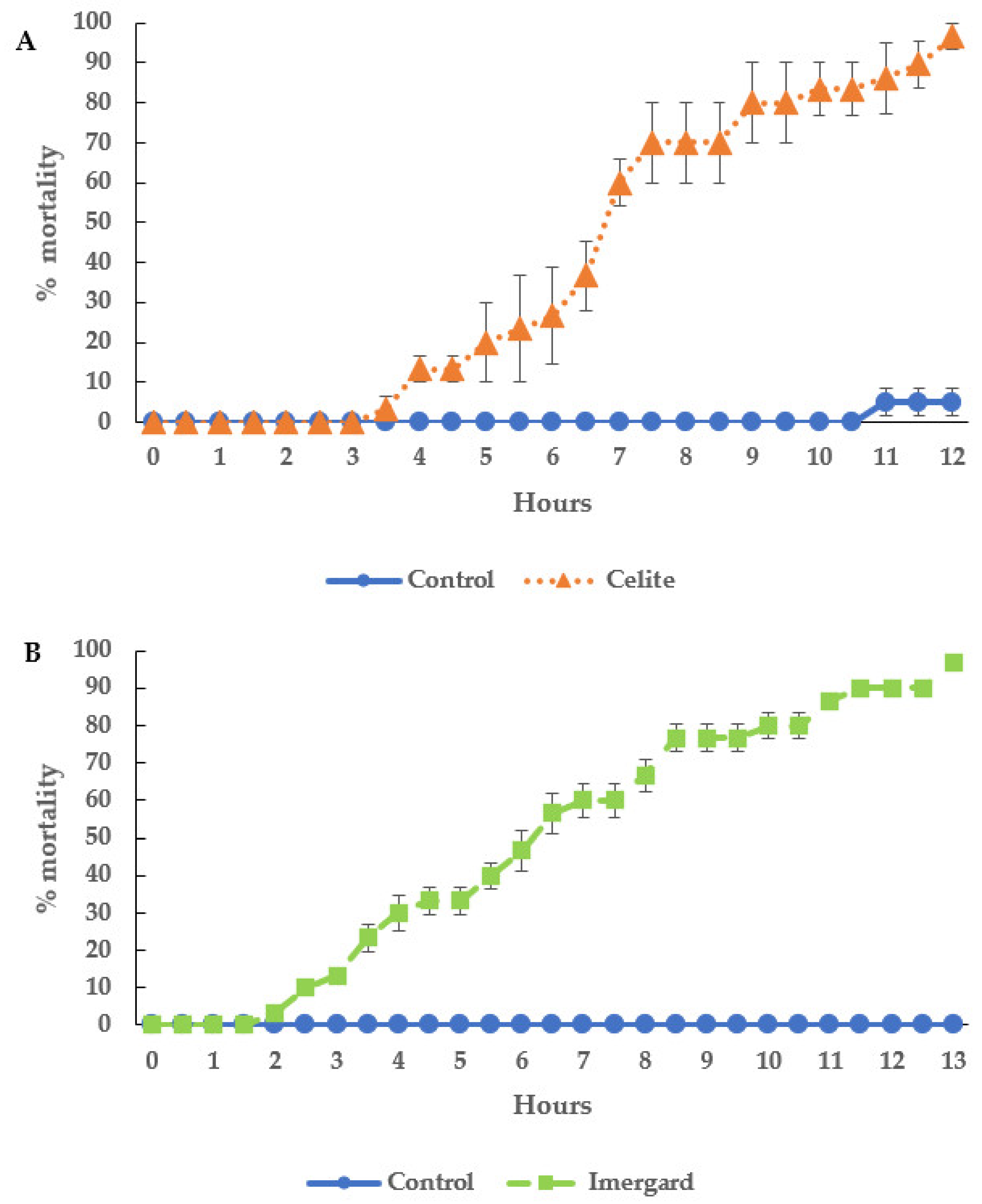
3.1. Mortality of Dipped I. scapularis Adults
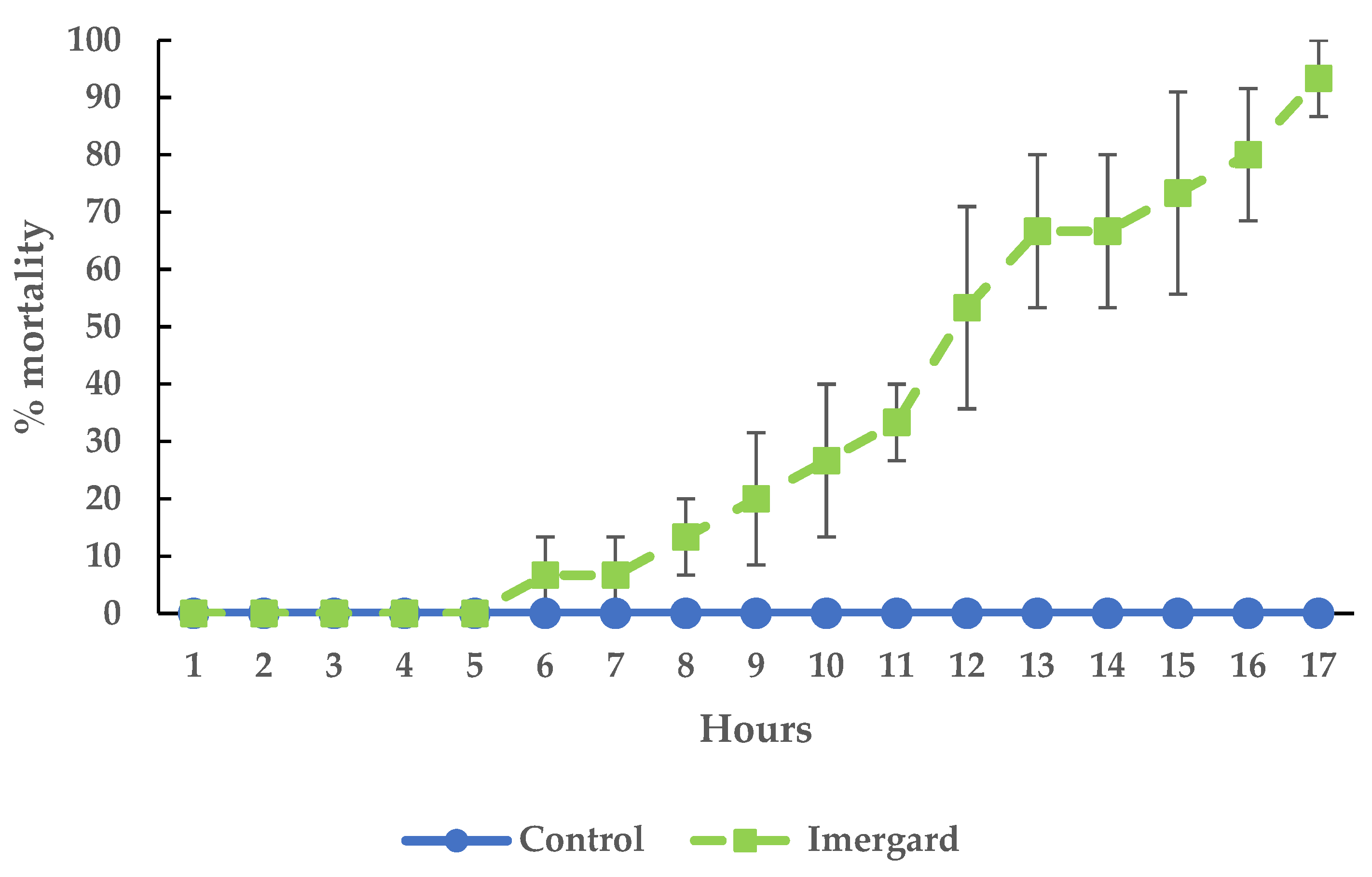
3.2. Mortality of I. scapularis Adults in the Choice Mortality Assay
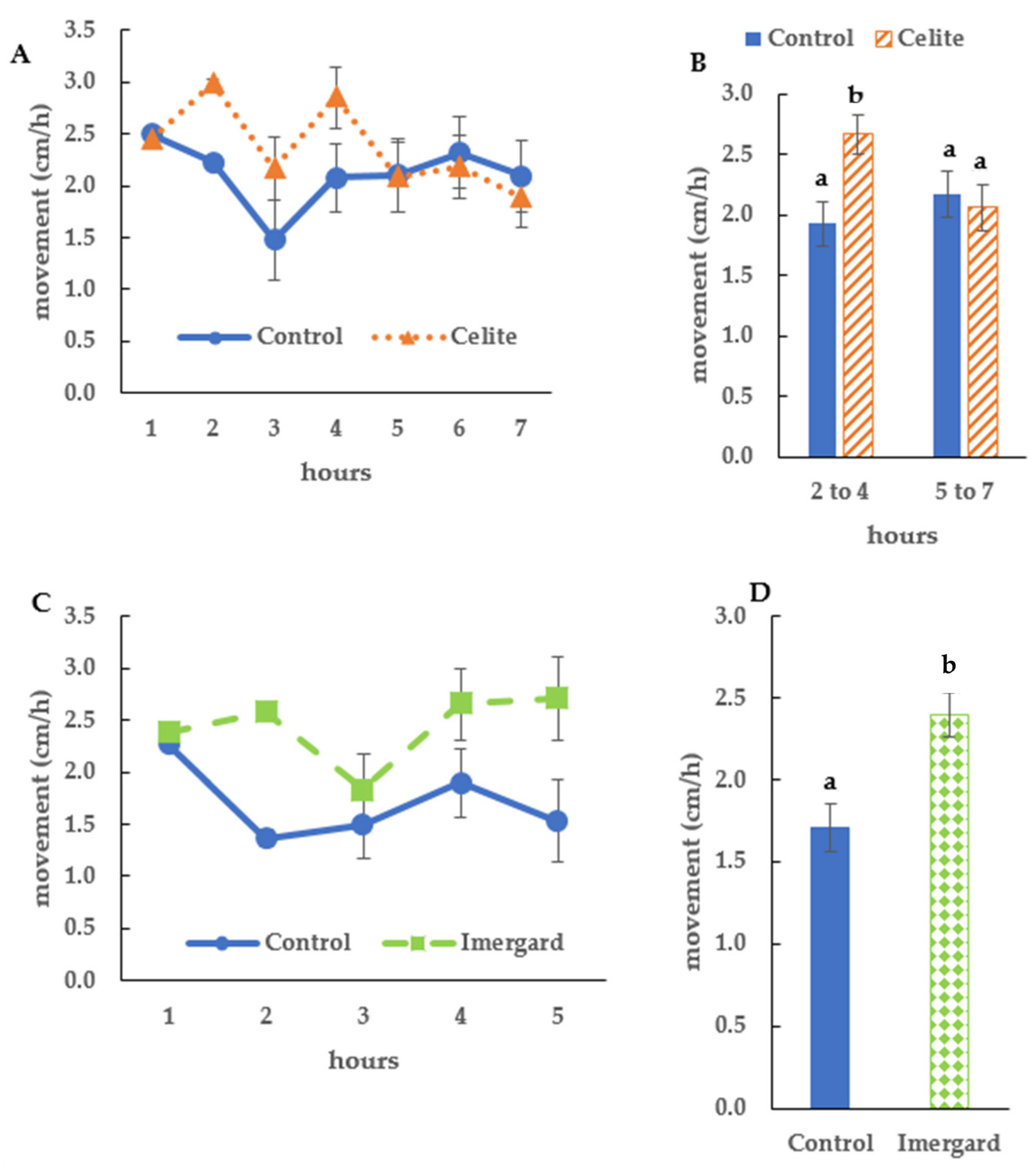
3.3. Mineral Exposure by Dipping on I. scapularis Movement
3.4. Scanning Electron Microscopy of I. scapularis Exposed to Imergard in the Choice Mortality Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adams, D.A.; Thomas, K.R.; Jajosky, R.A.; Foster, L.; Sharp, P.; Onweh, D.H.; Schley, A.W.; Anderson, W.J. Summary of Notifiable Infectious Diseases and Conditions—United States. Morb. Mortal. Wkly. Rep. 2014, 2016, 63. [Google Scholar]
- Eisen, R.J.; Eisen, L. The Blacklegged Tick, Ixodes scapularis: An Increasing Public Health Concern. Trends Parasitol. 2018, 34, 295–309. [Google Scholar] [CrossRef] [PubMed]
- CDC Lyme. Available online: https://www.cdc.gov/lyme/index.html (accessed on 15 April 2022).
- Eiden, A.L.; Kaufman, P.E.; Oi, F.M.; Allan, S.A.; Miller, R.J. Detection of permethrin resistance and fipronil tolerance in Rhipicephalus sanguineus (acari: Ixodidae) in the United States. J. Med. Entomol. 2015, 52, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Burtis, J.C.; Poggi, J.D.; Payne, B.; Campbell, S.R.; Harrington, L.C. Susceptibility of Ixodes scapularis (Acari: Ixodidae) to Permethrin under a Long-Term 4-Poster Deer Treatment Area on Shelter Island, NY. J. Med. Entomol. 2021, 58, 1966–1969. [Google Scholar] [CrossRef]
- Abbas, R.Z.; Zaman, M.A.; Colwell, D.D.; Gilleard, J.; Iqbal, Z. Acaricide resistance in cattle ticks and approaches to its management: The state of play. Vet. Parasitol. 2014, 203, 6–20. [Google Scholar] [CrossRef]
- Piesman, J.; Eisen, L. Prevention of tick-borne diseases. Annu. Rev. Entomol. 2008, 53, 323–343. [Google Scholar] [CrossRef]
- Stafford, K.C.; Williams, S.C.; Molaei, G. Integrated pest management in controlling ticks and tick-associated diseases. J. Integr. Pest Manag. 2017, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- Stafford, K.C. Tick Management Handbook; The Connecticut Agricultural Experiment Station: New Haven, CT, USA, 2004; p. 71. Available online: https://stacks.cdc.gov/view/cdc/11444 (accessed on 16 May 2023).
- Eisen, R.J.; Eisen, L.; Beard, C.B. County-scale distribution of Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae) in the continental United States. J. Med. Entomol. 2016, 53, 349–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beard, C.B.; Eisen, R.J.; Barker, C.M.; Garofalo, J.F.; Hahn, M.; Mary, H.; Monaghan, A.J.; Ogden, N.H.; Schramm, P.J. 5 Vector-Borne Diseases; Washington, DC, USA, 2016. Available online: https://health2016.globalchange.gov/low/ClimateHealth2016_05_Vector_small.pdf (accessed on 16 May 2023).
- Nabbout, A.E.; Ferguson, L.V.; Miyashita, A.; Adamo, S.A. Adult ticks (Ixodes scapularis) infected with B. burgdorferi have increased overwintering survival, with implications for tick population growth. bioRxiv 2022. [Google Scholar] [CrossRef]
- Deguenon, J.M.; Riegel, C.; Cloherty-Duvernay, E.R.; Chen, K.; Stewart, D.A.; Wang, B.; Gittins, D.; Tihomirov, L.; Apperson, C.S.; McCord, M.G.; et al. New mosquitocide derived from volcanic rock. J. Med. Entomol. 2021, 58, 458–464. [Google Scholar] [CrossRef]
- Deguenon, J.M.; Azondekon, R.; Agossa, F.R.; Padonou, G.G.; Anagonou, R.; Ahoga, J.; N’dombidje, B.; Akinro, B.; Stewart, D.A.; Wang, B.; et al. ImergardTMWP: A non-chemical alternative for an indoor residual spray, effective against pyrethroid-resistant Anopheles gambiae (s.l.) in Africa. Insects 2020, 11, 322. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Deguenon, J.M.; Cave, G.L.; Denning, S.S.; Reiskind, M.H.; Watson, D.W.; Stewart, D.A.; Gittins, D.; Zheng, Y.; Liu, X.; et al. New thinking for filth fly control: Residual, non-chemical wall spray from volcanic glass. Med. Vet. Entomol. 2021, 35, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Rahman, M.M. Diatomaceous Earth-Induced Alterations in the Reproductive Attributes in the Housefly Musca domestica L. (Diptera: Muscidae). SSRN Electron. J. 2021, 96, 41941–41944. [Google Scholar] [CrossRef]
- Korunić, Z.K.; Rozman, V.; Liška, A.; Lucić, P. A review of natural insecticides based on diatoaceous earths. Agriculture 2016, 22, 10–18. [Google Scholar] [CrossRef]
- Doggett, S.; Geary, M. The Efficacy of Diatomaceous Earth against the Common Bed Bug, Cimex lectularius; A Report for Mount Sylvia Diatomite Westmead Hospital, Sydney 2008. Available online: https://www.researchgate.net/publication/322974799_The_Efficacy_of_Diatomaceous_Earth_against_the_Common_Bed_Bug_Cimex_lectularius (accessed on 16 May 2023).
- Athanassiou, C.G.; Kavallieratos, N.G.; Chiriloaie, A.; Vassilakos, T.N.; Fătu, V.; Drosu, S.; Ciobanu, M.; Dudoiu, R. Insecticidal efficacy of natural diatomaceous earth deposits from Greece and Romania against four stored grain beetles: The effect of temperature and relative humidity. Bull. Insectology 2016, 69, 25–34. [Google Scholar]
- Arthur, F.H. Toxicity of diatomaceous earth to red flour beetles and confused flour beetles (Coleoptera: Tenebrionidae): Effects of temperature and relative humidity. J. Econ. Entomol. 2000, 93, 526–532. [Google Scholar] [CrossRef]
- le Patourel, G.N.J. The effect of grain moisture content on the toxicity of a sorptive silica dust to four species of grain beetle. J. Stored Prod. Res. 1986, 22, 63–69. [Google Scholar] [CrossRef]
- Athanassiou, C.G.; Kavallieratos, N.G.; Andris, N.S. Insecticidal effect of three diatomaceous earth formulations against adults of Sitophilus oryzae (Coleoptera: Curculionidae) and Tribolium confusum (Coleoptera: Tenebrionidae) on oat, rye, and triticale. J. Econ. Entomol. 2004, 97, 2160–2167. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Bazrafkan, S.; Vatandoost, H.; Abaei, M.R.; Ahmadi, M.S.; Tavassoli, M.; Mansoreh Shayeghi, R. The insecticidal effect of diatomaceous earth against adults and nymphs of Blattella germanica. Asian Pac. J. Trop. Biomed. 2014, 4, S228–S232. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, R.D.; Mott, D.W.; Dhammi, A.; Reisig, D.D.; Roe, R.M.; Stewart, D.A. Field Evaluation of a new thrips control agent for coton: A mechanical insecticide. In Proceedings of the Beltwide Cotton Conference, San Antonio, TX, USA, 3–5 January 2018; pp. 786–795. [Google Scholar]
- Showler, A.T.; Flores, N.; Caesar, R.M.; Mitchel, R.D.; De León, A.A.P. Lethal effects of a commercial diatomaceous earth dust product on Amblyomma americanum (ixodida: Ixodidae) larvae and nymphs. J. Med. Entomol. 2020, 57, 1575–1581. [Google Scholar] [CrossRef] [Green Version]
- Showler, A.T.; Harlien, J.L. Lethal Effects of Imergard WP, a Perlite-based Dust, on Amblyomma americanum (Ixodida: Ixodidae) Larvae and Nymphs. J. Med. Entomol. 2023, 60, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Richardson, E.A.; Ponnusamy, L.; Roe, R.M. Mechanical Acaricides Active against the Blacklegged Tick, Ixodes scapularis. Insects 2022, 13, 672. [Google Scholar] [CrossRef] [PubMed]
- Showler, A.T.; Harlien, J.L. Desiccant Dusts, With and Without Bioactive Botanicals, Lethal to Rhipicephalus (Boophilus) microplus Canestrini (Ixodida: Ixodidae) in the Laboratory and on Cattle. J. Med. Entomol. 2023, 60, 346–355. [Google Scholar] [CrossRef]
- Sonenshine, D.E.; Roe, R.M. Biology of Ticks, 2nd ed.; Sonenshine, D.E., Roe, R.M., Eds.; Oxford Unversity Press: New York, NY, USA, 2014. [Google Scholar]
- Dawson, R.D. Efficacy of diatomaceous earth at reducing populations of nest-dwelling ectoparasites in Tree Swallows. J. Field Ornithol. 2004, 75, 232–238. [Google Scholar] [CrossRef]
- Yoder, J.A.; Selim, M.E.; Needham, G.R. Impact of feeding, molting and relative humidity on cuticular wax deposition and water loss in the lone star tick, Amblyomma americanum. J. Insect Physiol. 1997, 43, 547–551. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mechanical Insecticide | N | Slope (SEM) | LT50 † (95% CL) | LT90 † (95% CL) | Χ2 |
|---|---|---|---|---|---|
| Celite 610 dip | 30 | 0.399 (0.0246) | 7.29 A ‡ (6.98–7.60) | 10.5 A ‡ (10.0–11.1) | 17.56 |
| Imergard dip | 30 | 0.358 (0.0310) | 6.72 A (6.35–7.09) | 11.2 A (10.6–11.9) | 8.90 |
| Imergard choice-mortality | 15 | 0.280 (0.0321) | 11.2 B (10.5–12.0) | 15.8 B (14.6–17.5) | 2.32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cave, G.L.; Richardson, E.A.; Chen, K.; Watson, D.W.; Roe, R.M. Acaricidal Biominerals and Mode-of-Action Studies against Adult Blacklegged Ticks, Ixodes scapularis. Microorganisms 2023, 11, 1906. https://doi.org/10.3390/microorganisms11081906
Cave GL, Richardson EA, Chen K, Watson DW, Roe RM. Acaricidal Biominerals and Mode-of-Action Studies against Adult Blacklegged Ticks, Ixodes scapularis. Microorganisms. 2023; 11(8):1906. https://doi.org/10.3390/microorganisms11081906
Chicago/Turabian StyleCave, Grayson L., Elise A. Richardson, Kaiying Chen, David W. Watson, and R. Michael Roe. 2023. "Acaricidal Biominerals and Mode-of-Action Studies against Adult Blacklegged Ticks, Ixodes scapularis" Microorganisms 11, no. 8: 1906. https://doi.org/10.3390/microorganisms11081906