
Optimizing Crop Production with Bacterial Inputs: Insights into Chemical Dialogue between Sphingomonas sediminicola and Pisum sativum

 , , and
, , and 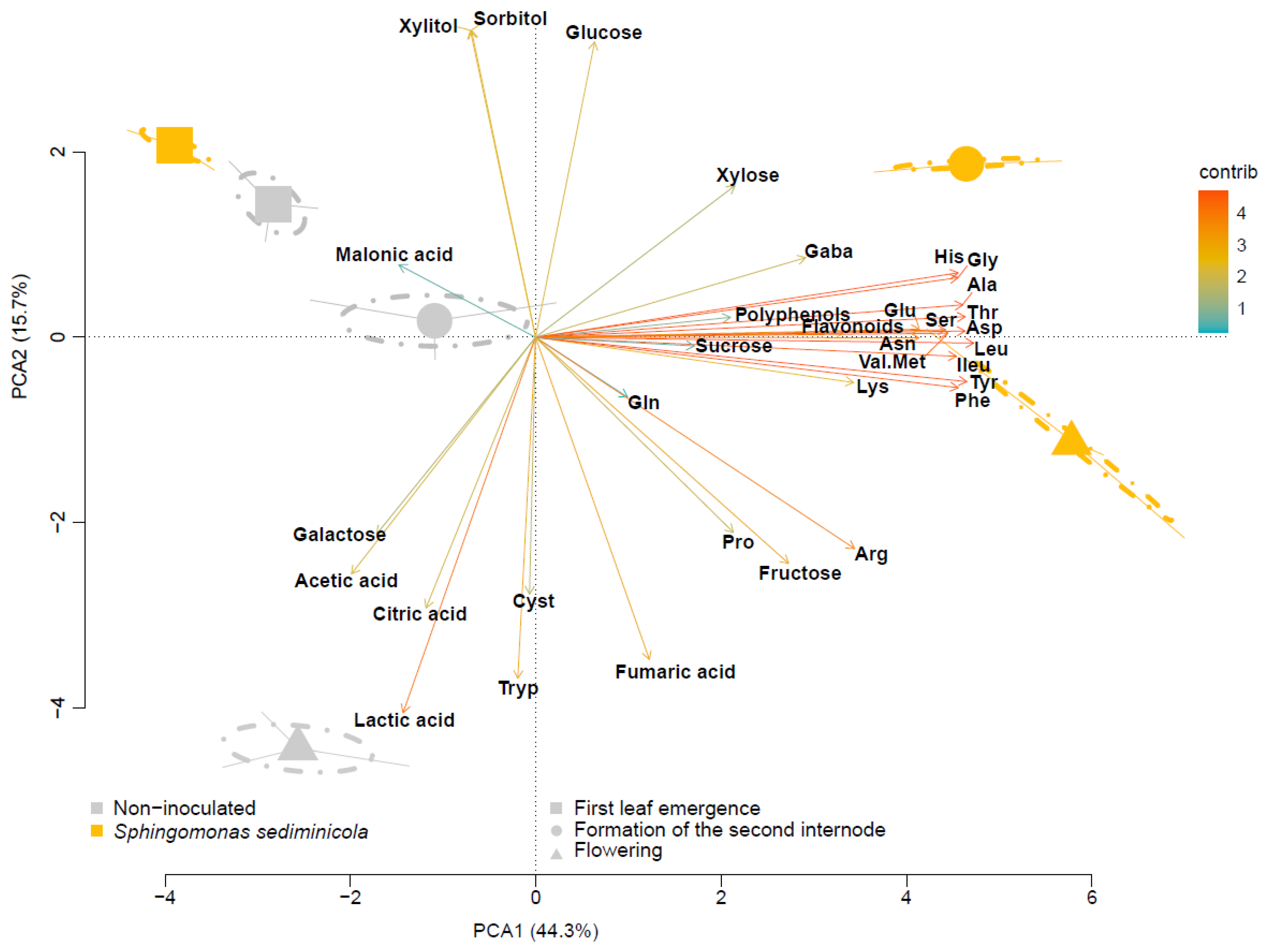{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Bacteria Culture
2.2. Hydroponic Growth Conditions
2.3. Extraction and Analysis of Hydroponic Compounds
2.4. Microarray Plates for Phenotypic Characterization of S. sediminicola
2.5. Untargeted Metabolomics of Sphingomonas sediminicola Culture Medium
2.6. Data Analysis
3. Results
3.1. The Composition of the Hydroponic Solution Is Influenced by S. sediminicola and the Stage of Pea Development
3.2. S. sediminicola Had a Specialized Carbon Metabolism and a Generalist Nitrogen Metabolism
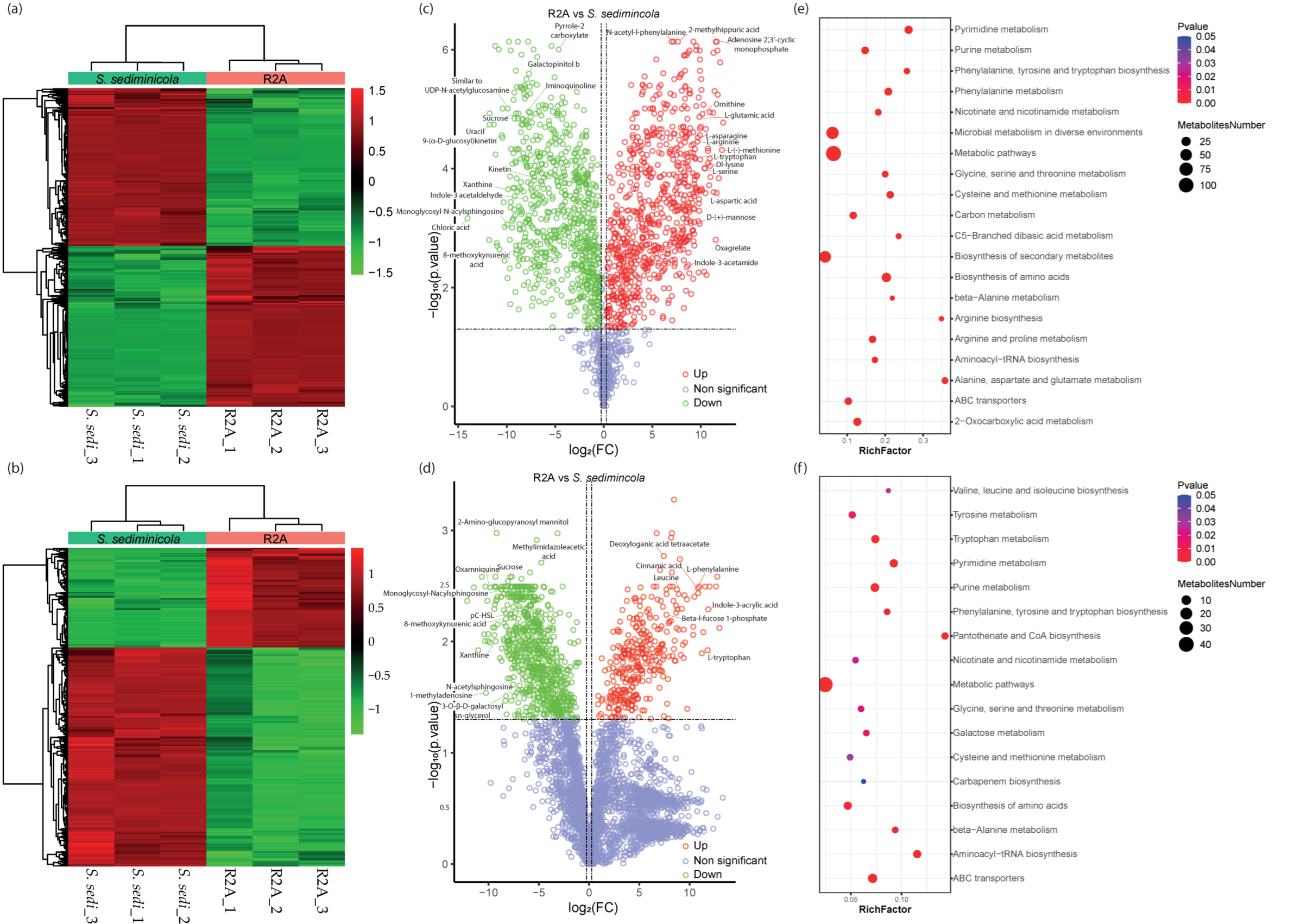
3.3. Untargeted Metabolomics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumari, B.; Mallick, M.A.; Solanki, M.K.; Solanki, A.C.; Hora, A.; Guo, W. Plant Growth Promoting Rhizobacteria (PGPR): Modern Prospects for Sustainable Agriculture. In Plant Health Under Biotic Stress; Ansari, R.A., Mahmood, I., Eds.; Volume 2: Microbial Interactions; Springer: Singapore, 2019; pp. 109–127. ISBN 9789811360404. [Google Scholar]
- Oleńska, E.; Małek, W.; Wójcik, M.; Swiecicka, I.; Thijs, S.; Vangronsveld, J. Beneficial Features of Plant Growth-Promoting Rhizobacteria for Improving Plant Growth and Health in Challenging Conditions: A Methodical Review. Sci. Total Environ. 2020, 743, 140682. [Google Scholar] [CrossRef]
- Khan, A.L.; Waqas, M.; Kang, S.-M.; Al-Harrasi, A.; Hussain, J.; Al-Rawahi, A.; Al-Khiziri, S.; Ullah, I.; Ali, L.; Jung, H.-Y.; et al. Bacterial Endophyte Sphingomonas Sp. LK11 Produces Gibberellins and IAA and Promotes Tomato Plant Growth. J. Microbiol. 2014, 52, 689–695. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate Solubilizing Microbes: Sustainable Approach for Managing Phosphorus Deficiency in Agricultural Soils. SpringerPlus 2013, 2, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etesami, H.; Adl, S.M. Plant Growth-Promoting Rhizobacteria (PGPR) and Their Action Mechanisms in Availability of Nutrients to Plants. In Phyto-Microbiome in Stress Regulation; Kumar, M., Kumar, V., Prasad, R., Eds.; Environmental and Microbial Biotechnology; Springer: Singapore, 2020; pp. 147–203. ISBN 9789811525766. [Google Scholar]
- Shah, A.; Nazari, M.; Antar, M.; Msimbira, L.A.; Naamala, J.; Lyu, D.; Rabileh, M.; Zajonc, J.; Smith, D.L. PGPR in Agriculture: A Sustainable Approach to Increasing Climate Change Resilience. Front. Sustain. Food Syst. 2021, 5, 667546. [Google Scholar] [CrossRef]
- Geddes, B.A.; Kearsley, J.; Morton, R.; diCenzo, G.C.; Finan, T.M. Chapter Eight—The Genomes of Rhizobia. In Advances in Botanical Research; Frendo, P., Frugier, F., Masson-Boivin, C., Eds.; Regulation of Nitrogen-Fixing Symbioses in Legumes; Academic Press: Cambridge, MA, USA, 2020; Volume 94, pp. 213–249. [Google Scholar]
- Smith, D.; Praslickova, D.; Ilangumaran, G. Inter-Organismal Signaling and Management of the Phytomicrobiome. Front. Plant Sci. 2015, 6, 722. [Google Scholar] [CrossRef] [Green Version]
- Leach, J.E.; Triplett, L.R.; Argueso, C.T.; Trivedi, P. Communication in the Phytobiome. Cell 2017, 169, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Zipfel, C. Plant Pattern-Recognition Receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef]
- Lareen, A.; Burton, F.; Schäfer, P. Plant Root-Microbe Communication in Shaping Root Microbiomes. Plant Mol. Biol. 2016, 90, 575–587. [Google Scholar] [CrossRef] [Green Version]
- Mhlongo, M.I.; Piater, L.A.; Madala, N.E.; Labuschagne, N.; Dubery, I.A. The Chemistry of Plant–Microbe Interactions in the Rhizosphere and the Potential for Metabolomics to Reveal Signaling Related to Defense Priming and Induced Systemic Resistance. Front. Plant Sci. 2018, 9, 112. [Google Scholar] [CrossRef] [Green Version]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the Root Microbiome by Plant Molecules: The Basis for Targeted Disease Suppression and Plant Growth Promotion. Front. Plant Sci. 2020, 10, 1741. [Google Scholar] [CrossRef]
- Kaschuk, G.; Leffelaar, P.A.; Giller, K.E.; Alberton, O.; Hungria, M.; Kuyper, T.W. Responses of Legumes to Rhizobia and Arbuscular Mycorrhizal Fungi: A Meta-Analysis of Potential Photosynthate Limitation of Symbioses. Soil Biol. Biochem. 2010, 42, 125–127. [Google Scholar] [CrossRef]
- Haichar, F.e.Z.; Santaella, C.; Heulin, T.; Achouak, W. Root Exudates Mediated Interactions Belowground. Soil Biol Biochem. 2014, 77, 69–80. [Google Scholar] [CrossRef]
- Ferguson, B.J.; Mens, C.; Hastwell, A.H.; Zhang, M.; Su, H.; Jones, C.H.; Chu, X.; Gresshoff, P.M. Legume Nodulation: The Host Controls the Party. Plant Cell Environ. 2019, 42, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venturi, V.; Keel, C. Signaling in the Rhizosphere. Trends Plant Sci. 2016, 21, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Haichar, F.e.Z.; Heulin, T.; Guyonnet, J.P.; Achouak, W. Stable Isotope Probing of Carbon Flow in the Plant Holobiont. Curr. Opin. Biotechnol. 2016, 41, 9–13. [Google Scholar] [CrossRef]
- Vives-Peris, V.; de Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Root Exudates: From Plant to Rhizosphere and Beyond. Plant Cell Rep. 2020, 39, 3–17. [Google Scholar] [CrossRef]
- Abedini, D.; Jaupitre, S.; Bouwmeester, H.; Dong, L. Metabolic Interactions in Beneficial Microbe Recruitment by Plants. Curr. Opin. Biotechnol. 2021, 70, 241–247. [Google Scholar] [CrossRef]
- Pereira, S.I.A.; Abreu, D.; Moreira, H.; Vega, A.; Castro, P.M.L. Plant Growth-Promoting Rhizobacteria (PGPR) Improve the Growth and Nutrient Use Efficiency in Maize (Zea mays L.) under Water Deficit Conditions. Heliyon 2020, 6, e05106. [Google Scholar] [CrossRef]
- Badri, D.V.; Chaparro, J.M.; Zhang, R.; Shen, Q.; Vivanco, J.M. Application of Natural Blends of Phytochemicals Derived from the Root Exudates of Arabidopsis to the Soil Reveal That Phenolic-Related Compounds Predominantly Modulate the Soil Microbiome. J. Biol. Chem. 2013, 288, 4502–4512. [Google Scholar] [CrossRef] [Green Version]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global Consequences of Land Use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [Green Version]
- Paungfoo-Lonhienne, C.; Redding, M.; Pratt, C.; Wang, W. Plant Growth Promoting Rhizobacteria Increase the Efficiency of Fertilisers While Reducing Nitrogen Loss. J. Environ. Manag. 2019, 233, 337–341. [Google Scholar] [CrossRef]
- Ke, X.; Feng, S.; Wang, J.; Lu, W.; Zhang, W.; Chen, M.; Lin, M. Effect of Inoculation with Nitrogen-Fixing Bacterium Pseudomonas stutzeri A1501 on Maize Plant Growth and the Microbiome Indigenous to the Rhizosphere. Syst. Appl. Microbiol. 2019, 42, 248–260. [Google Scholar] [CrossRef]
- Pii, Y.; Aldrighetti, A.; Valentinuzzi, F.; Mimmo, T.; Cesco, S. Azospirillum Brasilense Inoculation Counteracts the Induction of Nitrate Uptake in Maize Plants. J. Exp. Bot. 2019, 70, 1313–1324. [Google Scholar] [CrossRef] [PubMed]
- Kumawat, K.C.; Singh, I.; Nagpal, S.; Sharma, P.; Gupta, R.K.; Sirari, A. Co-Inoculation of Indigenous Pseudomonas oryzihabitans and Bradyrhizobium sp. Modulates the Growth, Symbiotic Efficacy, Nutrient Acquisition, and Grain Yield of Soybean. Pedosphere 2022, 32, 438–451. [Google Scholar] [CrossRef]
- Rather, S.A.; Hussain, M.A.; Sharma, N.L. Effect of Bio-Fertilizers on Growth, Yield and Economics of Field Pea (Pisum sativum L.). Int. J. Agric. Sci. 2010, 6, 65–66. [Google Scholar]
- Checcucci, A.; Marchetti, M. The Rhizosphere Talk Show: The Rhizobia on Stage. Front. Agron. 2020, 2, 591494. [Google Scholar] [CrossRef]
- Valdés-López, O.; Reyero-Saavedra, M.d.R.; Isidra-Arellano, M.C.; Sánchez-Correa, M.d.S. Early Molecular Dialogue Between Legumes and Rhizobia: Why Are They So Important? In Symbiosis: Cellular, Molecular, Medical and Evolutionary Aspects; Kloc, M., Ed.; Results and Problems in Cell Differentiation; Springer International Publishing: Cham, Switzerland, 2020; pp. 409–419. ISBN 978-3-030-51849-3. [Google Scholar]
- Dong, W.; Liu, E.; Yan, C.; Tian, J.; Zhang, H.; Zhang, Y. Impact of No Tillage vs. Conventional Tillage on the Soil Bacterial Community Structure in a Winter Wheat Cropping Succession in Northern China. Eur. J. Soil Biol. 2017, 80, 35–42. [Google Scholar] [CrossRef]
- Alahmad, A.; Decocq, G.; Spicher, F.; Kheirbeik, L.; Kobaissi, A.; Tetu, T.; Dubois, F.; Duclercq, J. Cover Crops in Arable Lands Increase Functional Complementarity and Redundancy of Bacterial Communities. J. Appl. Ecol. 2019, 56, 651–664. [Google Scholar] [CrossRef]
- Li, Y.; Jian, X.; Li, Y.; Zeng, X.; Xu, L.; Khan, M.U.; Lin, W. OsPAL2-1 Mediates Allelopathic Interactions between Rice and Specific Microorganisms in the Rhizosphere Ecosystem. Front. Microbiol. 2020, 11, 1411. [Google Scholar] [CrossRef]
- Peltoniemi, K.; Velmala, S.; Fritze, H.; Lemola, R.; Pennanen, T. Long-Term Impacts of Organic and Conventional Farming on the Soil Microbiome in Boreal Arable Soil. Eur. J. Soil Biol. 2021, 104, 103314. [Google Scholar] [CrossRef]
- Mazoyon, C.; Hirel, B.; Pecourt, A.; Catterou, M.; Gutierrez, L.; Sarazin, V.; Dubois, F.; Duclercq, J. Sphingomonas Sediminicola Is an Endosymbiotic Bacterium Able to Induce the Formation of Root Nodules in Pea (Pisum sativum L.) and to Enhance Plant Biomass Production. Microorganisms 2023, 11, 199. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Atanassova, M.; Georgieva, S.; Ivancheva, K. Total Phenolic and Total Flavonoid Contents, Antioxidant Capacity and Biological Contaminants in Medicinal Herbs. J. Univ. Chem. Technol. Metall. 2011, 46, 81–88. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Cawthray, G.R. An Improved Reversed-Phase Liquid Chromatographic Method for the Analysis of Low-Molecular Mass Organic Acids in Plant Root Exudates. J. Chromatogr. A 2003, 1011, 233–240. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for Large-Scale Metabolic Profiling of Serum and Plasma Using Gas Chromatography and Liquid Chromatography Coupled to Mass Spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef]
- Wen, B.; Mei, Z.; Zeng, C.; Liu, S. MetaX: A Flexible and Comprehensive Software for Processing Metabolomics Data. BMC Bioinform. 2017, 18, 183. [Google Scholar] [CrossRef] [Green Version]
- de Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research Version 1.3-5. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 13 February 2023).
- Hütsch, B.W.; Augustin, J.; Merbach, W. Plant Rhizodeposition—An Important Source for Carbon Turnover in Soils. J. Plant Nutr. Soil Sci. 2002, 165, 397–407. [Google Scholar] [CrossRef]
- Wang, Y.; Thorup-Kristensen, K.; Jensen, L.S.; Magid, J. Vigorous Root Growth Is a Better Indicator of Early Nutrient Uptake than Root Hair Traits in Spring Wheat Grown under Low Fertility. Front. Plant Sci. 2016, 7, 865. [Google Scholar] [CrossRef] [Green Version]
- Lv, X.; Wang, Q.; Zhang, X.; Hao, J.; Li, L.; Chen, W.; Li, H.; Wang, Y.; Ma, C.; Wang, J.; et al. Community Structure and Associated Networks of Endophytic Bacteria in Pea Roots throughout Plant Life Cycle. Plant Soil 2021, 468, 225–238. [Google Scholar] [CrossRef]
- Anzuay, M.S.; Viso, N.P.; Ludueña, L.M.; Morla, F.D.; Angelini, J.G.; Taurian, T. Plant Beneficial Rhizobacteria Community Structure Changes through Developmental Stages of Peanut and Maize. Rhizosphere 2021, 19, 100407. [Google Scholar] [CrossRef]
- Tian, T.; Sun, B.; Shi, H.; Gao, T.; He, Y.; Li, Y.; Liu, Y.; Li, X.; Zhang, L.; Li, S.; et al. Sucrose Triggers a Novel Signaling Cascade Promoting Bacillus Subtilis Rhizosphere Colonization. ISME J. 2021, 15, 2723–2737. [Google Scholar] [CrossRef]
- Knee, E.M.; Gong, F.-C.; Gao, M.; Teplitski, M.; Jones, A.R.; Foxworthy, A.; Mort, A.J.; Bauer, W.D. Root Mucilage from Pea and Its Utilization by Rhizosphere Bacteria as a Sole Carbon Source. MPMI 2001, 14, 775–784. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Feng, H.; Fu, R.; Zhang, N.; Du, W.; Shen, Q.; Zhang, R. Induced Root-Secreted d-Galactose Functions as a Chemoattractant and Enhances the Biofilm Formation of Bacillus Velezensis SQR9 in an McpA-Dependent Manner. Appl. Microbiol. Biotechnol. 2020, 104, 785–797. [Google Scholar] [CrossRef]
- Gunina, A.; Kuzyakov, Y. Sugars in Soil and Sweets for Microorganisms: Review of Origin, Content, Composition and Fate. Soil Biol. Biochem. 2015, 90, 87–100. [Google Scholar] [CrossRef]
- Souza, W.R.d. Microbial Degradation of Lignocellulosic Biomass; IntechOpen: London, UK, 2013; ISBN 978-953-51-1119-1. [Google Scholar]
- Honvault, N.; Houben, D.; Firmin, S.; Meglouli, H.; Laruelle, F.; Fontaine, J.; Lounès-Hadj Sahraoui, A.; Coutu, A.; Lambers, H.; Faucon, M.-P. Interactions between Below-Ground Traits and Rhizosheath Fungal and Bacterial Communities for Phosphorus Acquisition. Funct. Ecol. 2021, 35, 1603–1619. [Google Scholar] [CrossRef]
- Lambers, H.; Hayes, P.E.; Laliberté, E.; Oliveira, R.S.; Turner, B.L. Leaf Manganese Accumulation and Phosphorus-Acquisition Efficiency. Trends Plant Sci. 2015, 20, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sindhu, S.S.; Sehrawat, A.; Glick, B.R. The Involvement of Organic Acids in Soil Fertility, Plant Health and Environment Sustainability. Arch. Microbiol. 2022, 204, 720. [Google Scholar] [CrossRef]
- Ling, N.; Raza, W.; Ma, J.; Huang, Q.; Shen, Q. Identification and Role of Organic Acids in Watermelon Root Exudates for Recruiting Paenibacillus polymyxa SQR-21 in the Rhizosphere. Eur. J. Soil Biol. 2011, 47, 374–379. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, S.; Chai, Y.; Clardy, J.; Kolter, R.; Guo, J.; Losick, R. A Bacillus Subtilis Sensor Kinase Involved in Triggering Biofilm Formation on the Roots of Tomato Plants. Mol. Microbiol. 2012, 85, 418–430. [Google Scholar] [CrossRef] [Green Version]
- Vora, S.M.; Joshi, P.; Belwalkar, M.; Archana, G. Root Exudates Influence Chemotaxis and Colonization of Diverse Plant Growth Promoting Rhizobacteria in the Pigeon Pea—Maize Intercropping System. Rhizosphere 2021, 18, 100331. [Google Scholar] [CrossRef]
- Raffi, M.M.; Charyulu, P.B.B.N. Chapter 18—Azospirillum-Biofertilizer for Sustainable Cereal Crop Production: Current Status. In Recent Developments in Applied Microbiology and Biochemistry; Viswanath, B., Ed.; Academic Press: Cambridge, MA, USA, 2021; pp. 193–209. ISBN 978-0-12-821406-0. [Google Scholar]
- Archana, G.; Buch, A.; Kumar, G.N. Pivotal Role of Organic Acid Secretion by Rhizobacteria in Plant Growth Promotion. In Microorganisms in Sustainable Agriculture and Biotechnology; Satyanarayana, T., Johri, B.N., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 35–53. ISBN 978-94-007-2214-9. [Google Scholar]
- Panichikkal, J.; Krishnankutty, R.E. Root Exudate Components Induced Production of Plant Beneficial Metabolites in Rhizospheric Pseudomonas Spp. Rhizosphere 2021, 19, 100366. [Google Scholar] [CrossRef]
- Israr, D.; Mustafa, G.; Khan, K.S.; Shahzad, M.; Ahmad, N.; Masood, S. Interactive Effects of Phosphorus and Pseudomonas putida on Chickpea (Cicer arietinum L.) Growth, Nutrient Uptake, Antioxidant Enzymes and Organic Acids Exudation. Plant Physiol. Biochem. 2016, 108, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Mukerji, K.G. Root Exudates as Determinant of Rhizospheric Microbial Biodiversity. In Microbial Activity in the Rhizoshere; Mukerji, K.G., Manoharachary, C., Singh, J., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2006; pp. 39–53. ISBN 978-3-540-29420-7. [Google Scholar]
- Macias-Benitez, S.; Garcia-Martinez, A.M.; Caballero Jimenez, P.; Gonzalez, J.M.; Tejada Moral, M.; Parrado Rubio, J. Rhizospheric Organic Acids as Biostimulants: Monitoring Feedbacks on Soil Microorganisms and Biochemical Properties. Front. Plant Sci. 2020, 11, 633. [Google Scholar] [CrossRef]
- Wang, H.-W.; Ma, C.-Y.; Xu, F.-J.; Lu, F.; Zhang, W.; Dai, C.-C. Root Endophyte-Enhanced Peanut-Rhizobia Interaction Is Associated with Regulation of Root Exudates. Microbiol. Res. 2021, 250, 126765. [Google Scholar] [CrossRef]
- Ghasemi, A.; Moosavi-Nasab, M.; Setoodeh, P.; Mesbahi, G.; Yousefi, G. Biosurfactant Production by Lactic Acid Bacterium Pediococcus dextrinicus SHU1593 Grown on Different Carbon Sources: Strain Screening Followed by Product Characterization. Sci. Rep. 2019, 9, 5287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raman, J.; Kim, J.-S.; Choi, K.R.; Eun, H.; Yang, D.; Ko, Y.-J.; Kim, S.-J. Application of Lactic Acid Bacteria (LAB) in Sustainable Agriculture: Advantages and Limitations. Int. J. Mol. Sci. 2022, 23, 7784. [Google Scholar] [CrossRef]
- Saranraj, P.; Naidu, M.A.; Sivasakthivelan, P. Lactic Acid Bacteria and Its Antimicrobial Properties: A Review. Int. J. Pharm. Biol. Arch. 2013, 4, 1124–1133. [Google Scholar]
- Naik, K.; Mishra, S.; Srichandan, H.; Singh, P.K.; Sarangi, P.K. Plant Growth Promoting Microbes: Potential Link to Sustainable Agriculture and Environment. Biocatal. Agric. Biotechnol. 2019, 21, 101326. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, D.; Liu, Y.; Li, S.; Shen, Q.; Zhang, R. Effects of Different Plant Root Exudates and Their Organic Acid Components on Chemotaxis, Biofilm Formation and Colonization by Beneficial Rhizosphere-Associated Bacterial Strains. Plant Soil 2014, 374, 689–700. [Google Scholar] [CrossRef]
- Yuan, J.; Zhang, N.; Huang, Q.; Raza, W.; Li, R.; Vivanco, J.M.; Shen, Q. Organic Acids from Root Exudates of Banana Help Root Colonization of PGPR Strain Bacillus amyloliquefaciens NJN-6. Sci. Rep. 2015, 5, 13438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Liu, G.; Li, Y.; Zhu, S.; Zou, A.; Qi, J.; Yang, Y. Rhizosphere Microbial Communities and Organic Acids Secreted by Aluminum-Tolerant and Aluminum-Sensitive Soybean in Acid Soil. Biol. Fertil. Soils 2012, 48, 97–108. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Badri, D.V.; Bakker, M.G.; Sugiyama, A.; Manter, D.K.; Vivanco, J.M. Root Exudation of Phytochemicals in Arabidopsis Follows Specific Patterns that Are Developmentally Programmed and Correlate with Soil Microbial Functions. PLoS ONE 2013, 8, e55731. [Google Scholar] [CrossRef]
- Canarini, A.; Kaiser, C.; Merchant, A.; Richter, A.; Wanek, W. Root Exudation of Primary Metabolites: Mechanisms and Their Roles in Plant Responses to Environmental Stimuli. Front. Plant Sci. 2019, 10, 157. [Google Scholar] [CrossRef] [Green Version]
- Smirnova, I.; Sadanov, A.; Baimakhanova, G.; Faizulina, E.; Tatarkina, L. Metabolic Interaction at the Level of Extracellular Amino Acids between Plant Growth-Promoting Rhizobacteria and Plants of Alfalfa (Medicago sativa L.). Rhizosphere 2022, 21, 100477. [Google Scholar] [CrossRef]
- Shaw, L.J.; Morris, P.; Hooker, J.E. Perception and Modification of Plant Flavonoid Signals by Rhizosphere Microorganisms. Environ. Microbiol. 2006, 8, 1867–1880. [Google Scholar] [CrossRef]
- Singh, U.P.; Sarma, B.K.; Singh, D.P. Effect of Plant Growth-Promoting Rhizobacteria and Culture Filtrate of Sclerotium rolfsii on Phenolic and Salicylic Acid Contents in Chickpea (Cicer arietinum). Curr. Microbiol. 2003, 46, 131–140. [Google Scholar] [CrossRef]
- Feng, H.; Zhang, N.; Du, W.; Zhang, H.; Liu, Y.; Fu, R.; Shao, J.; Zhang, G.; Shen, Q.; Zhang, R. Identification of Chemotaxis Compounds in Root Exudates and Their Sensing Chemoreceptors in Plant-Growth-Promoting Rhizobacteria Bacillus amyloliquefaciens SQR9. MPMI 2018, 31, 995–1005. [Google Scholar] [CrossRef] [Green Version]
- Bag, S.; Mondal, A.; Majumder, A.; Mondal, S.K.; Banik, A. Flavonoid Mediated Selective Cross-Talk between Plants and Beneficial Soil Microbiome. Phytochem. Rev. 2022, 21, 1739–1760. [Google Scholar] [CrossRef]
- Shah, A.; Smith, D.L. Flavonoids in Agriculture: Chemistry and Roles in, Biotic and Abiotic Stress Responses, and Microbial Associations. Agronomy 2020, 10, 1209. [Google Scholar] [CrossRef]
- Singh, K.; Chandra, R.; Purchase, D. Unraveling the Secrets of Rhizobacteria Signaling in Rhizosphere. Rhizosphere 2022, 21, 100484. [Google Scholar] [CrossRef]
- Cannesan, M.A.; Gangneux, C.; Lanoue, A.; Giron, D.; Laval, K.; Hawes, M.; Driouich, A.; Vicré-Gibouin, M. Association between Border Cell Responses and Localized Root Infection by Pathogenic Aphanomyces euteiches. Ann. Bot. 2011, 108, 459–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selim, S.; Sanssené, J.; Rossard, S.; Courtois, J. Systemic Induction of the Defensin and Phytoalexin Pisatin Pathways in Pea (Pisum sativum) against Aphanomyces euteiches by Acetylated and Nonacetylated Oligogalacturonides. Molecules 2017, 22, 1017. [Google Scholar] [CrossRef] [Green Version]
- Olanrewaju, O.S.; Ayangbenro, A.S.; Glick, B.R.; Babalola, O.O. Plant Health: Feedback Effect of Root Exudates-Rhizobiome Interactions. Appl. Microbiol. Biotechnol. 2019, 103, 1155–1166. [Google Scholar] [CrossRef] [Green Version]
- Turetschek, R.; Desalegn, G.; Epple, T.; Kaul, H.-P.; Wienkoop, S. Key Metabolic Traits of Pisum sativum Maintain Cell Vitality during Didymella pinodes Infection: Cultivar Resistance and the Microsymbionts’ Influence. J. Proteom. 2017, 169, 189–201. [Google Scholar] [CrossRef]
- Novák, K.; Lisá, L.; Škrdleta, V. Rhizobial Nod Gene-Inducing Activity in Pea Nodulation Mutants: Dissociation of Nodulation and Flavonoid Response. Physiol. Plant. 2004, 120, 546–555. [Google Scholar] [CrossRef]
- Bringhurst, R.M.; Cardon, Z.G.; Gage, D.J. Galactosides in the Rhizosphere: Utilization by Sinorhizobium meliloti and Development of a Biosensor. Proc. Natl. Acad. Sci. 2001, 98, 4540–4545. [Google Scholar] [CrossRef]
- Xie, Z.-P.; Staehelin, C.; Broughton, W.J.; Wiemken, A.; Boller, T.; Müller, J. Accumulation of Soluble Carbohydrates, Trehalase and Sucrose Synthase in Effective (Fix+) and Ineffective (Fix–) Nodules of Soybean Cultivars that Differentially Nodulate with Bradyrhizobium japonicum. Funct. Plant Biol. 2003, 30, 965–971. [Google Scholar] [CrossRef]
- Palacios, O.A.; Espinoza-Hicks, J.C.; Camacho-Dávila, A.A.; López, B.R.; de-Bashan, L.E. Differences in Exudates between Strains of Chlorella sorokiniana Affect the Interaction with the Microalga Growth-Promoting Bacteria Azospirillum brasilense. Microb. Ecol. 2022, 85, 1412–1422. [Google Scholar] [CrossRef]
- Poole, P.S.; Ledermann, R. Maintaining Osmotic Balance in Legume Nodules. J. Exp. Bot. 2022, 73, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Hickey, A.; Pardo, L.M.; Reen, F.J.; McGlacken, G.P. Pyrones Identified as LuxR Signal Molecules in Photorhabdus and Their Synthetic Analogues Can Alter Multicellular Phenotypic Behavior of Bacillus atropheaus. ACS Omega 2021, 6, 33141–33148. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, A.L.; Greenberg, E.P.; Oliver, C.M.; Oda, Y.; Huang, J.J.; Bittan-Banin, G.; Peres, C.M.; Schmidt, S.; Juhaszova, K.; Sufrin, J.R.; et al. A New Class of Homoserine Lactone Quorum-Sensing Signals. Nature 2008, 454, 595–599. [Google Scholar] [CrossRef]
- Lynch, D.V.; Dunn, T.M. An Introduction to Plant Sphingolipids and a Review of Recent Advances in Understanding Their Metabolism and Function. New Phytol. 2004, 161, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Michaelson, L.V.; Napier, J.A.; Molino, D.; Faure, J.-D. Plant Sphingolipids: Their Importance in Cellular Organization and Adaption. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 1329–1335. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J. Auxin and Plant-Microbe Interactions. Cold Spring Harb. Perspect. Biol. 2011, 3, a001438. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazoyon, C.; Firmin, S.; Bensaddek, L.; Pecourt, A.; Chabot, A.; Faucon, M.-P.; Sarazin, V.; Dubois, F.; Duclercq, J. Optimizing Crop Production with Bacterial Inputs: Insights into Chemical Dialogue between Sphingomonas sediminicola and Pisum sativum. Microorganisms 2023, 11, 1847. https://doi.org/10.3390/microorganisms11071847
Mazoyon C, Firmin S, Bensaddek L, Pecourt A, Chabot A, Faucon M-P, Sarazin V, Dubois F, Duclercq J. Optimizing Crop Production with Bacterial Inputs: Insights into Chemical Dialogue between Sphingomonas sediminicola and Pisum sativum. Microorganisms. 2023; 11(7):1847. https://doi.org/10.3390/microorganisms11071847
Chicago/Turabian StyleMazoyon, Candice, Stéphane Firmin, Lamine Bensaddek, Audrey Pecourt, Amélie Chabot, Michel-Pierre Faucon, Vivien Sarazin, Fréderic Dubois, and Jérôme Duclercq. 2023. "Optimizing Crop Production with Bacterial Inputs: Insights into Chemical Dialogue between Sphingomonas sediminicola and Pisum sativum" Microorganisms 11, no. 7: 1847. https://doi.org/10.3390/microorganisms11071847