
MALDI-TOF MS: A Quick Method to Detect the Susceptibility of Fusarium spp. Clinical Isolates to Amphotericin B
 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Isolates and Strains
2.2. Broth Microdilution Susceptibility Method and the Minimal Inhibitory Concentration
2.3. MALDI-TOF MS Spectra Acquisition
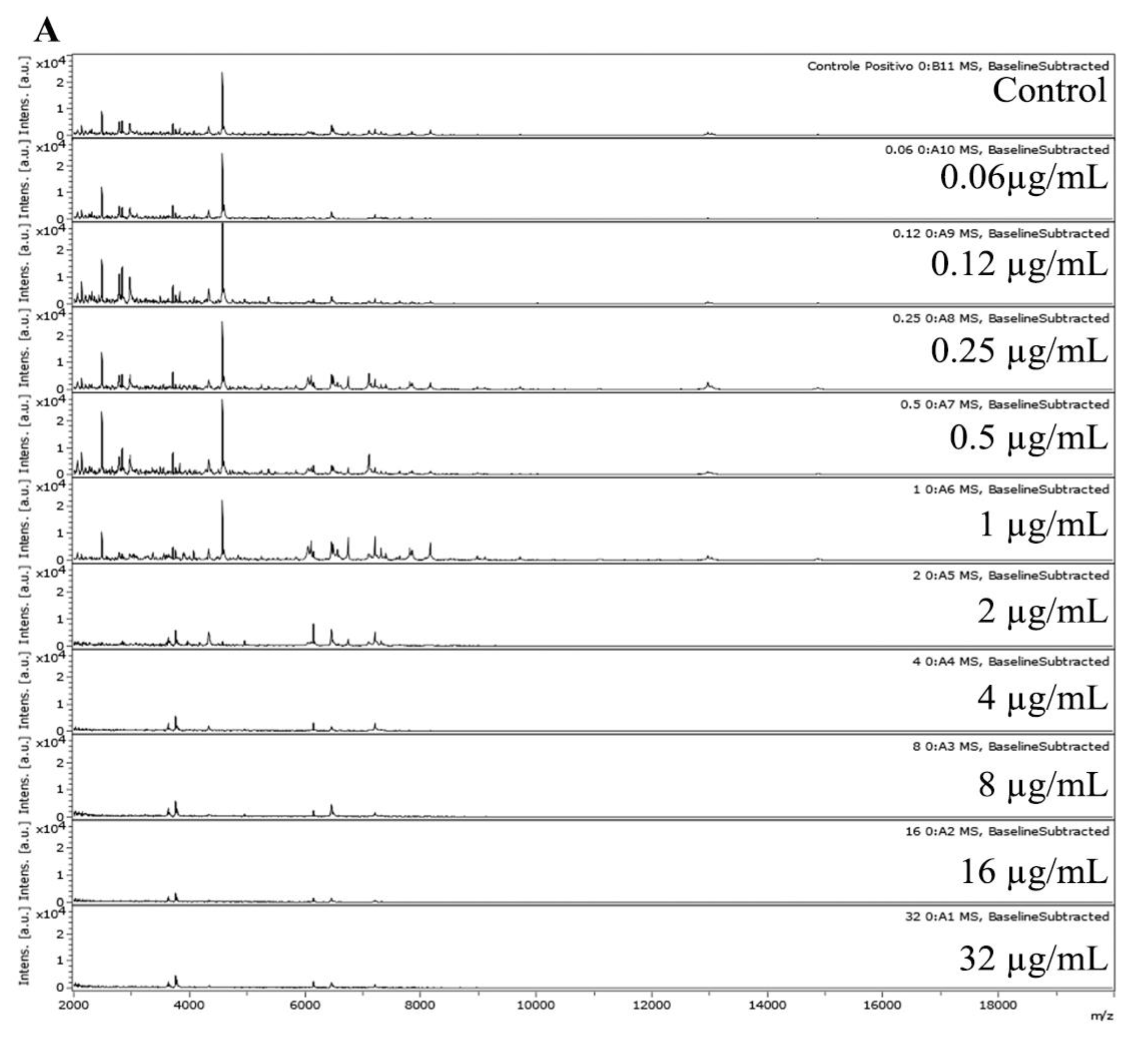
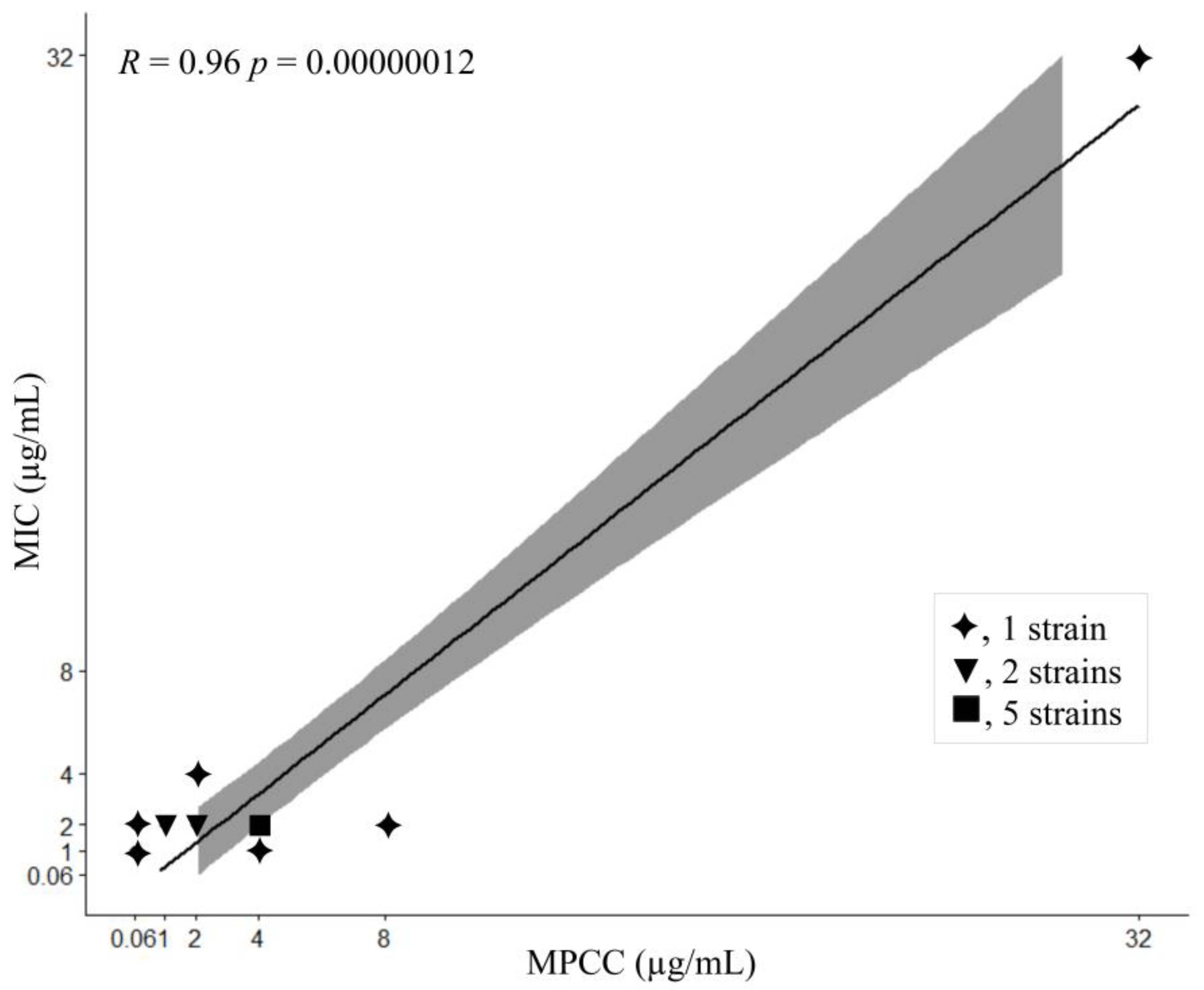
2.4. Data Analysis and MPCC Determination
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Lombard, L.; Sandoval-Denis, M.; Cai, L.; Crous, P.W. Changing the game: Resolving systematic issues in key Fusarium species complexes. Persoonia 2019, 43, i–ii. [Google Scholar] [CrossRef] [PubMed]
- Lainhart, W. Fusarium spp., a Genus of Common Plant Pathogens That Can Cause Devastating, Opportunistic Human Disease. Clin. Microbiol. Newsl. 2017, 40, 1–5. [Google Scholar] [CrossRef]
- Herkert, P.F.; Al-Hatmi, A.M.S.; de Oliveira Salvador, G.L.; Muro, M.D.; Pinheiro, R.L.; Nucci, M.; Queiroz-Telles, F.; De Hoog, G.S.; Meis, J.F. Molecular characterization and antifungal susceptibility of clinical Fusarium species from Brazil. Front. Microbiol. 2019, 10, 737. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, G.; Keller, N. Crossover fungal pathogens: The biology andpathogenesis of fungi capable of crossing kingdoms to infect plants and humans. Fung. Genet. Biol. 2013, 61, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Van Diepeningen, A.D.; De Hoog, G.S. Challenges in Fusarium, a Trans-Kingdom Pathogen. Mycopathologia 2016, 181, 161–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Summerell, B.A. Resolving Fusarium: Current status of the genus. Annu. Rev. Phytopathol. 2019, 57, 323–339. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J.; Bennett, J.E.; Wickes, B.L.; Meyer, W.; Cuomo, C.A.; Wollenburg, K.R.; Bicanic, T.A.; Castañeda, E.; Chang, Y.C.; Chen, J.; et al. The case for adopting the “species complex” nomenclature for the etiologic agents of cryptococcosis. mSphere 2017, 2, e00357-16. [Google Scholar] [CrossRef] [Green Version]
- Geiser, D.M.; Al-Hatmi, A.; Aoki, T.; Arie, T.; Balmas, V.; Barnes, I.; Bergstrom, G.C.; Bhattacharyya, M.K.K.; Blomquist, C.L.; Bowden, R.; et al. Phylogenomic analysis of a 55.1 kb 19-gene dataset resolves a monophyletic Fusarium that includes the Fusarium solani species complex. Phytopathology 2021, 111, 1064–1079. [Google Scholar] [CrossRef]
- Al-Hatmi, A.M.; Ende, A.G.V.D.; Stielow, J.B.; Van Diepeningen, A.D.; Seifert, K.A.; McCormick, W.; Assabgui, R.; Gräfenhan, T.; De Hoog, G.S.; Levesque, C.A. Evaluation of two novel barcodes for species recognition of opportunistic pathogens in Fusarium. Fung. Biol. 2016, 120, 231–245. [Google Scholar] [CrossRef]
- Batista, M.; Costa, S.; Shikanai-Yasuda, M.; Moss, R. Current treatment options for invasive aspergillosis. Drugs Today 2013, 3, 213–226. [Google Scholar] [CrossRef]
- Triest, D.; Stubbe, D.; De Cremer, K.; Piérard, D.; Normand, A.-C.; Piarroux, R.; Detandt, M.; Hendrickx, M. Use of matrix-assisted laser desorption ionization-time of flight mass spectrometry for identification of molds of the Fusarium genus. J. Clin. Microbiol. 2015, 53, 465–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.X.; Zheng, L.; Hsiang, T.; Yang, G.L.; Zhao, D.L.; Lv, B.; Chen, Y.F.; Huang, J.B. Detection and quantification of Fusarium commune in host tissue and infested soil using real-time PCR. Plant. Pathology 2016, 65, 218–226. [Google Scholar] [CrossRef]
- Tortorano, A.M.; Richardson, M.; Roilides, E.; van Diepeningen, A.; Caira, M.; Munoz, P.; Johnson, E.; Meletiadis, J.; Pana, Z.-D.; Lackner, M.; et al. ESCMID and ECMM joint guidelines on diagnosis and management of hyalohyphomycosis: Fusarium spp., Scedosporium spp. and others. Clin. Microbiol. Infect. 2014, 20, 27–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harpaz, R.; Dahl, R.; Dooling, K. The prevalence of immunocompromised adults: United States, 2013. Open Forum Infect. Dis. 2016, 3 (Suppl. S1), 1439. [Google Scholar] [CrossRef]
- Lass-Florl, C.; Cuenca-Estrella, M. Changes in the epidemiological landscape of invasive mould infections and disease. J. Antimicrob. Chemother. 2017, 72 (Suppl. S1), i5–i11. [Google Scholar] [CrossRef]
- Smith, K.D.; Achan, B.; Hullsiek, K.H.; McDonald, T.R.; Okagaki, L.H.; Alhadab, A.A.; Akampurira, A.; Rhein, J.R.; Meya, D.B.; Boulware, D.R.; et al. Increased antifungal drug resistance in clinical isolates of Cryptococcus neoformans in Uganda. Antimicrob. Agents Chemother. 2015, 59, 7197–7204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, M.W.; Kontoyiannis, D.P.; Cornely, O.A.; Perfect, J.R.; Walsh, T.J. Novel Agents and Drug Targets to Meet the Challenges of Resistant Fungi. J. Infect. Dis. 2017, 216, S474–S483. [Google Scholar] [CrossRef] [Green Version]
- Sikora, A.; Hashmi, M.F.; Zahra, F. Candida Auris. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK563297/ (accessed on 27 April 2023).
- Lucas, J.A.; Hawkins, N.J.; Fraaije, B.A. The evolution of fungicide resistance. Adv. Appl. Microbiol. 2015, 90, 29–92. [Google Scholar] [CrossRef]
- Al-Hatmi, A.M.S.; Meis, J.F.; De Hoog, G.S. Fusarium: Molecular diversity and intrinsic drug resistance. PLoS Pathog. 2016, 4, e1005464. [Google Scholar] [CrossRef]
- Ray, S.; Das, S.; Suar, M. Molecular Mechanism of Drug Resistance. In Drug Resistance in Bacteria, Fungi, Malaria, and Cancer, 1st ed.; Arora, G., Sajid, A., Kalia, V.C., Eds.; Springer International Publishing: Cham, Switzerland, 2017; Volume 22, pp. 47–110. [Google Scholar] [CrossRef]
- Lagudah, E.S.; Krattinger, S.G. A new player contributing to durable Fusarium resistance. Nat. Genet. 2019, 51, 1070–1071. [Google Scholar] [CrossRef]
- Blaize, M.; Normand, A.-C.; Imbert, S.; Al-Hatmi, A.M.S.; Chryssanthou, E.; Cassaing, S.; Schuttler, C.; Hasseine, L.; Mahinc, C.; Costa, D.; et al. Antifungal susceptibility of 182 Fusarium species isolates from 20 European centers: Comparison between EUCAST and gradient concentration strip methods. Antimicrob. Agents Chemother. 2021, 65, e0149521. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Sutton, D.A.; Fothergill, A.; McCarthy, D.; Rinaldi, M.G.; Brandt, M.E.; Zhang, N.; Geiser, D.M. Molecular phylogenetic diversity, multilocus haplotype nomenclature, and in vitro antifungal resistance within the Fusarium solani species complex. J. Clin. Microbiol. 2008, 46, 2477–2490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Hatmi, A.M.S.; Van Diepeningen, A.D.; Curfs-Breuker, I.; De Hoog, G.S.; Meis, J.F. Specific antifungal susceptibility profiles of opportunists in the Fusarium fujikuroi complex. J. Antimicrob. Chemother. 2015, 70, 1068–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Liu, X.; Yang, Z.; Meng, X.; Xue, R.; Yu, J.; Al-Hatmi, A.M.; de Hoog, G.S.; Li, R. Molecular and MALDI-TOF MS differentiation and antifungal susceptibility of prevalent clinical Fusarium species in China. Mycoses 2021, 64, 1261–1271. [Google Scholar] [CrossRef]
- Anaissie, E.; Nelson, P.; Beremand, M.; Kontoyiannis, D.; Rinaldi, M. Fusarium caused hyalohyphomycosis. An: Overview. Curr. Top. Med. Mycol. 1992, 4, 231–249. [Google Scholar]
- Sutton, D.A.; Brandt, M.E. Fusarium and other opportunistic hyaline fungi. In Manual of Clinical Microbiology, 10th ed.; ASM Press: Washington, DC, USA, 2011; pp. 1853–1879. [Google Scholar]
- Moretti, M.L.; Busso-Lopes, A.; Moraes, R.; Muraosa, Y.; Mikami, Y.; Trabasso, P.; Tominaga, K.; Reichert-Lima, F.; Lyra, L.; Gonoi, T.; et al. Environment as a potential source of Fusarium spp. invasive infections in immunocompromised patients. Open Forum Infect. Dis. 2014, 1, S38. [Google Scholar] [CrossRef]
- Garnica, M.; Da Cunha, M.O.; Portugal, R.; Maiolino, A.; Colombo, A.L.; Nucci, M. Risk factors for invasive fusariosis in patients with acute myeloid leukemia and in hematopoietic cell transplant recipients. Clin. Infect. Dis. 2015, 60, 875–880. [Google Scholar] [CrossRef] [Green Version]
- Rotjanapan, P.; Chen, Y.C.; Chakrabarti, A.; Li, R.Y.; Rudramurthy, S.; Yu, J.; Kung, H.C.; Watcharananan, S.; Tan, A.L.; Saffari, S.E.; et al. Epidemiology and clinical characteristics of invasive mould infections: A multicenter, retrospective analysis in five Asian countries. Med. Mycol. 2018, 56, 186–196. [Google Scholar] [CrossRef] [Green Version]
- WHO. Fungal Priority Pathogens List to Guide Research, Development and Public Health Action; World Health Organization: Geneva, Switzerland, 2022; Licence: CC BY-NC-SA 3.0 IGO. [Google Scholar]
- Approved Standard 2008, M38-A2, 3rd; Reference Methods for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi. Clinical and Laboratory Standartd Institute: Wayne, PA, USA, 2008.
- EUCAST. The European Committee on Antimicrobial Susceptibility Testing: Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for conidia forming moulds. In EUCAST Antifungal MIC Method for Moulds; EUCAST: Växjö, Sweden, 2020; DOCUMENT E.DEF 9.3.2. [Google Scholar]
- Espinel-Ingroff, A.; Colombo, A.L.; Cordoba, S.; Dufresne, P.J.; Fuller, J.; Ghannoum, M.; Gonzalez, G.M.; Guarro, J.; Kidd, S.E.; Meis, J.F.; et al. An international evaluation of MIC distributions and ECV definition for Fusarium species identified by molecular methods for the CLSI broth microdilution method. Antimicrob. Agents Chemother. 2016, 60, 1079–1084. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Velásquez, J.C.; Mojica-Figueroa, I.L.; Santos, C.; Lima, N.; Mesa-Arango, A.C. MALDI-TOF MS: Foundations and a practical approach to the clinical relevant filamentous fungi identification. Curr. Fungal Infect. Rep. 2021, 15, 162–170. [Google Scholar] [CrossRef]
- Pereira, L.; Dias, N.; Santos, C.; Lima, N. The use of MALDI-TOF ICMS as an alternative tool for Trichophyton rubrum identification and typing. Enfermedades Infecc. Microbiol. Clín. 2014, 32, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Rabello, V.B.d.S.; Corrêa-Moreira, D.; Santos, C.; Pinto, T.C.A.; Procopio-Azevedo, A.C.; Boechat, J.; Coelho, R.A.; Almeida-Paes, R.; Costa, G.; Lima, N.; et al. Preservation Methods in Isolates of Sporothrix Characterized by Polyphasic Approach. J. Fungi 2023, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- Marinach, C.; Alanio, A.; Palous, M.; Fekkar, A.; Brossas, J.-Y.; Brun, S.; Snounou, G.; Hennequin, C.; Sanglard, D.; Datry, A.; et al. MALDI-TOF MS-based drug susceptibility testing of pathogens: The example of Candida albicans and fluconazole. Proteomics 2009, 9, 4627–4631. [Google Scholar] [CrossRef]
- De Carolis, E.; Vella, A.; Florio, A.R.; Posteraro, P.; Perlin, D.S.; Sanguinetti, M.; Posteraro, B. Use of matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDITOF MS) for caspofungin susceptibility testing of Candida and Aspergillus species. J. Clin. Microbiol. 2012, 50, 2479–2483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberto, A.E.M.; Xavier, D.E.; Vidal, E.E.; Vidal, C.F.d.L.; Neves, R.P.; de Lima-Neto, R.G. Rapid Detection of Echinocandins Resistance by MALDI-TOF MS in Candida parapsilosis Complex. Microorganisms 2020, 8, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vella, A.; De Carolis, E.; Vaccaro, L.; Posteraro, P.; Perlin, D.S.; Kostrzewa, M.; Posteraro, B.; Sanguinetti, M. Rapid antifungal susceptibility testing by Matrix-Assisted Laser Desorption Ionization Time-of-Flight Mass Spectrometry analysis. J. Clin. Microbiol. 2013, 51, 2964–2969. [Google Scholar] [CrossRef] [Green Version]
- Saracli, M.A.; Fothergill, A.W.; Sutton, D.A.; Wiederhold, N.P. Detection of triazole resistance among Candida species by matrix-assisted laser desorption/ionization-time of flight mass spectrometry (MALDI-TOF MS). Med. Mycol. 2015, 53, 736–742. [Google Scholar] [CrossRef] [Green Version]
- Vella, A.; De Carolis, E.; Mello, E.; Perlin, D.S.; Sanglard, D.; Sanguinetti, M.; Posteraro, B. Potential use of MALDI-TOF mass spectrometry for rapid detection of antifungal resistance in the human pathogen Candida glabrata. Sci. Rep. 2017, 7, 9099. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.; Singh, P.; AS, S.; Rudramurthy, S.M.; Chakrabarti, A.; Ghosh, A.K. Rapid detection of fluconazole resistance in Candida tropicalis by MALDI-TOF MS. Med. Mycol. 2018, 56, 234–241. [Google Scholar] [CrossRef]
- Knoll, M.A.; Ulmer, H.; Lass-Flörl, C. Rapid Antifungal Susceptibility Testing of Yeasts and Molds by MALDI-TOF MS: A Systematic Review and Meta-Analysis. J. Fungi 2021, 7, 63. [Google Scholar] [CrossRef]
- Paziani, M.H.; Carvalho, L.T.; Melhem, M.D.S.C.; de Almeida, M.T.G.; da Silva, M.E.N.B.; Martinez, R.; Santos, C.; Kress, M.R.V.Z. First Comprehensive Report of Clinical Fusarium Strains Isolated in the State of Sao Paulo (Brazil) and Identified by MALDI-TOF MS and Molecular Biology. Microorganisms 2020, 8, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand, C.; Maubon, D.; Cornet, M.; Wang, Y.; Aldebert, D.; Garnaud, C. Can We Improve Antifungal Susceptibility Testing? Front. Cell. Infect. Microbiol. 2021, 11, 720609. [Google Scholar] [CrossRef] [PubMed]
- Araújo, E.; Gusmão, N.; Silva, T.; Le Pape, P.; Lima-Neto, R.G. MALDI-TOF MS-based evaluation for azole-susceptibility testing of Aspergillus fumigatus over reference broth microdilution method. Res. Sq. 2022; preprint. [Google Scholar] [CrossRef]
- Faustino, C.; Pinheiro, L. Lipid Systems for the Delivery of Amphotericin B in Antifungal Therapy. Pharmaceutics 2020, 12, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Complex | Species | Strain | MIC (µg/mL) | MPCC (µg/mL) |
|---|---|---|---|---|
| FOSC | F. oxysporum | ATCC48112 | 2 | 4 |
| LMC7137.01 | 2 | 4 | ||
| LMC7170.01 | 2 | 2 | ||
| FSSC | F. keratoplasticum | ATCC36031 | 2 | 4 |
| LMC7108.01 | >32 * | >32 * | ||
| LMC7113.02 | 2 | 1 | ||
| LMC7163.01 | 1 | <0.06 | ||
| LMC7205.01 | 4 | 2 | ||
| F. falciforme | LMC7178.01 | 2 | <0.06 | |
| LMC7193.01 | 2 | 8 | ||
| F. petroliphilum | LMC7162.02 | 2 | 4 | |
| FFSC | F. proliferatum | LMC7138.01 | 2 | 1 |
| LMC21300.01 | 2 | 4 | ||
| F. sacchari | LMC21303.01 | 2 | 2 | |
| FDSC | F. delphinoides | LMC7215.01 | 1 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grizante Barião, P.H.; Cayún, Y.; Sepúlveda, M.; Tonani, L.; Gonçalves de Almeida, O.G.; Cornejo, P.; Dias, N.; Santos, C.; von Zeska Kress, M.R. MALDI-TOF MS: A Quick Method to Detect the Susceptibility of Fusarium spp. Clinical Isolates to Amphotericin B. Microorganisms 2023, 11, 1834. https://doi.org/10.3390/microorganisms11071834
Grizante Barião PH, Cayún Y, Sepúlveda M, Tonani L, Gonçalves de Almeida OG, Cornejo P, Dias N, Santos C, von Zeska Kress MR. MALDI-TOF MS: A Quick Method to Detect the Susceptibility of Fusarium spp. Clinical Isolates to Amphotericin B. Microorganisms. 2023; 11(7):1834. https://doi.org/10.3390/microorganisms11071834
Chicago/Turabian StyleGrizante Barião, Patrícia Helena, Yasna Cayún, Marcela Sepúlveda, Ludmilla Tonani, Otavio Guilherme Gonçalves de Almeida, Pablo Cornejo, Nathalia Dias, Cledir Santos, and Marcia Regina von Zeska Kress. 2023. "MALDI-TOF MS: A Quick Method to Detect the Susceptibility of Fusarium spp. Clinical Isolates to Amphotericin B" Microorganisms 11, no. 7: 1834. https://doi.org/10.3390/microorganisms11071834