

Molecular Characterization of Clinical Linezolid-Resistant Staphylococcus epidermidis in a Tertiary Care Hospital

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Setting and Bacterial Isolates
2.2. Microbiological Methods
2.3. Whole-Genome Sequencing and Data Analysis
2.4. Statistical Analysis
3. Results
3.1. Patients′ Characteristics
3.2. Antimicrobial Susceptibility of LRSE Isolates
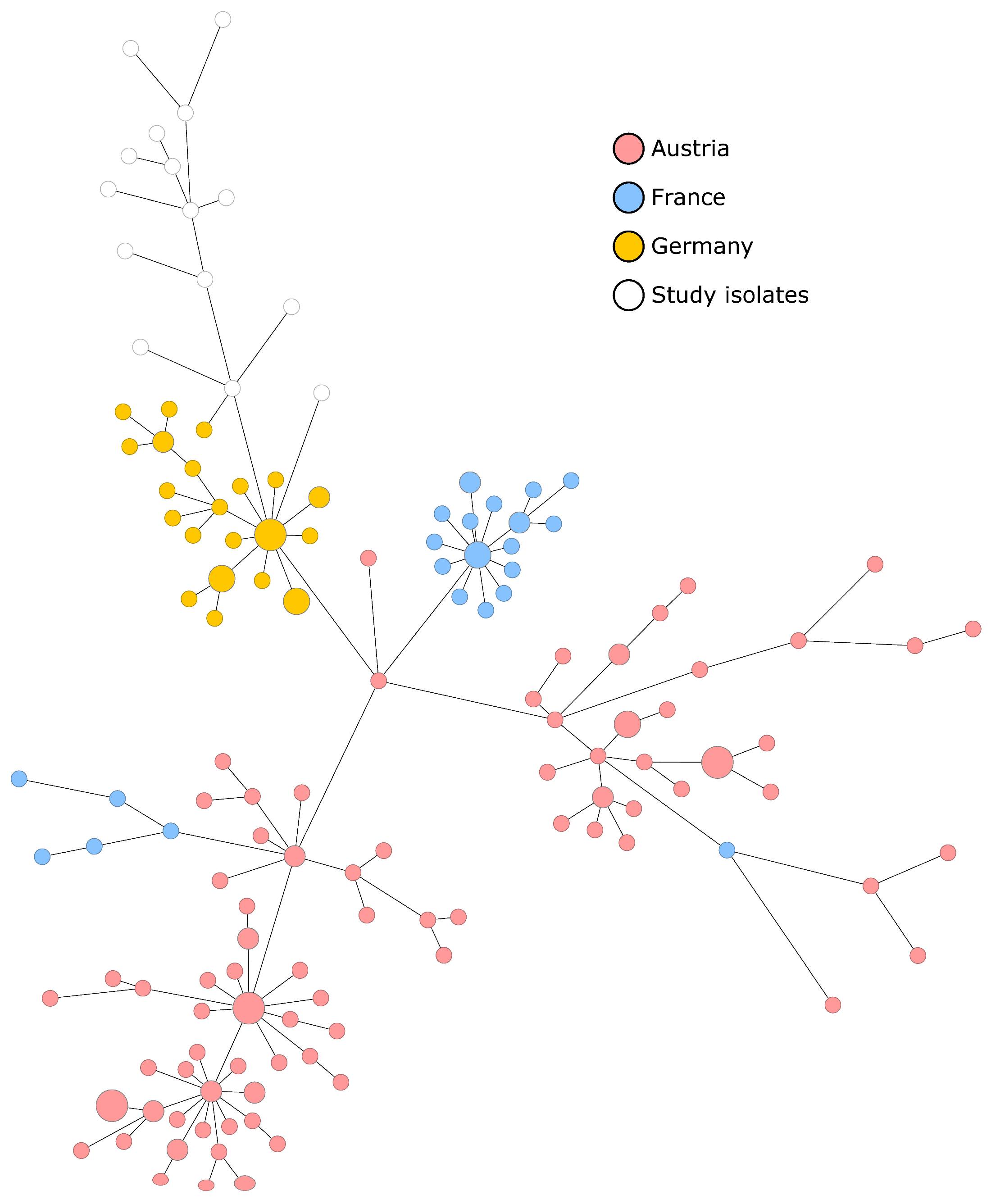
3.3. Molecular Characterization of Isolates and Genotypic Relationship
3.4. Comparison of European ST2 European Isolates Associated with the Hospital Setting
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Becker, K.; Both, A.; Weißelberg, S.; Heilmann, C.; Rohde, H. Emergence of coagulase-negative staphylococci. Expert Rev. Anti-infective Ther. 2020, 18, 349–366. [Google Scholar] [CrossRef]
- Michels, R.; Last, K.; Becker, S.L.; Papan, C. Update on Coagulase-Negative Staphylococci—What the Clinician Should Know. Microorganisms 2021, 9, 830. [Google Scholar] [CrossRef]
- Lee, E.; Anjum, F. StatPearls. In Staphylococcus Epidermidis Infection; National Center for Biotechnology Information: Bethesda, MD, USA, 2023. [Google Scholar]
- Pouget, C.; Chatre, C.; Lavigne, J.P.; Pantel, A.; Reynes, J.; Dunyach-Remy, C. Effect of Antibiotic Exposure on Staphylococcus epidermidis Responsible for Catheter-Related Bac-teremia. Int. J. Mol. Sci. 2023, 24, 1547. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, V.; Passerotto, R.A.; Chiuchiarelli, M.; Leanza, G.M.; Ojetti, V. Difficult-to-Treat Pathogens: A Review on the Man-agement of Multidrug-Resistant Staphylococcus epidermidis. Life 2023, 13, 1126. [Google Scholar]
- Vuong, C.; Voyich, J.M.; Fischer, E.R.; Braughton, K.R.; Whitney, A.R.; DeLeo, F.R.; Otto, M. Polysaccharide intercellular adhesin (PIA) protects Staphylococcus epidermidis against major components of the human innate immune system. Cell Microbiol. 2004, 6, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Swaney, S.M.; Aoki, H.; Ganoza, M.C.; Shinabarger, D.L. The oxazolidinone linezolid inhibits initiation of pro-tein synthesis in bacteria. Antimicrob. Agents Chemother. 1998, 42, 3251–3255. [Google Scholar] [CrossRef] [Green Version]
- Côrtes, M.F.; André, C.; Martins Simões, P.; Corvec, S.; Caillon, J.; Tristan, A.; Bes, M.; Vandenesch, F.; Figueiredo, A.M.; Dupieux, C.; et al. Persistence of a multidrug-resistant worldwide-disseminated methicillin-resistant Staphylococcus epidermidis clone harbouring the cfr linezolid resistance gene in a French hospital with evidence of interspecies transfer to several Staph-ylococcus aureus lineages. J. Antimicrob. Chemother. 2022, 77, 1838–1846. [Google Scholar]
- Bouiller, K.; Ilic, D.; Wicky, P.H.; Cholley, P.; Chirouze, C.; Bertrand, X. Spread of clonal linezolid-resistant Staphylococcus epidermidis in an intensive care unit associated with linezolid exposure. Eur. J. Clin. Microbiol. Infect. Dis 2020, 39, 1271–1277. [Google Scholar] [CrossRef]
- Alhumaid, S.; Al Mutair, A.; Al Alawi, Z.; Alzahrani, A.J.; Tobaiqy, M.; Alresasi, A.M.; Bu-Shehab, I.; Al-Hadary, I.; Alhmeed, N.; Alismail, M.; et al. Antimicrobial susceptibility of gram-positive and gram-negative bacteria: A 5-year retrospective analysis at a multi-hospital healthcare system in Saudi Arabia. Ann. Clin. Microbiol. Antimicrob. 2021, 20, 1–18. [Google Scholar] [CrossRef]
- Gebremariam, N.M.; Bitew, A.; Tsige, E.; Woldesenbet, D.; Tola, M.A. A High Level of Antimicrobial Resistance in Gram-Positive Cocci Isolates from Different Clinical Samples Among Patients Referred to Arsho Advanced Medical Laboratory, Addis Ababa, Ethiopia. Infect. Drug Resist. 2022, 15, 4203–4212. [Google Scholar] [CrossRef]
- Huber, S.; Knoll, M.A.; Berktold, M.; Würzner, R.; Brindlmayer, A.; Weber, V.; Posch, A.E.; Mrazek, K.; Lepuschitz, S.; Ante, M.; et al. Genomic and Phenotypic Analysis of Linezolid-Resistant Staphylococcus epidermidis in a Tertiary Hospital in Innsbruck, Austria. Microorganisms 2021, 9, 1023. [Google Scholar] [CrossRef]
- Coustillères, F.; Renault, V.; Corvec, S.; Dupieux, C.; Simões, P.M.; Lartigue, M.F.; Plouzeau-Jayle, C.; Tande, D.; Lamoureux, C.; Lemarié, C. Clinical, Bacteriological, and Genetic Characterization of Bone and Joint Infections Involving Linezolid-Resistant Staphylococcus epidermidis: A Retrospective Multicenter Study in French Reference Centers. Microbiol. Spectr. 2023, 11, e0419022. [Google Scholar] [CrossRef] [PubMed]
- Folan, S.A.; Marx, K.R.; Tverdek, F.P.; Raad, I.; Mulanovich, V.E.; Tarrand, J.J.; Shelburne, S.A.; Aitken, S.L. Clinical Outcomes Associated With Linezolid Resistance in Leukemia Patients With Linezol-id-Resistant Staphylococcus epidermidis Bacteremia. Open Forum Infect. Dis. 2018, 5, ofy167. [Google Scholar] [CrossRef] [PubMed]
- Lucas, C.R.; Rodicio, M.; Càmara, J.; Domínguez, M.; Alaguero, M.; Fernández, J. Long-term endemic situation caused by a linezolid- and meticillin-resistant clone of Staphylococcus epidermidis in a tertiary hospital. J. Hosp. Infect. 2020, 105, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Mulanovich, V.E.; Huband, M.D.; McCurdy, S.P.; Lemmon, M.M.; Lescoe, M.; Jiang, Y.; Rolston, K.V.I.; LaSala, P.R. Emergence of linezolid-resistant coagulase-negative Staphylococcus in a cancer centre linked to increased linezolid utilization. J. Antimicrob. Chemother. 2010, 65, 2001–2004. [Google Scholar] [CrossRef] [PubMed]
- Tsiodras, S.; Gold, H.S.; Sakoulas, G.; Eliopoulos, G.M.; Wennersten, C.; Venkataraman, L.; Moellering, R.C.; Ferraro, M.J. Linezolid resistance in a clinical isolate of Staphylococcus aureus. Lancet 2001, 358, 207–208. [Google Scholar] [CrossRef]
- Du, X.; Zhu, Y.; Song, Y.; Li, T.; Luo, T.; Sun, G.; Yang, C.; Cao, C.; Lu, Y.; Li, M. Molecular analysis of Staphylococcus epidermidis strains isolated from community and hospital envi-ronments in China. PLoS ONE 2013, 8, e62742. [Google Scholar]
- Feßler, A.T.; Calvo, N.; Gutiérrez, N.; Muñoz Bellido, J.L.; Fajardo, M.; Garduño, E.; Monecke, S.; Ehricht, R.; Kadlec, K.; Schwarz, S. Cfr-mediated linezolid resistance in methicillin-resistant Staphylococcus aureus and Staphylo-coccus hae-molyticus associated with clinical infections in humans: Two case reports. J. Antimicrob. Chemother. 2014, 69, 268–270. [Google Scholar] [CrossRef] [Green Version]
- Long, K.S.; Poehlsgaard, J.; Kehrenberg, C.; Schwarz, S.; Vester, B. The Cfr rRNA methyltransferase confers re-sistance to Phenicols, Lincosamides, Oxazolidinones, Pleuromutilins, and Streptogramin A antibiotics. Antimicrob. Agents Chemother. 2006, 50, 2500–2505. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; A Arias, C.; Aitken, S.L.; Peña, J.G.; Panesso, D.; Chang, M.; Diaz, L.; Rios, R.; Numan, Y.; Ghaoui, S.; et al. Clonal Emergence of Invasive Multidrug-Resistant Staphylococcus epidermidis Deconvoluted via a Combination of Whole-Genome Sequencing and Microbiome Analyses. Clin. Infect. Dis. 2018, 67, 398–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Kong, Z.; Liu, K.; Cheng, C.; Jiang, T.; Ma, P.; Li, R. Phenotypic and genotypic characterisation of linezolid-resistant coagulase-negative staphylococci possessing cfr-carrying plasmid. J. Glob. Antimicrob. Resist. 2022, 28, 226–232. [Google Scholar] [CrossRef]
- Bender, J.; Strommenger, B.; Steglich, M.; Zimmermann, O.; Fenner, I.; Lensing, C.; Dagwadordsch, U.; Kekulé, A.S.; Werner, G.; Layer, F. Linezolid resistance in clinical isolates of Staphylococcus epidermidis from German hospitals and characterization of two cfr-carrying plasmids. J. Antimicrob. Chemother. 2015, 70, 1630–1638. [Google Scholar] [CrossRef] [Green Version]
- Mauch, H.; Podbielski, A.; Hermann, M. MiQ 03a: Blutkulturdiagnostik–Sepsis, Endokarditis, Katheterinfektionen (Teil I), 2nd ed.; Elsevier Urban & Fischer: Munich, Germany, 2007. [Google Scholar]
- Mellmann, A.; Bletz, S.; Böking, T.; Kipp, F.; Becker, K.; Schultes, A.; Prior, K.; Harmsen, D. Real-Time Genome Sequencing of Resistant Bacteria Provides Precision Infection Control in an Institutional Setting. J. Clin. Microbiol. 2016, 54, 2874–2881. [Google Scholar] [CrossRef] [Green Version]
- Papan, C.; Schröder, M.; Hoffmann, M.; Knoll, H.; Last, K.; Albrecht, F.; Geisel, J.; Fink, T.; Gärtner, B.C.; Mellmann, A.; et al. Combined antibiotic stewardship and infection control measures to contain the spread of linezol-id-resistant Staphylococcus epidermidis in an intensive care unit. Antimicrob. Resist. Infect. Control. 2021, 10, 99. [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.H.; McDermott, P.F.; et al. Validating the AMRFinder Tool and Resistance Gene Database by Using Antimicrobial Re-sistance Gen-otype-Phenotype Correlations in a Collection of Isolates. Antimicrob. Agents Chemother. 2019, 63, 10–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dortet, L.; Glaser, P.; Kassis-Chikhani, N.; Girlich, D.; Ichai, P.; Boudon, M.; Samuel, D.; Creton, E.; Imanci, D.; Bonnin, R.; et al. Long-lasting successful dissemination of resistance to oxazolidinones in MDR Staphylococcus epi-dermidis clinical isolates in a tertiary care hospital in France. J. Antimicrob. Chemother. 2018, 73, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Santos, V.I.; Torres-Añorve, D.A.; Echániz-Aviles, G.; Parra-Rojas, I.; Ramírez-Peralta, A.; Castro-Alarcón, N. Characterization of Staphylococcus epidermidis clinical isolates from hospitalized patients with bloostream infection obtained in two time periods. PeerJ 2022, 10, e14030. [Google Scholar] [CrossRef]
- Ruiz-Ripa, L.; Feßler, A.T.; Hanke, D.; Eichhorn, I.; Azcona-Gutiérrez, J.M.; Alonso, C.A.; Pérez-Moreno, M.O.; Aspiroz, C.; Bellés, A.; Schwarz, S.; et al. Mechanisms of Linezolid Resistance Among Clinical Staphylococcus spp. in Spain: Spread of Methicillin- and Linezolid-Resistant S. epidermidis ST2. Microb. Drug Resist. 2021, 27, 145–153. [Google Scholar] [CrossRef]
- Martínez-Meléndez, A.; Morfín-Otero, R.; Villarreal-Treviño, L.; Camacho-Ortíz, A.; González-González, G.; Llaca-Díaz, J.; Rodríguez-Noriega, E.; Garza-González, E. Molecular epidemiology of coagulase-negative bloodstream isolates: Detection of Staphylococcus epidermidis ST2, ST7 and linezolid-resistant ST23. Braz. J. Infect. Dis. 2016, 20, 419–428. [Google Scholar] [CrossRef]
- Baos, E.; Candel, F.J.; Merino, P.; Pena, I.; Picazo, J.J. Characterization and monitoring of linezolid-resistant clini-cal isolates of Staphylococcus epidermidis in an intensive care unit 4 years after an outbreak of infection by cfr-mediated linezolid-resistant Staphylococcus aureus. Diagn. Microbiol. Infect. Dis. 2013, 76, 325–329. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Isolate | OXA | GEN | CIP | LEV | MXF | ERY | CL | TGC | DAP | VAN | TET | RIF | TMP | LZD | LZD (MIC mg/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P1 | R | R | R | R | R | I | R | S | S | S | R | R | R | R | >256 |
| P2 | R | R | R | R | R | R | R | S | S | S | R | R | R | R | >32 |
| P3 | R | R | R | R | R | R | R | S | S | S | R | R | R | R | >256 |
| P4 | R | R | R | R | R | R | R | S | S | S | R | R | R | R | >256 |
| P5 | R | R | R | R | R | R | R | S | S | S | R | R | S | R | >32 |
| P6 | R | R | R | R | R | R | R | S | S | S | R | R | R | R | >32 |
| P7 | R | R | R | R | R | R | R | S | S | S | R | R | R | R | >256 |
| P8 | R | R | R | R | R | R | R | S | S | S | R | R | R | R | >256 |
| P9 | R | R | R | R | R | R | R | S | S | S | R | R | S | R | >256 |
| P10 | R | R | R | R | R | R | R | S | S | S | R | R | R | R | >64 |
| P11 | R | R | R | R | R | R | R | S | S | S | R | R | S | R | >128 |
| P12 | R | R | R | R | R | R | R | S | S | S | R | R | R | R | >64 |
| P13 | R | R | R | R | R | R | R | S | S | S | R | R | R | R | >256 |
| P14 | R | R | R | R | R | R | R | S | S | S | R | R | R | R | >32 |
| P15 | R | R | R | R | R | R | R | S | S | S | R | R | R | R | >64 |
| Isolate | AK | GEN | KA | T | BLA | MC | F | FA | TMP |
|---|---|---|---|---|---|---|---|---|---|
| P11 | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | blaZ | mecA mecI mecR1 | fosB | fusB | dfrS1 |
| P9 | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | blaI blaZ | mecA mecI mecR1 | fosB | fusB | dfrS1 |
| P15 | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | blaI blaZ | mecA mecI mecR1 | fosB | fusB | dfrS1 |
| P10 | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | blaI blaZ | mecA mecI mecR1 | fosB | fusB | dfrS1 |
| P12 | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | blaI blaZ | mecA mecI mecR1 | fosB | fusB | dfrS1 |
| P13 | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | blaI blaZ | mecA mecI mecR1 | fosB | fusB | dfrS1 |
| P5 | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | blaI blaZ | mecA mecI mecR1 | fosB | fusB | dfrS1 |
| P14 | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | blaI blaZ | mecA mecI mecR1 | fosB | fusB | dfrS1 |
| P2 | aph(3′)-IIIa | aph(3′)-IIIa | blaI blaZ | mecA mecI mecR1 | fosB | fusB | dfrS1 | ||
| P4 | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | blaI blaZ | mecA mecI mecR1 | fosB | fusB | dfrS1 |
| P1 | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | blaI blaZ | mecA mecI mecR1 | fusB | fusB | dfrS1 |
| P6 | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | blaI blaZ | mecA mecI mecR1 | fosB | fusB | dfrS1 |
| P7 | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | blaI blaZ | mecA mecI mecR1 | fosB | fusB | dfrS1 |
| P3 | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | blaI blaZ | mecA mecI mecR1 | fosB | fusB | dfrS1 |
| P8 | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | aac(6′)-Ie/aph(2″)-Ia | blaI blaZ | mecA mecI mecR1 | fosB | fusB | dfrS1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campmann, F.; Tönnies, H.; Böing, C.; Schuler, F.; Mellmann, A.; Schwierzeck, V. Molecular Characterization of Clinical Linezolid-Resistant Staphylococcus epidermidis in a Tertiary Care Hospital. Microorganisms 2023, 11, 1805. https://doi.org/10.3390/microorganisms11071805
Campmann F, Tönnies H, Böing C, Schuler F, Mellmann A, Schwierzeck V. Molecular Characterization of Clinical Linezolid-Resistant Staphylococcus epidermidis in a Tertiary Care Hospital. Microorganisms. 2023; 11(7):1805. https://doi.org/10.3390/microorganisms11071805
Chicago/Turabian StyleCampmann, Florian, Hauke Tönnies, Christian Böing, Franziska Schuler, Alexander Mellmann, and Vera Schwierzeck. 2023. "Molecular Characterization of Clinical Linezolid-Resistant Staphylococcus epidermidis in a Tertiary Care Hospital" Microorganisms 11, no. 7: 1805. https://doi.org/10.3390/microorganisms11071805