
Ocular Bacterial Infections: A Ten-Year Survey and Review of Causative Organisms Based on the Oklahoma Experience
 ,
,  ,
,
Abstract
:1. Introduction
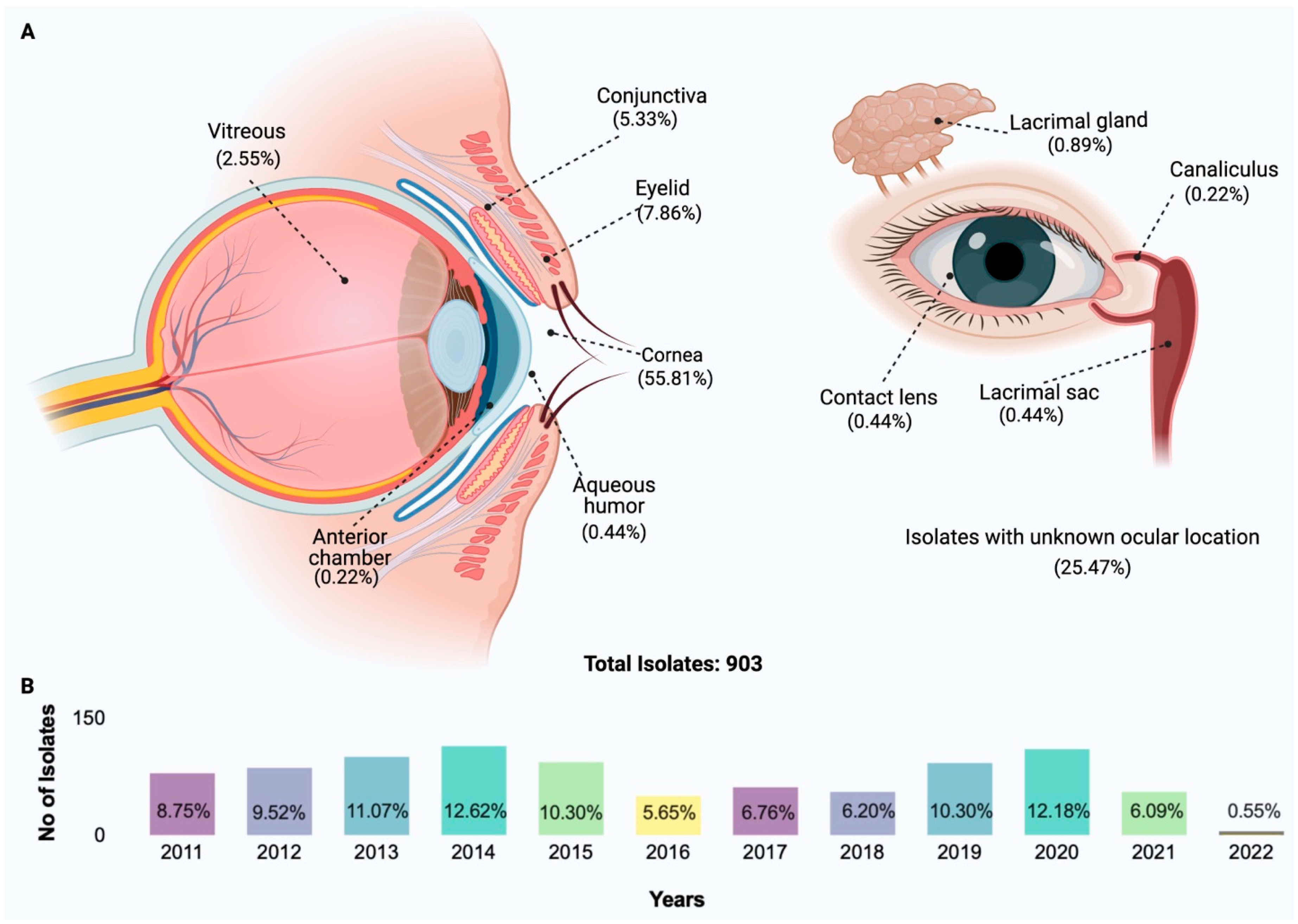
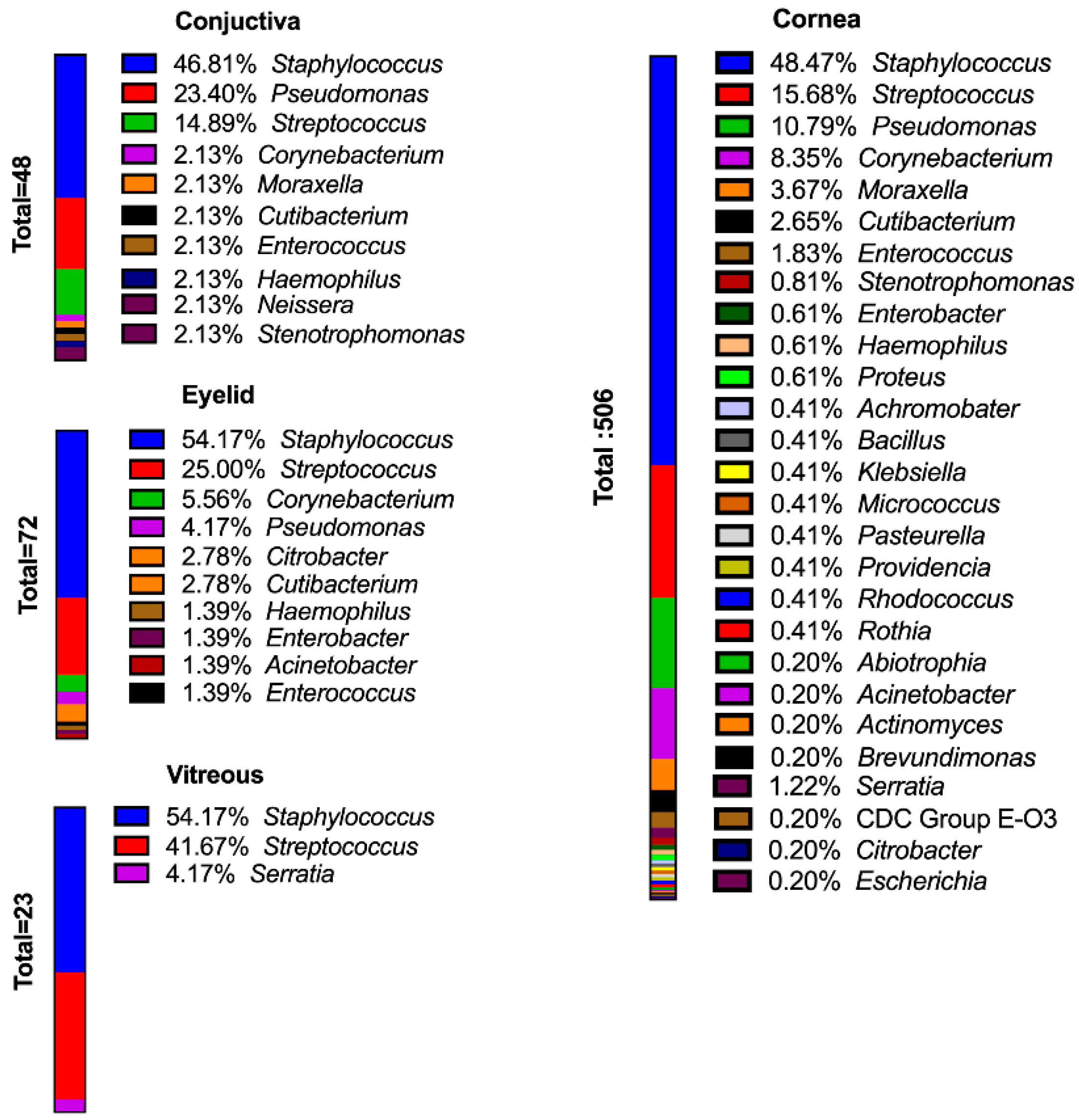
2. Species Distribution of Isolates
3. Gram-Positive Ocular Pathogens
3.1. Bacillus
3.2. Corynebacterium
3.3. Cutibacterium acnes (Propionibacterium acnes)
3.4. Enterococcus
3.5. Micrococcus
3.6. Staphylococci
3.6.1. Staphylococcus aureus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Study Population | Gram-Positive | Bacillus | CoNS | S. epidermidis | S. aureus | Gram-Negative | Pseudomonas | P. aeruginosa | |
|---|---|---|---|---|---|---|---|---|---|---|
| Arantes [88] | Brazil | Pre-cataract | 88.90% | 54% | 8% | 11.10% | ||||
| Capriotti [89] | Sierra Leone | Healthy Individuals | 78.1% | 5.5% | 36.1% | 25.1% | 21.9% | 7.8% | ||
| Dave [90] | USA, Nashville | Intravitreal Injections | 45.70% | 6.50% | 8.70% | |||||
| Dorrepaal [91] | Toronto | Intravitreal Injections | 64% | 1.80% | 0.90% | |||||
| Hsu [92] | USA, St. Louis | Pre-cataract | 90.5% | 74.8% | 57.2% | 5.0% | 9.5% | 1.8% | ||
| Lin [93] | Taiwan | Pre-cataract | 91.70% | 45.20% | 16.70% | 2.40% | 8.30% | 4.80% | ||
| Mamah [94] | Nigeria | Pre-cataract | 73.70% | 34.20% | 34.20% | 13.20% | 2.60% | |||
| Martins [95] | Sao Paulo | Healthy Individuals | 61.70% | 11.70% | 3.40% | |||||
| Mshangila [96] | Uganda | Pre-cataract | 65.90% | 76.90% | 21.00% | 10.10% | ||||
| Papa [97] | Italy | Pre-cataract | 95% | 67.90% | 58.00% | 15.30% | 4.60% | |||
| Rubio [98] | Madrid | Pre-cataract | 56.80% | 6.40% | 7.30% | |||||
| Suto [99] | Japan | Pre-cataract | 67% | 57.20% | 3.90% | 6.30% | 0.70% |
3.6.2. Coagulase-Negative Staphylococci
3.6.3. Staphylococcus pseudintermedius
3.7. Streptococci
4. Gram-Negative Ocular Pathogens
4.1. Achromobacter
4.2. Acinetobacter
4.3. Citrobacter
4.4. Enterobacter
4.5. Escherichia coli
4.6. Haemophilus
4.7. Klebsiella
4.8. Moraxella
4.9. Pseudomonas
4.10. Proteus mirabilis
4.11. Serratia
4.12. Stenotrophomonas maltophilia
5. Hapax Legomenon Isolates
5.1. Abiotrophia, Actinomyces, and Brevundimonas diminuta
5.2. CDC Group EO-3, Cronobacter, and Lactobacillus
5.3. Neisseria
5.4. Pasteurella
5.5. Providencia
5.6. Rhodococcus
5.7. Rothia
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Armstrong, R.A. The microbiology of the eye. Ophthal. Physiol. Opt. 2000, 20, 429–441. [Google Scholar] [CrossRef]
- Ung, L.; Chodosh, J. Foundational concepts in the biology of bacterial keratitis. Exper. Eye Res. 2021, 209, 108647. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.; Iovieno, A. The Role of microbial flora on the ocular surface. Curr. Opin. Allergy Clin. Immunol. 2009, 9, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Hovding, G. Acute bacterial conjunctivitis. Acta Ophthalmol. 2008, 86, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Azari, A.A.; Barney, N.P. Conjunctivitis A Systematic Review of Diagnosis and Treatment. JAMA 2013, 310, 1721–1729. [Google Scholar] [CrossRef] [PubMed]
- Lichtinger, A.; Yeung, S.N.; Kim, P.; Amiran, M.D.; Iovieno, A.; Elbaz, U.; Ku, J.Y.F.; Wolff, R.; Rootman, D.S.; Slomovic, A.R. Shifting Trends in Bacterial Keratitis in Toronto an 11-Year Review. Ophthalmology 2012, 119, 1785–1790. [Google Scholar] [CrossRef] [PubMed]
- Benz, M.S.; Scott, I.U.; Flynn, H.W., Jr.; Unonius, N.; Miller, D. Endophthalmitis Isolates and Antibiotic Sensitivities: A 6-Year Review of Culture-proven Cases. Am. J. Ophthalmol. 2004, 137, 38–42. [Google Scholar] [CrossRef]
- Olson, R.; Donnenfeld, E.; Bucci, F.A., Jr.; Raizman, M.; Solomon, K.; Devgan, U.; Trattler, W.; Dell, S.; Wallace, R.B.; Callegan, M.; et al. Methicillin resistance of Staphylococcus species among health care and nonhealth care workers undergoing cataract surgery. Clin. Ophthal. 2010, 4, 1505–1514. [Google Scholar] [CrossRef] [Green Version]
- Al-Dhaheri, H.S.; Al-Tamimi, M.D.; Khandekar, R.B.; Dip, P.G.; Khan, M.; Stone, D.U. Ocular Pathogens and Antibiotic Sensitivity in Bacterial Keratitis Isolates at King Khaled Eye Specialist Hospital, 2011 to 2014. Cornea 2016, 35, 789–794. [Google Scholar] [CrossRef]
- Booth, M.; Pence, L.; Mahasreshti, P.; Callegan, M.; Gilmore, M. Clonal Associations among Staphylococcus aureus Isolates from Various Sites of Infection. Infect. Immun. 2001, 69, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Lakhundi, S.; Siddiqui, R.; Khan, N. Pathogenesis of microbial keratitis. Microb. Pathog. 2016, 104, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, K.; Deepthi, G.; Prabagaran, S. Ocular bacterial infections: Pathogenesis and diagnosis. Microb. Pathog. 2020, 145, 104206. [Google Scholar]
- Bernard, K. The Genus Corynebacterium and other Medically Relevant Coryneform-Like Bacteria. J. Clin. Microbiol. 2012, 50, 3152–3158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, T.; Kitazawa, K.; Deguchi, H.; Sotozono, C. Current Evidence of Corynebacterium on the Ocular Surface. Microorganisms 2021, 9, 254. [Google Scholar] [CrossRef]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus species in soil as a natural resource for plant health and nutrition. J. Appl. Microbiol. 2020, 128, 1583–1594. [Google Scholar] [CrossRef] [Green Version]
- Ehling-Schulz, M.; Lereclus, D.; Koehler, T.M. The Bacillus cereus Group: Bacillus Species with Pathogenic Potential. Microbiol. Spectr. 2019, 7, 10–1128. [Google Scholar] [CrossRef]
- Choudhuri, K.; Sharma, S.; Garg, P.; Rao, G. Clinical and Microbiological Profile of Bacillus Keratitis. Cornea 2000, 19, 301–306. [Google Scholar] [CrossRef]
- Donzis, P.; Mondino, B.; Weissman, B. Bacillus Keratitis Associated with Contaminated Contact Lens Care Systems. Am. J. Ophthalmol. 1988, 105, 195–197. [Google Scholar] [CrossRef]
- Pinna, A.; Sechi, L.; Zanetti, S.; Usai, D.; Delogu, G.; Capuccinelli, P.; Carta, F. Bacillus cereus Keratitis Associated with Contact Lens Wear. Ophthalmology 2001, 108, 1830–1834. [Google Scholar] [CrossRef]
- Pitt, T.; McClure, J.; Parker, M.; Amezquita, A.; McClure, P. Bacillus cereus in personal care products: Risk to consumers. Internat. J. Cosmet. Sci. 2015, 37, 165–174. [Google Scholar] [CrossRef]
- Land, A.; Penno, K.; Brzezinski, J. Identification of Microorganisms Isolated from Counterfeit and Unapproved Decorative Contact Lenses. J. Forensic. Sci. 2018, 63, 635–639. [Google Scholar] [CrossRef]
- Ramos-Esteban, J.; Servat, J.; Tauber, S.; Bia, F. Bacillus megaterium Delayed Onset Lamellar Keratitis after LASIK. J. Refact. Surg. 2006, 22, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Callegan, M.; Cochran, D.; Kane, S.; Ramadan, R.; Chodosh, J.; McLean, C.; Stroman, D. Virulence Factor Profiles and Antimicrobial Susceptibilities of Ocular Bacillus Isolates. Curr. Eye Res. 2006, 31, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Das, T.; Choudhury, K.; Sharma, S.; Jalali, S.; Nuthethi, R.; Endophthalmitis Research Group. Clinical Profile and Outcome in Bacillus Endophthalmitis. Am. Acad. Ophthal. 2001, 108, 1819–1825. [Google Scholar] [CrossRef] [PubMed]
- Mursalin, M.H.; Livingston, E.T.; Callegan, M.C. The cereus matter of Bacillus endophthalmitis. Exp. Eye Res. 2020, 193, 107959. [Google Scholar] [CrossRef]
- Coburn, P.; Miller, F.; Enty, M.; Land, C.; LaGrow, A.; Mursalin, M.; Callegan, M. The Bacillus virulome in endophthalmitis. Microbiology 2021, 167, 001057. [Google Scholar] [CrossRef]
- Callegan, M.; Engelbert, M.; Parke, D.; Jett, B.; Gilmore, M. Bacterial Endophthalmitis: Epidemiology, Therapeutics, and Bacterium-Host Interactions. Clin. Microbiol. Rev. 2002, 15, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Bhagat, N.; Nagori, S.; Zarbin, M. Post-traumatic Infectious Endophthalmitis. Surv. Ophthalmol. 2011, 59, 214–251. [Google Scholar] [CrossRef]
- Coburn, P.; Miller, F.; LaGrow, A.; Land, C.; Mursalin, H.; Livingston, E.; Amayem, O.; Chen, Y.; Gao, W.; Zhang, L.; et al. Disarming Pore-Forming Toxins with Biomimetic Nanosponges in Intraocular Infections. mSphere 2019, 4, e00262-19. [Google Scholar] [CrossRef] [Green Version]
- Wiskur, B.J.; Robinson, M.L.; Farrand, A.J.; Novosad, B.D.; Callegan, M.C. Toward improving therapeutic regimens for Bacillus endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4931–4938. [Google Scholar] [CrossRef] [Green Version]
- Sugumaran, R.; Sistla, S.; Chavhan, P.; Deb, A. Corynebacterium amycolatum: An unusual cause of corneal ulcer. BMJ Case Rep. 2020, 13, e237818. [Google Scholar] [CrossRef] [PubMed]
- Badenoch, P.; O’Daniel, L.; Wise, R.; Slattery, J.; Mills, R. Corynebacterium propinquum Keratitis Identified Using MALDI-TOF. Cornea 2016, 35, 686–687. [Google Scholar] [CrossRef] [PubMed]
- Duignan, E.; Farrell, S.; Treacy, M.; Fulcher, T.; O’Brien, P.; Power, W.; Murphy, C. Corneal inlay, implantation complicated by infectious keratitis. Br. J. Ophthalmol. 2016, 100, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Rubinfeld, R.S.; Cohen, E.J.; Arentsen, J.J.; Laibson, P.R. Diphtheroids as Ocular Pathogens. Am. J. Ophthalmol. 1989, 108, 251–254. [Google Scholar] [CrossRef]
- Sagerfors, S.; Poehlein, A.; Afshar, M.; Lindblad, B.; Bruggemann, H.; Soderquist, B. Clinical and genomic features of Corynebacterium macginleyi-associated infectious keratitis. Sci. Rep. 2021, 11, 6015. [Google Scholar] [CrossRef]
- Hoshi, S.; Hashida, M.; Urabe, K. Risk factors for aerobic bacterial conjunctival flora in preoperative cataract patients. Eye 2016, 30, 1439–1446. [Google Scholar] [CrossRef] [Green Version]
- Tejada, E. Endocarditis and aortic absess due to Corynebacterium macginleyi. Rev. Clin. Esp. 2007, 207, 478–479. [Google Scholar] [CrossRef]
- Cacopardo, B.; Stefani, S.; Cardi, F.; Cardi, C.; Pinzone, M.; Nunnari, G. Surgical Site Infection by Corynebacterium macginleyi in a Patient with Neurofibromatosis Type 1. Case Rep. Infect. Dis. 2013, 2013, 970678. [Google Scholar]
- Villanueva, J.; Dominguez, A.; Rios, M.; Iglesias, C. Corynebacterium macginleyi Isolated from Urine in a Patient with a Permanent Bladder Catheter. Scand. J. Infect. Dis. 2002, 34, 699–700. [Google Scholar] [CrossRef]
- Qin, V.; Laurent, T.; Ledoux, A. Corynebacterium macginleyi-associated Blebitis: A Case Report. J. Glauc. 2018, 27, e174–e176. [Google Scholar] [CrossRef]
- Eguchi, H.; Kuwahara, T.; Miyamoto, T.; Nakayama-Imaohji, H.; Ichimura, M.; Hayashi, T.; Shiota, H. High-Level Fluoroquinolone Resistance in Ophthalmic Clinical Isolates Belonging to the Species Corynebacterium macginleyi. J. Clin. Microbiol. 2008, 46, 527–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrer, C.; Ruiz-Moreno, J.M.; Rodriquez, A.; Montero, J.; Alio, J.L. Postoperative Corynebacterium macginleyi endophthalmitis. J. Cataract. Refract. Surg. 2004, 30, 2441–2444. [Google Scholar] [CrossRef] [PubMed]
- Ruoff, K.; Toutain-Kidd, C.; Srinivasan, M.; Lalitha, P.; Acharya, N.; Zegans, M.; Schwartzman, J. Corynebacterium macginleyi isolated from a corneal ulcer. Inf. Dis. Rep. 2010, 2, e3. [Google Scholar] [CrossRef] [Green Version]
- St. Leger, A.; Caspi, R.R. Visions of Eye Commensals: The Known and Unknown About How the Microbiome Affects Eye Disease. BioEssays 2018, 40, e1800046. [Google Scholar] [CrossRef]
- Rigas, Y.; Treat, B.R.; Shane, J.; Shanks, R.; St. Leger, A.J. Genetic Manipulation of Corynebacterium mastitidis to Better Understand the Ocular Microbiome. Investig. Ophthalmol. Vis. Sci. 2023, 64, 19. [Google Scholar] [CrossRef]
- Perry, A.; Lambert, P. Propionibacterium acnes. Lett. Appl. Microbiol. 2006, 42, 185–188. [Google Scholar] [CrossRef]
- Achermann, Y.; Goldstein, E.; Coenye, T.; Shirtliff, M. Propionibacterium acnes: From Commensal to Opportunistic Biofilm-Associated Implant Pathogen. Clin. Microbiol. Rev. 2014, 27, 419–440. [Google Scholar] [CrossRef] [Green Version]
- Doan, T.; Akileswaran, L.; Andersen, D.; Johnson, B.; Ko, N.; Shrestha, A.; Shestopalov, V.; Lee, C.; Lee, A.; Van Gelder, R. Paucibacterial Microbiome and Resident DNA Virome of the Healthy Conjunctiva. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5116–5126. [Google Scholar] [CrossRef] [Green Version]
- Ovodenko, B.; Seedor, J.; Ritterband, D.; Shah, M.; Yang, R.; Koplin, R. The Prevalence and Pathogenicity of Propionibacterium acnes Keratitis. Cornea 2009, 28, 36–39. [Google Scholar] [CrossRef]
- Lim, S.; Na, K.; Joo, C. Clinical Features of Infectious Keratitis Caused by Propionibacterium Acnes. Eye Contact Lens 2017, 43, 330–333. [Google Scholar] [CrossRef]
- Durand, M.L. Bacterial and Fungal Endophthalmitis. Clin. Microbiol. Rev. 2017, 30, 597–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowler, B.; Miller, D.; Yan, X.; Yannuzzi, N.; Flynn, H. Postoperative Endophthalmitis Caused by Cutibacterium (Formerly Propionibacterium) Acnes: Case Series and Review. Case Rep. Ophthalmol. 2021, 12, 1–10. [Google Scholar] [CrossRef]
- Dreno, B.; Pecastaings, S.; Corvec, S.; Veraldi, S.; Khammari, A.; Roques, C. Cutibacterium acne (Propionibacterium acnes) and acne vulgaris: A brief look at the latest updates. JEADV 2018, 32, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Gunthard, H.; Hany, A.; Turina, M.; Wust, J. Propionibacterium acnes as a Cause of Aggressive Aortic Valve Endocarditis and Importance of Tissue Grinding: Case Report and Review. J. Clin. Microbiol. 1994, 32, 3043–3045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiore, E.; Van Tyne, D.; Gilmore, M.S. Pathogenicity of Enterococci. Microbiol. Spectr. 2019, 7, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Vu, J.; Carvalho, J. Enterococcus: Review of its physiology, pathogenesis, diseases and the challenges it poses for clinical microbiology. Front. Biol. 2011, 6, 357–366. [Google Scholar] [CrossRef]
- Endophthalmitis Vitrectomy Study Group. Microbiologic factors and visual outcome in the Endophthalmitis Vitrectomy Study. Am. J. Ophthalmol. 1996, 122, 830–846. [Google Scholar] [CrossRef]
- Lemley, C.; Han, D. Endophthalmitis: A review of current evaluation and management. Retina 2007, 27, 662–680. [Google Scholar] [CrossRef]
- Mandelbaum, S.; Forster, R.K. Endophthalmitis associated with filtering blebs. Int. Ophthalmol. Clin. 1987, 27, 107–111. [Google Scholar] [CrossRef]
- Scott, I.U.; Loo, R.H.; Flynn, R.W.; Miller, D. Endophthalmitis caused by Enterococcus faecalis. Ophthalmology 2003, 110, 1573–1577. [Google Scholar] [CrossRef]
- Weber, D.J.; Hoffman, K.L.; Thoft, R.A.; Baker, A.S. Endophthalmitis following intraocular lens implantation: Report of 30 cases and review of the literature. Rev. Infect. Dis. 1986, 8, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Archimbaud, C.; Shankar, N.; Forestier, C.; Baghdayan, A.; Gilmore, C.F.; Joly, B. In vitro adhesive properties and virulence factors of Enterococcus faecalis strains. Res. Microbiol. 2002, 153, 75–80. [Google Scholar] [CrossRef]
- Van Tyne, D.; Martin, M.; Gilmore, M. Structure, Function, and Biology of the Enterococcus faecalis Cytolysin. Toxins 2013, 5, 895–911. [Google Scholar] [CrossRef] [Green Version]
- Jett, B.; Jensen, H.; Nordquist, R.; Gilmore, M. Contribution of the pAD1-Encoded Cytolysin to the Severity of Experimental Enterococcus faecalis Endophthalmitis. Infect. Immun. 1992, 60, 2445–2452. [Google Scholar] [CrossRef] [Green Version]
- Gilmore, M.S.; Coburn, P.S.; Nallapareddy, S.R.; Murray, B.E. Enterococcal virulence. In The Enterococci: Pathogenesis, Molecular Biology, and Antibiotic Resistance; ASM Press: Washington, DC, USA, 2002; pp. 301–354. [Google Scholar]
- De Oliveira, D.; Forde, B.; Kidd, T.; Harris, P.; Schembri, M.; Beatson, S.; Paterson, D.; Walker, M. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef] [PubMed]
- Taneja, M.; Rathi, V.M.; Bagga, B.; Murthy, S.I.; Ashar, J.; Reddy, A.K.; Vaddavalli, P.K. Micrococcus keratitis following microkeratome-assisted laser in situ keratomileusis. Oman J. Ophthalmol. 2019, 12, 203–205. [Google Scholar] [PubMed]
- Busse, H. Micrococcus. In Bergey’s Manual of Systematics of Archaea and Bacteria, Online; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar]
- Zhu, M.; Zhu, Q.; Yang, Z.; Liang, Z. Clinical characteristics of patients with Micrococcus luteus bloodstream infection in a Chinese Tertiary-Care Hospital. Pol. J. Microbiol. 2021, 70, 321–326. [Google Scholar] [CrossRef]
- Albertson, D.; Natisos, G.; Gleckman, R. Septic Shock with Micrococcus luteus. Arch. Inter. Med. 1978, 138, 487–488. [Google Scholar] [CrossRef]
- Fosse, T.; Peloux, Y.; Granthil, C.; Toga, B.; Bertrando, J.; Sethian, M. Meningitis due to Micrococcus luteus. Infection 1985, 13, 280–281. [Google Scholar] [CrossRef]
- Oudiz, R.; Widlitz, A.; Beckmann, X.; Camnga, D.; Alfie, J.; Brundage, B.; Barst, R. Micrococcus-Associated Central Venous Catheter Infection in Patients with Pulmonary Arterial Hypertension. Chest 2004, 126, 90–94. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Nava, G.; Mohamed, A.; Yanez-Bello, M.; Trelles-Garcia, D. Advances in medicine and positive natural selection: Prosthetic valve endocarditis due to biofilm producer Micrococcus luteus. IDCases 2020, 20, e00743. [Google Scholar] [CrossRef] [PubMed]
- Hennekinne, J.; De Buyser, M.; Dragacci, S. Staphylococcus aureus and its food poisoning toxins: Characterization and outbreak investigation. FEMS Microbiol. Rev. 2012, 36, 815–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ondusko, D.; Nolt, D. Staphylococcus aureus. Ped. Rev. 2018, 39, 287–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, G.; Carrol, K.; Butel, J.; Morse, S.J. Melnick & Adelberg’s Medical Microbiology, 26th ed.; McGraw Hill: New York, NY, USA, 2012. [Google Scholar]
- Otto, M. Staphylococcus epidermidis—The ‘accidental’ pathogen. Nat. Rev. 2009, 7, 555–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksen, N.; Espersen, F.; Rosdahl, V.; Jensen, K. Carrage of Staphyloccus aureus among 104 healty persons during a 19-month period. Epidemiol. Infect. 1995, 115, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Putnam, C.M. Diagnosis and management of blepharitis: An optometrist’s prospective. Clin. Optom. 2016, 8, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Chira, S.; Miller, L. Staphylococcus aureus is the most common identified cause of cellulitis: A systemic review. Epidemiol. Infect. 2010, 138, 313–317. [Google Scholar] [CrossRef] [Green Version]
- Harford, D.; Greenan, E.; Knowles, S.; Fitzgerald, S.; Murphy, C. The burden of methicillin-resistant, Staphylococcus aureus in the delivery of eye care. Eye 2021, 36, 1368–1372. [Google Scholar] [CrossRef]
- Bourcier, T.; Thomas, F.; Borderie, V.; Chaumeil, C.; Laroche, L. Bacterial keratitis: Predisposing factors, clinical and microbiological review of 300 cases. Br. J. Ophthalmol. 2003, 87, 834–838. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.; Chayani, N.; Mohapatra, D.; Barik, M.R.; Sharma, S.; Basu, S. High prevalence of biofilm-forming MRSA in the conjunctival flora in chronic dacryocystitis. Semin. Ophthalmol. 2019, 34, 74–79. [Google Scholar] [CrossRef]
- Luo, B.; Li, M.; Xiang, N.; Hu, W.; Liu, R.; Xiaoqin, Y. The microbiologic spectrum of dacryocystitis. BMC Ophthalmol. 2021, 21, 29. [Google Scholar] [CrossRef]
- Yannuzzi, N.A.; Si, N.; Relhan, N.; Kuriyan, A.E.; Albini, T.A.; Berrocal, A.M.; Davis, J.L.; Smiddy, W.E.; Townsend, J.; Miller, D.; et al. Endophthalmitis After Clear Corneal Cataract Surgery: Outcomes Over Two Decades. Am. J. Ophthalmol. 2016, 174, 155–159. [Google Scholar] [CrossRef] [Green Version]
- Grumann, D.; Nubel, U.; Broker, B. Staphylococcus aureus toxins—Their functions and genetics. Infect. Genet. Evol. 2013, 21, 583–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astley, R.; Miller, F.; Mursalin, M.; Coburn, P.; Callegan, M. An Eye on Staphylococcus aureus Toxins: Roles in Ocular Damage and Inflammation. Toxins 2019, 11, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arantes, T.; Cavalcanti, R.; de Fatima Alves Diniz Severo, M.; Neto, J.; Machado Barbosa de Castro, C. Conjunctival bacterial flora and antibiotic resistance patterns in patients undergoing cataract surgery. Arq. Bras. Oftamol. 2006, 69, 33–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capriotti, J.A.; Pelletier, J.S.; Shah, M.; Caivano, D.M.; Ritterband, D.C. Normal ocular flora in heathy eyes from a rural population in Sierra Leone. Int. Ophthalmol. 2009, 29, 81–84. [Google Scholar] [CrossRef]
- Dave, S.B.; Hassanain, S.T.; Stephen, J.K.; Kim, M.D. Changes in Ocular Flora in Eyes Exposed to Ophthalmic Antibiotics. Ophthalmology 2013, 120, 937–941. [Google Scholar] [CrossRef]
- Dorrepaal, S.J.; Gale, J.; El-Defrawy, S.; Sharma, S. Resistance of ocular flora to gatifloxacin in patients undergoing intravitreal injections. Can. J. Ophthalmol. 2014, 49, 66–71. [Google Scholar] [CrossRef]
- Hsu, H.; Lind, J.T.; Tseng, L.; Miller, D. Ocular Flora and Their Antibiotic Resistance Patterns in the Midwest: A Prospective Study of Patients Undergoing Cataract Surgery. Am. J. Ophthalmol. 2013, 155, 36–44. [Google Scholar] [CrossRef]
- Lin, Y.; Kang, Y.; Hou, C.; Huang, Y.; Chen, C.; Shu, J.; Hsieh, P.; Hsiao, C. Antibiotic susceptibility profiles of acular and nasal flora in patients undergoing cataract surgery in Tiawan: An observational and cross-sectional study. BMJ Open 2017, 7, 66–77. [Google Scholar] [CrossRef] [Green Version]
- Mamah, C.; Anyalebechi, O.; Onwubiko, S.; Okoloagu, M.; Maduka-Okafor, F.; Ebede, S.; Umeh, R. Conjunctival bacterial flora and their antibiotic sensitivity among patients scheduled for cataract surgery in a tertiary hospital in south-east Nigeria. Graefes Arch. Clin. Exp. Ophthalmol. 2020, 259, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Martins, E.N.; Alvarenga, L.S.; Hofing-Lima, A.L.; Freitas, D.; Zorat-Yu, M.C.; Farah, M.E.; Mannis, M.J. Aeraobic Bacterial Conjunctival Flora in Diabetic Patients. Cornea 2004, 2, 136–142. [Google Scholar] [CrossRef]
- Mshangila, B.; Paddy, M.; Kajumbula, H.; Ateenyi-Agaba, C.; Kahwa, B.; Seni, J. External ocular surface bacterial isolates and their antimicrobial susceptibility patterns among pre-operative cataract patients at Mulago National Hospital in Kampala, Uganda. BMC Ophthalmol. 2013, 13, 71. [Google Scholar] [CrossRef] [Green Version]
- Papa, V.; Blanco, A.R.; Santocono, M. Ocular flora and their antibiotic susceptibility in patients having cataract surgery in Italy. J. Cataract Refract. Surg. 2016, 42, 1312–1317. [Google Scholar] [CrossRef]
- Rubio, E. Climatic influence on conjunctival bacteria of patients undergoing cataract surgery. Eye 2004, 18, 778–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suto, C.; Morinaga, M.; Yagi, T.; Tsuji, C.; Toshida, H. Conjunctival sac bacterial flora isolated prior to cataract surgery. Infect. Drug Resist. 2012, 5, 37–41. [Google Scholar] [PubMed] [Green Version]
- Glenny, A.T.; Stevens, M.F. Staphylococcus Toxins and Antitoxins. J. Pathol. Bacteriol. 1935, 40, 201–210. [Google Scholar] [CrossRef]
- Berube, B.; Wardenburg, J. Staphylococcus aureus—Toxin: Nearly a Century of Intrigue. Toxins 2013, 5, 1140–1166. [Google Scholar] [CrossRef] [Green Version]
- Callegan, M.C.; Engel, L.S.; Hill, J.M.; O’Callaghan, R.J. Corneal Virulence of Staphylococcus aureus: Roles of Alpha-Toxin and Protein A in Pathogenesis. Infect. Immun. 1994, 62, 2478–2482. [Google Scholar] [CrossRef] [Green Version]
- Girgis, D.O.; Sloop, G.D.; Reed, J.M.; O’Callaghan, R.J. Effects of Toxin Production in a Murine Model of Staphylococcus aureus Keratitis. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2064–2070. [Google Scholar] [CrossRef] [Green Version]
- Putra, I.; Rabiee, B.; Anwar, K.N.; Gidfar, S.; Shen, X.; Babalooee, M.; Grassemi, M.; Afsharkhamseh, N.; Bakhsh, S.; Missiakas, D.; et al. Staphylococcus aureus Alpha-Hemolysin Impairs Corneal Epithelial Wound Healing and Promotes Intracellular bacterial Invasion. Exp. Eye Res. 2019, 181, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, A. Role of Staphylococcus aureus Virulence Factors in Inducing Inflammation and Vascular Permeability in a Mouse Model of Bacterial Endophthalmitis. PLoS ONE 2015, 10, e0128423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wang, D.; Duan, Y.; Zhou, Z.; Gao, W.; Zhang, L. Cellular Nanosponges for Biological Neutralization. Adv. Mater. 2022, 34, 2107719. [Google Scholar] [CrossRef] [PubMed]
- Jarraud, S.; Mougel, C.; Thioulouse, J.; Lina, G.; Meugnier, H.; Forey, F.; Nesme, X.; Etienne, J.; Vandernesch, F. Relationships between Staphylococcus aureus Genetic Background, Virulence Factors agr Groups (Alleles), and Human Disease. Infect. Immun. 2002, 70, 631–641. [Google Scholar] [CrossRef] [Green Version]
- Aarestrup, F.; Larsen, H.; Eriksen, N.; Elsberg, C.; Jensen, N. Frequency of α- and β- haemolysin in Staphylococcus aureus of bovine and human origin. Apmis 1999, 170, 425–430. [Google Scholar] [CrossRef]
- Freer, J.H.; Arbuthnott, J.P. Toxins of Staphylococcus aureus. Pharmacol. Ther. 1983, 19, 55–106. [Google Scholar] [CrossRef]
- Goerke, C.; Wirtz, C.; Fluckiger, U.; Wolz, C. Extensive phage dynamics in Staphylococcus aureus contributes to adaptation to the human host during infection. Mol. Microbiol. 2006, 61, 1673–1685. [Google Scholar] [CrossRef]
- Salgado-Pabon, W.; Herrera, A.; Vu, B.; Stach, C.; Merriman, J.; Spaulding, A.; Schlievert, P. Staphylococcus aureus β-toxin Production is Common in Strains With the β-toxin Gene Inactivation by Bacteriophage. J. Infect. Dis. 2014, 210, 784–792. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan, R.J.; Callegan, M.C.; Moreau, J.M.; Green, L.C.; Foster, T.J.; Hartford, O.M.; Engel, L.S.; Hill, J.M. Specific roles of alpha-toxin and beta-toxin during Staphylococcus aureus corneal infection. Infect. Immun. 1997, 65, 1571–1578. [Google Scholar] [CrossRef] [Green Version]
- Spann, A.; Schiepers, A.; de Haas, C.; van Hooijdonk, D.; Badiou, C.; Contamin, H.; Vandenesch, F.; Lina, G.; Gerard, N.; Gerard, C.; et al. Differential Interactions of the Staphylococcal Toxins Panton-Valentine Leucocidin and Hemolysin CG with Human C5a Receptors. J. Immunol. 2015, 195, 1034–1043. [Google Scholar] [CrossRef] [Green Version]
- Wannet, W. Virulent MRSA strains containing Panton Valentine Leukociden gene in the Netherlands. Euro Surveil. 2003, 7, 2173. [Google Scholar]
- Holmes, A.; Ganner, M.; McGaune, P.B.; Kearns, A. Staphylococcus aureus Isolates Carrying Panton-Valentine Leucocidin Genes in England and Wales: Frequency, Characterizations, and Association with Clinical Disease. J. Clin. Microbiol. 2005, 43, 2384–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bispo, P.; Ung, L.; Chodosh, J.; Gilmore, M. Hospital-Associated Multidrug-Resistant MRSA Lineages Are Tropic to the Ocular Surface and Cause Severe Microbiocidal Keratitis. Front. Public Health 2020, 8, 204. [Google Scholar] [CrossRef]
- Foster, C.; Yarotsky, E.; Mason, E.; Kaplan, S.; Hulten, K. Molecular Characterization of Staphylococcus aureus Isolates from Children With Periorbital or Orbital Cellulitis. J. Ped. Infect. Dis. Soc. 2017, 3, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Sueke, H.; Shankar, J.; Neal, T.; Winstanley, C.; Tuft, S.; Coates, R.; Horsbourgh, M.J.; Kaye, S.; Microbiology Ophthalmic Group. lukSF-PV in Staphylococcus aureus Keratitis Isolates and Association with Clinical Outcomes. Investig. Ophthalmol. Vis. Sci. 2013, 54, 3410–3416. [Google Scholar] [CrossRef] [Green Version]
- Thomas, R.; Melton, R.; Asbell, P. Antibiotic resistance among ocular pathogens: Current trends from the ARMOR surveillance study (2009–2016). Clin. Optomet. 2019, 11, 15–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franca, A.; Gaio, V.; Lopes, N.; Melo, L. Virulence Factors in Coagulase-Negative Staphylococci. Pathogens 2021, 10, 170. [Google Scholar] [CrossRef]
- Argemi, X.; Hansmann, Y.; Prola, K.; Prevost, G. Coagulase-Negative Staphylococci Pathogenomics. Int. J. Mol. Sci. 2019, 20, 1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, M. Coagulase-negative staphylococcus reservoirs of genes facilitating MRSA infection. Bioessays 2012, 35, 4–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Monk, I.R.; Goncalves da Silva, A.; Seemann, T.; Chua, K.; Kearns, A.; Hill, R.; Woodford, N.; Bartels, M.D.; Strommernger, B.; et al. Global spread of three multidrug-resistant lineages of Staphylococcus epidermidis. Nat. Microbiol. 2018, 3, 1175–1185. [Google Scholar] [CrossRef]
- Sievert, D.; Ricks, P.; Edwards, J. Antimicrobial-Resistance Pathogens Associated with Healthcare-Associated Infections: Summary of Data Reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009–2010. Infect. Control Hosp. Epidemiol. 2013, 34, 188–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayness, N.; Obeid, A.; Storey, P.; Juliano, J.; Rahimy, E.; Moshfeghi, A.; Garg, S.; Hsu, J. Long-Term Visual Outcomes and Clinical Features After Anti-Vascular Endothelial Growth Factor Injection-Related Endophthalmitis. Retina 2019, 39, 2070–2076. [Google Scholar] [CrossRef] [PubMed]
- Alexandrakis, G.; Eduardo, M.; Alfonso, M.; Miller, D. Shifting Trends in Bacterial Keratitis in South Florida and Emerging Resistance to Fluoroquinolones. Ophthalmology 2000, 107, 1497–1502. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, F.; Bruttin, O.; Zografos, L.; Guex-Crosier, Y. Bacterial keratitis: A prospective clinical and microbiological study. Br. J. Ophthalmol. 2001, 85, 842–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leck, A.; Thomas, P.; Hagan, M.; Kaliamurthy, J.; Ackuaku, E.; John, M.; Newman, M.; Codjoe, F.; Opintan, J.; Kalavathy, C.; et al. Aetiology of suppurative corneal ulcers in Ghana and south India, and epidemiology of fungal keratitis. Br. J. Ophthalmol. 2002, 86, 1211–1215. [Google Scholar] [CrossRef] [PubMed]
- Lam, D.; Houang, E.; Fan, D.; Lyon, D.; Seal, D.; Wong, E.; Hong Kong Microbial Keratitis Study Group. Incidence and risk factors for microbial keratitis in Hong Kong: Comparison with Europe and North America. Eye 2002, 16, 608–618. [Google Scholar] [CrossRef]
- Zhang, C.; Liang, Y.; Deng, S.; Wang, Z.; Li, R.; Sun, X. Distribution of bacterial keratitis and emerging resistance to antibiotics in China from 2001 to 2004. Clin. Ophthalmol. 2008, 2, 575–579. [Google Scholar]
- Geethankumari, P.V.; Remya, R.; Girijadevi, M.S.; Reena, A. Bacterial Keratitis and Fungal Keratitis in South Kerala: A Comparative Study. KJO 2011, 23, 43–46. [Google Scholar]
- Orlans, H.O.; Hornby, S.J.; Bowler, I. In vitro antibiotic susceptibility patterns of bacterial keratitis isolates in Oxford, UK: A 10-year review. Eye 2011, 25, 489–493. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.; Walkden, A.; Au, L.; Fullwood, C.; Hamilton, A.; Qamruddin, A.; Armstrong, M.; Brahma, A.; Carley, F. Twelve-year analysis of microbial keratitis trends at a UK tertiary hospital. Eye 2017, 31, 1229–1236. [Google Scholar] [CrossRef]
- Lin, C.; Prajna, L.; Srinivasan, M.; Prajna, N.; McLeod, S.; Acharya, N.; Lietman, T.; Porco, T. Seasonal trends of microbial keratitis in south India. Cornea 2012, 31, 1123–1127. [Google Scholar] [CrossRef] [Green Version]
- Ting, D.; Settle, C.; Morgan, S.; Baylis, O.; Ghosh, S. A 10-year analysis of microbiological profiles of microbial keratitis: The North East England Study. Eye 2018, 32, 1416–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, C.; Sun, C.; Yeh, L.; Ma, D.H.; Chen, P.Y.; Lin, H.; Tan, H.; Chen, H.; Chen, S.; Huang, Y. Shifting Trends in Bacterial Keratitis in Taiwan: A 10-Year Review in a Tertiary-Care Hospital. Cornea 2016, 35, 313–317. [Google Scholar] [CrossRef]
- Gautam, P.; Dahal, P.; Patel, S.; Singh, S. Epidemiology of Bacterial Keratitis in Lumbini Eye Institute. J. Chitwon Med. Coll. 2019, 9, 69–73. [Google Scholar] [CrossRef]
- Huebner, J.; Goldmann, D. Coagulase-Negative Staphylococci: Role as Pathogens. Annu. Rev. Med. 1999, 50, 223–236. [Google Scholar] [CrossRef]
- Schoenfelder, S.; Lange, C.; Eckart, M.; Hennig, S.; Kozytska, S.; Ziebuhr, W. Success through diversity—How Staphylococcus epidermidis establishes as a nosocomial pathogen. Internat. J. Med. Microbiol. 2010, 300, 380–386. [Google Scholar] [CrossRef]
- Kleck, J.; Donahue, J. Production of Thermostable Hemolysin by Cultures of Staphylococcus epidermidis. J. Infect. Dis. 1968, 317, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Okee, M.; Joloba, M.; Okello, M.; Najjuka, F.; Katabazi, F.; Bwanga, F.; Nanteza, A.; Kateete, D. Prevalence of virulence determinants in Staphylococcus epidermidis from ICU patients in Kampala, Uganda. J. Infect. Dev. Ctries. 2012, 6, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-Negative Staphylococci. Clin. Microbiol. Rev. 2014, 27, 870–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moraveji, Z.; Tabatabaei, M.; Shirzad, A.; Khoshbakht, R. Characterization of hemolysins of Staphylococcus strains isolated from human and bovine, southern Iran. Iran. J. Vet. Res. 2014, 15, 326–330. [Google Scholar] [PubMed]
- Nasaj, M.; Saeidi, Z.; Asghari, B.; Roshanaei, G.; Arabestani, M. Identification of hemolysin encoding genes and their association with antimicrobial resistance pattern amoung clinical isolates of coagulase-negative Staphylococci. BMC Res. Notes 2020, 13, 68. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, L.; Brito, C.; de Oliveria, A.; Martins, P.; Pereira, V.; da Cunha, M. Staphylococcus epidermidis and Staphylococcus haemolyticus: Molecular Detection of Cytotoxin and Enterotoxin Genes. Toxins 2015, 7, 3688–3699. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Guo, L.; Li, J.; Li, J.; Cui, L.; Dong, J.; Meng, X.; Qian, C.; Wang, H. Antibiotic resistance, biofilm formation, and virulence factors of isolates of Staphylococcus pseudintermedius from healthy dogs and dogs with keratitis. Front. Vet. Sci. 2022, 9, 903633. [Google Scholar] [CrossRef]
- Ruiz-Ripa, L.; Simon, C.; Ceballos, S.; Ortega, C.; Zarazaga, M.; Torres, C.; Gomez-Sanz, E. S. pseudintermedius and S. aureus lineages with transmission ability circulate as causative agents of infections in pets for years. BMC Vet. Res. 2021, 17, 42. [Google Scholar] [CrossRef] [PubMed]
- Somyaji, R.; Priyantha, M.A.R.; Rubin, J.E.; Church, D. Human infections due to Staphylococcus pseudintermedius, an emerging zoonosis of canine origin: Report of 24 cases. Diagn. Microbiol. Infect. Dis. 2016, 85, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, J.S.; Allbaugh, R.A.; Kenne, D.E.; Sebbag, L. Prevalence and Antibiotic Susceptibility of Bacterial Isolates from Dogs with Ulcerative Keratitis in Midwestern United States. Front. Vet. Sci. 2020, 7, 583965. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.T.; Amador, S.; McGonagle, C.J.; Needle, D.; Gibson, R.; Andam, C.P. Population genomics of Staphylococcus pseudintermedius in companion animals in the United States. Commun. Biol. 2020, 3, 282. [Google Scholar] [CrossRef]
- Pires dos Santos, T.; Damborg, P.; Moodley, P.; Guardabassi, L. Systematic Review on Global Epidemiology of Methicillin-Resistant Staphylococcus pseudintermedius: Inference of Population Structure from Multilocus Sequence Typing Data. Front. Microbiol. 2016, 7, 1599. [Google Scholar] [CrossRef] [Green Version]
- Maali, Y.; Badiou, C.; Martins-Simoes, P.; Hodille, E.; Bes, M.; Vandenesch, F.; Lina, G.; Diot, A.; Laurent, F.; Trouillet-Assant, S. Understanding the Virulence of Staphylococcus pseudintermedius: A Major Role of Pore-Forming Toxins. Front. Cell. Infect. Microbiol. 2018, 8, 221. [Google Scholar] [CrossRef]
- Whiley, R.; Hardie, J. Streptococcus. In Bergey’s Manual of Systematics of Archaea and Bacteria, Online; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar]
- Huang, Y.; Yang, B.; Li, W. Defining the normal core microbiome of conjunctival microbial communities. Clin. Microbiol. Infect. 2016, 22, 643.e7–643.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, H.; Price, K.; Albert, L.; Dodick, J.; Park, L.; Dominguez-Bello, M. Changes in the Eye Microbiota Associated with Contact Lens Wearing. mBio 2016, 7, e00198-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, M.; Norcross, E.; Robertson, Z.; Moore, Q.; Fratkin, J.; Marquart, M. The Streptococcus pneumoniae Capsule Is Required for Full Virulence in Pneumococcal Endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2011, 52, 865–872. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Chong, Y.; Sun, M.; Chen, H.; Liu, L.; Chen, Y.; Wu, W.; Kang, Y.; Lai, C. Streptococcus pneumoniae endophthalmitis: Clinical settings, antibiotic susceptibility, and visual outcomes. Sci. Rep. 2021, 11, 6195. [Google Scholar] [CrossRef]
- Bharathi, M.; Ramakrishnan, R.; Meenakshi, R.; Padmavathy, S.; Shivakumar, C.; Srivinasan, M. Microbial Keratitis in South India: Influence of Risk Factors, Climate, and Geographical Variation. Ophthal. Epidemiol. 2009, 14, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Hyams, C.; Camberlein, E.; Cohen, J.; Bax, K.; Brown, J. The Streptococcus pneumoniae Capsule Inhibits Complement Activity and Neutrophil Phagocytosis by Multiple Mechanisms. Infect. Immun. 2010, 78, 704–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, J.; O’Callaghan, R.; Girgis, D.; McCormick, C.; Caballero, A.; Marquart, M. Ocular Virulence of Capsule-Deficient Streptococcus pneumoniae in a Rabbit Keratitis Model. Investig. Ophthalmol. Vis. Sci. 2005, 46, 604–608. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.; Hobden, J.; Hagenah, M.; O’Callaghan, R.; Hill, J.; Chen, S. The Role of Pneumonolysin in Ocular Infections with Streptococcus pneumomiae. Curr. Eye Res. 1990, 9, 1107–1114. [Google Scholar] [CrossRef]
- Schmelter, V.; Schneider, F.; Priglinger, S.; Liegl, R.; Kreutzer, T.; Foerster, P. Pars-plana-vitrectomy for endophthalmitis treatment and the role of standardized ultrasound. Int. Ophthalmol. 2023, 43, 1111–1119. [Google Scholar] [CrossRef]
- Gower, E.; Keay, L.; Stare, D.; Arora, P.; Cassard, S.; Behrens, A.; Tielsch, J.; Schein, O. Characteristics of Endophthalmitis after Cataract Surgery in the United States Medicare Population. Ophthalmology 2015, 122, 1625–1632. [Google Scholar] [CrossRef] [Green Version]
- Valentino, M.; McGuire, A.; Rosch, J.; Bispo, P.; Burnham, C.; Sanfilippo, C.; Carter, R.; Zegans, M.; Beall, B.; Earl, A.; et al. Unencapsulated Streptococcus pneumoniae from conjunctivitis encode variant traits and belong to a distinct phylogenetic cluster. Nat. Commun. 2014, 5, 5411. [Google Scholar] [CrossRef] [Green Version]
- Andre, C.; Rouhana, J.; Scarpa de Mello, S.; Rosa da Cunha, G.; Van Camp, A.; Gilmore, M.; Bispo, P. Population structure of ocular Streptococcus pneumoniae is highly diverse and formed by lineages that escape current vaccines. Microb. Genom. 2022, 8, 000763. [Google Scholar] [CrossRef] [PubMed]
- Barragan, E.; Perez, J.; Corbella, L.; Orellana, M.; Fernandez-Ruiz, M. Achromobacter xylosoxidans bacteremia: Clinical and microbiological features in a 10-year case series. Rev. Esp. Quimioter 2018, 31, 268–273. [Google Scholar]
- Paplia, M.; Figueroa-Espinosa, R.; Steffanowski, C.; Barberis, C.; Almuzara, M.; Barrios, R.; Vay, C.; Gutkind, G.; Di Conza, J.; Radice, M. Expansion and improvement of MALDI-TOF MS databases for accurate identification of Achromobacter species. J. Microbiol. Meth. 2020, 172, 105889. [Google Scholar] [CrossRef] [PubMed]
- Amoureux, L.; Sauge, J.; Sarret, B.; Lhoumeau, M.; Bajard, A.; Tetu, J.; Bador, J.; Neuwirth, C.; MucoMicrobes group. Study of 109 Achromobacter spp. isolates from 9 French CF centres reveals the circulation of a multiresistant clone of A. xylosoxidans belonging to ST 137. J. Cyst. Fibros. 2019, 18, 804–807. [Google Scholar] [CrossRef] [PubMed]
- Marion-Sanchez, K.; Pailla, K.; Olive, C.; Le Coutour, X.; Derancourt, C. Achromobacter spp. healthcare associated infections in the French West Indies: A longitudinal study from 2006–2016. BMC Infect. Dis. 2019, 19, 795. [Google Scholar] [CrossRef] [PubMed]
- Isler, B.; Kidd, T.; Stewart, A.; Harris, P.; Paterson, D. Achromobacter Infections and Treatment Options. Antimicrob. Agents Chemother. 2020, 64, e01025-20. [Google Scholar] [CrossRef]
- Gabrielaite, M.; Bartell, J.; Norskov-Lauritsen, N.; Pressler, T.; Nielsen, F.; Johansen, M.R. Transmission and Antibiotic Resistance of Achromobacter in Cystic Fibrosis. J. Clin. Microbiol. 2021, 59, e02911-20. [Google Scholar] [CrossRef]
- Arshad, J.; Saud, A.; White, D.; Afshari, N.; Sayegh, R. Chronic Conjunctivitis from a Retained Contact Lens. Eye Contact Lens 2020, 46, e1–e4. [Google Scholar] [CrossRef]
- Reddy, A.; Garg, P.; Shah, V.; Gopinathan, U. Clinical, Microbiological Profile and Treatment Outcome of Ocular Infections Caused by Achromobacter xylosoxidans. Cornea 2009, 28, 1100–1103. [Google Scholar] [CrossRef]
- Lazzarini, T.; Al-khersan, H.; Patel, N.; Yannuzzi, N.; Martinez, J.; Altamirano, D.; Torres, L.; Miller, D.; Batlle, J.; Amescua, G.; et al. Chronic, Recurrent Bacterial Endophthalmitis Caused by Achromobacter xylosoxidans: Clinical Features and Management. Int. Med. Case Rep. J. 2020, 13, 265–269. [Google Scholar] [CrossRef]
- Bergogne-Berezin, E.; Tower, K. Acinetobacter spp. as Nosocomial Pathogens: Microbiological, Clinical, and Epidemiological Features. Clin. Microbiol. Rev. 1996, 9, 148–165. [Google Scholar] [CrossRef]
- Harding, C.; Hennon, S.; Feldman, M. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Microbiol. 2018, 16, 91–102. [Google Scholar] [CrossRef]
- Talreja, D.; Muraleedharan, C.; Gunathilaka, G.; Zhang, Y.; Kaye, K.; Walia, S.; Kumar, A. Virulence Properties of Multidrug Resistant Ocular Isolates of Acinetobacter baumannii. Curr. Eye Res. 2014, 39, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Mark, D.; Gaynon, M. Trauma-induced endophthalmitis caused by Acinetobacter anitratus. Br. J. Ophthalmol. 1983, 67, 124–126. [Google Scholar] [CrossRef]
- Crawford, P.; Conway, M.; Peyman, G. Trauma-Induced Acinetobacter iwoffi Endophthalmitis with Multi-Organism Recurrence: Strategies with Intravitreal Treatment. Eye 1997, 11, 863–864. [Google Scholar] [CrossRef] [PubMed]
- Broniek, G.; Langwinska-Wosko, E.; Szaflik, J.; Wroblewska, M. Acinetobacter junii as an aetiological agent of corneal ulcer. Infection 2014, 42, 1051–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, C.; Yin, Z.; Wang, J.; Qian, C.; Wei, Y.; Zhang, S.; Jiang, L.; Liu, B. Comparative Genomic Analysis of Citrobacter and Key Genes Essential for the Pathogenicity of Citrobacter koseri. Front. Microbiol. 2019, 10, 2774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaretti, A.; Righini, L.; Candeliere, F.; Musmeci, E.; Bonvicini, F.; Gentilomi, G.; Rossi, M.; Raimondi, S. Antibiotic Resistance, Virulence Factors, Phenotyping, and Genotyping of Non-Escherichia coli Enterobacterales from the Gut Microbiota of Healthy Subjects. Int. J. Mol. Sci. 2020, 21, 1847. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.; Liu, C.; Tong, J.; Luk, W.; Li, K. Citrobacter Endogenous Endophthalmitis: A Case Report and Review of the Literature. Retin. Cases Br. Rep. 2020, 14, 187–191. [Google Scholar] [CrossRef]
- Chen, K.; Sun, M.; Hwang, Y.; Chen, T.; Lai, C.; Chen, Y. Endophthalmitis Caused by Citrobacter Species. Ocul. Immunol. Inflam. 2008, 16, 147–153. [Google Scholar] [CrossRef]
- Goold, L.A.; Warrier, S.K.; Wittles, N.K.; Nathan, F. Microbial Keratitis Secondary to Infection with Citrobacter koseri. Cornea 2010, 29, 479. [Google Scholar] [CrossRef]
- Khanam, Z.; Gujral, G.; Khan, S. Infectious crystalline keratitis induced by Citrobacter. GMS Ophthalmol. Cases 2021, 11, Doc09. [Google Scholar] [PubMed]
- Balamurugan, R.; Gupta, P.; Raghulnadhan, R.; Angrup, A.; Yadav, B.; Ram, J. Microbial keratitis due to infection with Citrobacter freundii: A rare entity. Indian J. Ophthalmol. Case Rep. 2021, 1, 93. [Google Scholar]
- Ramirez, D.; Giron, M. Enterobacter Infections; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Mishra, M.; Panda, S.; Barik, S.; Sarkar, A.; Singh, D.; Mohapatra, H. Antibiotic Resistance Profile, Outer Membrane Proteins, Virulence Factors and Genomic Sequence Analysis Reveal Clinical Isolates of Enterobacter Are Potential Pathogens Compared to Environmental Isolates. Front. Cell. Infect. Microbiol. 2020, 10, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okhravi, N.; Ficker, L.; Matheson, M.M.; Lightman, S. Enterobacter cloacae endophthalmitis: Report of four cases. J. Clin. Microbiol. 1998, 36, 48–51. [Google Scholar] [CrossRef] [Green Version]
- Boldt, H.C.; Pulido, J.S.; Blodi, C.F.; Folk, J.C.; Weingeist, T.A. Rural endophthalmitis. Ophthalmology 1989, 12, 1722–1726. [Google Scholar] [CrossRef]
- Puliafito, C.A.; Baker, A.S.; Haaf, J.; Foster, C.S. Infectious endophthalmitis. Review of 36 cases. Ophthalmology 1982, 89, 921–929. [Google Scholar] [CrossRef]
- Dave, V.; Pathengay, A.; Behera, S.; Joseph, J.; Sharma, S.; Pappuru, R.; Das, T. Enterobacter endophthalmitis: Clinical settings, susceptibility profile, and management outcomes across two decades. Indian J. Ophthalmol. 2020, 68, 112–116. [Google Scholar] [CrossRef]
- Pathengay, A.; Trehan, H.; Mathai, A.; Jalali, S.; Majji, A.; Das, M.; Sharma, S.; Das, T. Enterobacter Endophthalmitis Clinicomicrobiologic Profile and Outcomes. Retina 2012, 32, 558–562. [Google Scholar] [CrossRef]
- Blount, Z.D. The unexhausted potential of E. coli. eLife 2015, 4, e05826. [Google Scholar] [CrossRef]
- Denamur, E.; Clermont, O.; Bonacorsi, S.; Gordon, D. The population genetics of pathogenic Escherichia coli. Nat. Rev. Microbiol. 2021, 19, 37–54. [Google Scholar] [CrossRef]
- Sarowsha, J.; Futoma-Kolock, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-Ploskonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjith, K.; Sharma, S.; Shivaji, S. Microbes of the human eye: Microbiome, antimicrobial resistance and biofilm formation. Exp. Eye Res. 2021, 205, 108476. [Google Scholar] [CrossRef] [PubMed]
- Zhai, H.; Bispo, P.; Kobashi, H.; Jacobs, D.; Gilmore, M.; Ciolino, J. Resolution of fluoroquinolone-resistant Escherichia coli keratitis with a PROSE device for enhanced targeted antibiotic delivery. Am. J. Ophthalomol. Case Rep. 2018, 12, 73–75. [Google Scholar] [CrossRef]
- Ranjith, K.; Abhilash, C.; Prashanthi, G.; Padakandla, S.; Sharma, S.; Shivaji, S. Phylogenetic Grouping of Human Ocular Escherichia coli based on Whole-Genome Sequence Analysis. Microorganisms 2020, 8, 422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, T.; Paraskevopoulos, T.; Georgalas, I. Systematic review of 342 cases of endogenous bacterial endophthalmitis. Surv. Ophthalmol. 2014, 59, 627–635. [Google Scholar] [CrossRef]
- Mihancea, N. Haemophilus infections in humans. Rom. Arch. Microbiol. Immunol. 1993, 52, 5–13. [Google Scholar]
- Kilian, M. Haemophilus. In Bergey’s Manual of Systematics of Archaea and Bacteria, Online; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar]
- Connor, J.D.; Nyhan, W.L. Haemophilus influenzae infections. N. Engl. J. Med. 1972, 286, 107–108. [Google Scholar]
- Wen, S.; Feng, D.; Chen, D.; Yang, L.; Xu, Z. Molecular epidemiology and evolution of Haemophilus influenzae. Infect. Genet. Evolut. 2020, 80, 1–6. [Google Scholar] [CrossRef]
- Leung, A.K.C.; Hon, K.L.; Wong, A.H.C.; Wong, A.S. Bacterial Conjunctivitis in Childhood: Etiology, Clinical Manifestations, Diagnosis, and Management. Recent Pat. Inflamm. Allergy Drug Discov. 2018, 12, 120–127. [Google Scholar] [CrossRef]
- Jin, Z.; Romero-Steiner, S.; Carlone, G.M.; Robbins, J.B.; Schneerson, R. Haemophilus influenzae type a infection and its prevention. Infect Immun. 2007, 75, 2650–2654. [Google Scholar] [CrossRef] [Green Version]
- Podschun, R.; Ullmann, U. Klebsiella spp. as nosocomial pathogens: Epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magill, S.; Edwards, J.; Stat, M.; Bamberg, W.; Beldavs, Z.G.; Dumyati, G.; Kainer, M.A.; Lynfield, R.; Maloney, M.; McAllister-Hollod, L.; et al. Multistate Point-Prevalence survey of Health Care-Associated Infections. N. Engl. J. Med. 2014, 370, 1198–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengoechea, J.; Sa Pessoa, J. Klebsiella pneumoniae infection biology: Living to counteract host defenses. FEMS Microbiol. Rev. 2018, 43, 123–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bratu, S.; Landman, D.; Haag, R.; Recco, R.; Eramo, A. Rapid spread of carbapenem-resistant Klebsiella pneumoniae in New York City: A new threat to our antibiotic armamentarium. Arch. Intern. Med. 2005, 165, 1430–1435. [Google Scholar] [CrossRef] [Green Version]
- Mazzariol, A.; Roelofsen, E.; Koncan, R.; Voss, A.; Cornaglia, G. Detection of a new SHV-Type extended-spectrum ß-lactamase, SHV-31, in a Klebsiella pneumoniae strain causing a large nosocomial outbreak in the Netherlands. Antimicrol. Agents Chemother. 2007, 51, 1082–1084. [Google Scholar] [CrossRef]
- Snitkin, E.S.; Zelazny, A.M.; Thomas, P.J.; Stock, F.; NISC Comparative Sequencing Program Group; Henderson, D.K.; Palmore, T.N.; Segre, J.A. Tracking a hospital outbreak of carbapenem-resistant Klebsiella pneumoniae with whole-genome sequencing. Sci. Transl. Med. 2012, 4, 148ra116. [Google Scholar] [CrossRef] [Green Version]
- Favre-Bonté, S.; Licht, T.R.; Forestier, C.; Krogfelt, K.A. Klebsiella pneumoniae capsule expression is necessary for colonization of large intestines of streptomycin-treated mice. Infect. Immun. 1999, 67, 6152–6156. [Google Scholar] [CrossRef] [Green Version]
- Pomakova, D.K.; Hsiao, C.-B.; Beanan, J.M.; Olson, R.; Macdonald, U.; Keynan, Y.; Russo, T.A. Clinical and phenotypic differences between classic and hypervirulent Klebsiella pneumonia: An emerging and under-recognized pathogenic variant. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 981–989. [Google Scholar] [CrossRef]
- Wiskur, B.; Hunt, J.; Callegan, M. Hypermucoviscosity as a virulence factor in experimental Klebsiella pneumoniae endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4931–4938. [Google Scholar] [CrossRef] [Green Version]
- Hunt, J.; Wang, J.; Callegan, M. Contribution of mucoviscosity associated gene A (magA) to virulence in experimental Klebsiella pneumoniae endophthalmitis. Investig. Opthalmol. Vis. Sci. 2011, 52, 6860–6866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, T.; Eykyn, S.; Graham, E.; Stanford, M. Endogenous bacterial endophthalmitis: A 17-year prospective series and review of 267 reported cases. Surv. Ophthalmol. 2003, 48, 403–423. [Google Scholar] [CrossRef] [PubMed]
- Mak, C.; Ho, M.; Iu, L.; Sin, H.; Chen, L.; Lui, G.; Brelen, M.; Young, A. Clinical features and treatment outcomes of endogenous Klebsiella endophthalmitis: A 12-year review. Int. J. Ophthalmol. 2020, 13, 1933–1940. [Google Scholar] [CrossRef] [PubMed]
- Coburn, P.S.; Wiskur, B.J.; Christy, E.; Callegan, M.C. The Diabetic Ocular Enviroment Facilitates the Development of Endogenous Bacterial Endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7426–7431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coburn, P.S.; Wiskur, B.J.; Astley, R.A.; Callegan, M.C. Blood-Retinal Barrier Compromise and Endogenous Staphylococcal aureus Endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7303–7311. [Google Scholar] [CrossRef] [Green Version]
- Dago, T.; Zewudie, A.; Mamo, Y.; Feyissa, D.; Geleta, S. Multi-Drug Resistant Post Corneal Repair Klebsiella oxytoca Keratitis. Intern. Med. Case Rep. J. 2020, 13, 537–541. [Google Scholar]
- Gao, Y.; Li, C.; Bu, P.; Zhang, L.; Bouchard, C. Infectious interface keratitis (IIK) following keratoplasty; A literature review. Ocul. Surf. 2019, 17, 635–643. [Google Scholar] [CrossRef]
- Morax, V. Note sur un diplobacilli pathogen pour la conjonctive humain. Ann. Inst. Pasteur 1896, 10, 337–345. [Google Scholar]
- Axenfeld, T. Uber die Chronishe Diplobacillenconjunctivitis. Zentralbl. Bakteriol. Abt. 1897, 21, 1–9. [Google Scholar]
- LaCroce, S.; Wilson, M.; Romanowski, J.; Newman, J.; Jhanji, V.; Shanks, R.; Kowalski, R. Moraxella nonliquefaciens and M. osloensis Are Important Moraxella Species That Cause Ocular Infections. Microorganisms 2019, 7, 163. [Google Scholar] [CrossRef] [Green Version]
- Durrani, A.; Faith, S.; Kowalski, R.; Yu, M.; Romanowski, E.; Shanks, R.; Dhaliwal, D.; Jhanji, V. Moraxella Keratitis: Analysis of Risk Factors, Clinical Characteristics, Management, and Treatment Outcomes. Am. J. Ophthalmol. 2018, 197, 17–22. [Google Scholar] [CrossRef] [PubMed]
- McSwiney, T.; Knowles, S.; Murphy, C. Clinical and microbiological characteristics of Moraxella keratitis. Br. J. Ophthalmol. 2019, 103, 1704–1709. [Google Scholar] [CrossRef]
- Das, S.; Constantinou, M.; Daniell, M.; Taylor, H. Moraxella keratitis: Predisposing factors and clinical review of 95 cases. Br. J. Ophthalmol. 2006, 90, 1236–1238. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, B.; Harrison, L.; Motter, J.; Motter, R.; Hightower, M.; Broome, C. Investigation of an Outbreak of Moraxella Conjunctivitis at a Navajo Boarding School. Am. J. Ophthalmol. 1989, 107, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Hoarau, G.; Merabet, L.; Brignole-Baudouin, F.; Mizrahi, A.; Borderie, V.; Bouheraoua, N. Moraxella keratitis: Epidemiology and outcomes. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 2317–2325. [Google Scholar] [CrossRef]
- Blakeway, L.; Tan, A.; Peak, I.; Seib, K. Virulence determinates of Moraxella catarrhalis: Distribution and considerations for vaccine development. Microbiology 2017, 163, 1371–1384. [Google Scholar] [CrossRef]
- Zafar, H.; Tan, S.; Walkden, A.; Fullwood, C.; Au, L.; Brahma, A.; Carley, F. Clinical Characteristics and Outcomes of Moraxella Keratitis. Cornea 2018, 37, 1551–1554. [Google Scholar] [CrossRef]
- Pachori, P.; Gothalwal, R.; Gandhi, P. Emergence of antibiotic resistance Pseudomonas aeruginosa in intensive care unit; a critical review. Genes Dis. 2019, 6, 109–119. [Google Scholar] [CrossRef]
- Pandey, C.; Kumari, S.; Tiwari, R. To determine the bacteriological profile of acute conjunctivitis in cases in NMCH. Eur. J. Mol. Clin. Med. 2020, 7, 3345–3350. [Google Scholar]
- Chen, K.; Sun, M.; Lai, C.; Wu, W.; Chen, T.; Kuo, Y.; Chao, A.; Hwang, Y.; Chen, Y.; Wang, N.; et al. Endophthalmitis Caused by Pseudomonas aeruginosa in Taiwan. Retina 2011, 31, 1193–1198. [Google Scholar] [CrossRef]
- Sridhar, J.; Kuriyan, A.; Flynn, H.; Miller, D. Endophthalmitis Caused by Pseudomonas aeruginosa Clinical Features, Antibiotic Susceptibilities, and Treatment Outcomes. Retina 2015, 35, 1101–1106. [Google Scholar] [CrossRef]
- Reedy, J.; Wood, K. Endogenous Pseudomonas aeruginosa endophthalmitis: A case report and literature review. Inten. Care Med. 2000, 26, 1386–1389. [Google Scholar] [CrossRef]
- Willcox, M. Pseudomonas aeruginosa Infection and Inflammation During Contact Lens Wear: A Review. Optom. Vis. Sci. 2007, 84, 273–278. [Google Scholar] [CrossRef]
- Rossetto, J.; Cavuoto, K.; Osigian, C.; Chang, T.; Miller, D.; Capo, H.; Spierer, O. Paediatric infectious keratitis: A case series of 107 children presenting to a tertiary referral centre. Br. J. Ophthalmol. 2017, 101, 1488–1492. [Google Scholar] [CrossRef] [PubMed]
- Hilliam, Y.; Kaye, S.; Winstanley, C. Pseudomonas aeruginosa and microbial keratitis. J. Med. Microbiol. 2020, 69, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Available online: cdc.gov/hai/outbreaks/crpa-artificial-tears.html#print (accessed on 30 March 2023).
- Montgomery, M.L.; Callegan, M.C.; Fuller, K.K.; Carr, D.J.J. Ocular Glands Become Infected Secondarily to Infectious Keratitis and Play a Role in Corneal Resistance to Infection. J. Virol. 2019, 93, e00314-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shankar, J.; Sueke, H.; Wiehlmann, L.; Horsburgh, M.J.; Tuft, S.; Neal, T.J.; Baye, S.B.; Winstanley, C. Genotypic analysis of UK keratitis-associated Pseudomonas aeruginosa suggests adaptation to environmental water as a key component in the development of eye infections. FEMS Microbiol. Lett. 2012, 334, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thibodeaux, B.A.; Caballero, A.R.; Marquart, M.E.; Tommassen, J.; O’Callaghan, R.J. Corneal Virulence of Pseudomonas aeruginosa Elastase B. and Alkaline Protease Produced by Pseudomonas putida. Curr. Eye Res. 2007, 32, 373–386. [Google Scholar] [CrossRef]
- O’Callaghan, R.; Caballero, A.; Tang, A.; Bierdeman, M. Pseudomonas aeruginosa keratitis: Protease IV and PASP as Corneal Virulence Mediators. Microorganisms 2019, 7, 281. [Google Scholar] [CrossRef] [Green Version]
- Engel, L.S.; Hobden, J.A.; Moreau, J.M.; Callegan, M.C.; Hill, J.M.; O’Callaghan, R.J. Pseudomonas deficient in protease IV has significantly reduced corneal virulence. Investig. Ophthalmol. Vis. Sci. 1997, 38, 1535–1542. [Google Scholar]
- Zuberbuhler, B.; Carifi, G. Pseudomonas putida infection of the conjunctiva. Infection 2012, 40, 579–580. [Google Scholar] [CrossRef]
- Deshmukh, D.; Joseph, J.; Chakrabarti, M.; Sharma, S.; Jayasudha, R.; Sama, K.; Sontam, B.; Tyagi, M.; Narayanan, R.; Shivaji, S. New insights into culture negative endophthalmitis by unbiased next generation sequencing. Sci. Rep. 2019, 9, 844. [Google Scholar] [CrossRef] [Green Version]
- Fleiszig, S.; Evans, D. The pathogenesis of bacterial keratitis: Studies with Pseudomonas aeruginosa. Elin. Exp. Optom. 2002, 85, 271–278. [Google Scholar] [CrossRef]
- Evans, E.; Fleiszig, S. Why Does the Healthy Cornea Resist Pseudomonas aeruginosa Infection? Am. J. Ophthalmol. 2013, 155, 961–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleiszig, S.; Kroken, A.; Nieto, V.; Grosser, M.; Wan, S.; Metruccio, M.; Evans, D. Contact lens-related corneal infection: Intrinsic resistance and its compromise. Prog. Ret. Eye Res. 2020, 76, 100804. [Google Scholar] [CrossRef] [PubMed]
- Subedi, D.; Vijay, A.; Willcox, M. Overview of mechanisms of antibiotic resistance in Pseudomonas aeruginosa: An ocular perspective. Clin. Exp. Optom. 2018, 101, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Soleimani, M.; Tabatabaei, S.; Masoumi, A.; Mirshahi, R.; Ghahvechian, H.; Tayebi, F.; Momenaei, B.; Mahdizad, Z.; Mohammadi, S. Infectious keratitis: Trends in microbiological and antibiotic sensitivity patterns. Eye 2021, 35, 3110–3115. [Google Scholar] [CrossRef]
- Armbruster, C.; Mobley, H.; Pearson, M. Pathogenesis of Proteus mirabilis Infection. EcoSal Plus 2018, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, S.; Durrani, A.; Safiullah, Z.; Kowalski, R.; Jhanji, V. Proteus mirabilis Keratitis: Risk Factors, Clinical Features, Treatment Outcomes, and Microbiological Characteristics. Cornea 2021, 40, 704–709. [Google Scholar] [CrossRef]
- Lam, D.; Kwok, A.; Chew, S. Post-keratoplasty endophthalmitis caused by Proteus mirabilis. Eye 1998, 12, 139–140. [Google Scholar] [CrossRef]
- Khanna, A.; Khanna, M.; Aggarwal, A. Serratia marcescens—A rare opportunistic nosocomial pathogen and measures to limit its spread in hospitalized patients. J. Clin. Diagn. Res. 2013, 7, 243–246. [Google Scholar]
- Grimont, F.; Grimont, P. The Genus Serratia. In The Prokaryotes: A Handbook on the Biology of Bacteria Volume 6: Proteobacteria: Gamma Subclass; Springer: New York, NY, USA, 2006; pp. 219–244. [Google Scholar]
- Mahlen, S. Serratia infections: From military experiments to current practice. Clin. Microbiol. Rev. 2011, 24, 755–791. [Google Scholar] [CrossRef] [Green Version]
- Shanks, R.; Stella, N.; Hunt, K.; Brothers, K.; Zhang, L.; Thibodeau, P. Identification of SlpB, a Cytotoxic Protease from Serratia marcescens. Infect. Immun. 2015, 83, 2907–2916. [Google Scholar] [CrossRef] [Green Version]
- Vafina, G.; Zainutdinova, E.; Bulatov, E.; Filimonova, M.N. Endonuclease from Gram-Negative Bacteria Serratia marcescens Is as Effective as Pulmozyme in the Hydrolysis of DNA in Sputum. Front. Pharmacol. 2018, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Zhang, R.; Sun, Y.; Platt, S.; Szczotka-Flynn, L.; Pearlman, E. Innate Immune Regulation of Serratia marcescens—Induced Corneal Inflammation and Infection. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7382–7388. [Google Scholar] [CrossRef] [Green Version]
- Romanowski, E.; Stella, N.; Romanowski, J.; Yates, K.; Dhaliwal, D.; St. Leger, A.; Shanks, R. The Rcs Stress Response System Regulator GumB Modulates Serratia marcescens-Induced Inflammation and Bacterial Proliferation in a Rabbit Keratitis Model and Cytotoxicity In Vitro. Infect. Immun. 2021, 89, e00111-21. [Google Scholar] [CrossRef] [PubMed]
- Hejazi, A.; Falkiner, F. Serratia marcescens. J. Med. Microbiol. 1997, 46, 903–912. [Google Scholar] [CrossRef] [Green Version]
- Atta, S.; Perera, C.; Nayyar, S.; Kowalski, R.; Jhanji, V. An 18-Year Overview of Serratia marcescens Ocular Infection. Eye Contact Lens 2021, 47, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Ranganatha, A.; Rana, M.; Murray, A. Serratia liquefactions Causing Severe Ocular Damage in Noncontact Lens Wearer. Eye Contact Lens 2018, 44, 320–321. [Google Scholar] [CrossRef] [PubMed]
- Mah-Sadorra, J.; Najjar, D.; Rapuano, J.; Laibson, P.; Cohen, E. Serratia Corneal Ulcers A Retrospective Clinical Study. Cornea 2005, 24, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Mariella, M.A.; Warwar, R. Endogenous endophthalmitis due to Serratia marcescens. South. Med. J. 1998, 91, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Equi, R.; Green, W. Endogenous Serratia marcescens Endophthalmitis with Dark Hypopyon: A Case Report and Review. Surv. Ophthalmol. 2001, 46, 259–268. [Google Scholar] [CrossRef]
- Brooke, J. Advances in the Microbiology of Stenotrophomonas maltophilia. Clin. Microbiol. Rev. 2021, 34, e0003019. [Google Scholar] [CrossRef] [PubMed]
- Das, T.; Deshmukh, H.; Mathai, A.; Reddy, A. Stenotrophomonas maltophilia endogenous endophthalmitis: Clinical presentation, sensitivity spectrum and management. J. Med. Microbiol. 2009, 58, 837–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.; Flynn, H.; Miller, D.; Smiddy, W. Stenotrophomonas maltophilia endophthalmitis following cataract surgery: Clinical and microbiological results. Clin. Ophthalmol. 2013, 7, 771–777. [Google Scholar] [CrossRef] [Green Version]
- Wu, A.; Yeh, L.; Ma, D.; Chen, P.; Lin, H.; Sun, C.; Tan, H.; Chen, H.; Chen, S.; Hsiao, C. Clinical Characteristics of Stenotrophomonas maltophilia Keratitis. Cornea 2016, 16, 795–800. [Google Scholar] [CrossRef]
- Park, B.; Lim, H.; Park, S.; Koh, J. Clinical Features and Management of Stenotrophomonas maltophilia Keratitis. Ophthalmol. Ther. 2021, 10, 525–533. [Google Scholar] [CrossRef]
- Anukolu, R.; Kura, Y.; Kumar, R.; Manderwad, G. Role of Abiotrophia defective in Ocular Infections—A Review. Intern. J. Cell Biol. Cell. Process. 2017, 3, 9–12. [Google Scholar]
- Mehrotra, N.; Baidya, A.; Brijwal, M.; Aggarwal, R.; Chaudhry, R. Actinomycosis of the eye: Forgotten but not uncommon. Anaerobe 2015, 35, 1–2. [Google Scholar] [CrossRef]
- Pandit, R.T. Brevundimonas diminuta Keratitis. Eye Contact Lens 2012, 38, 63–65. [Google Scholar] [CrossRef]
- Daley, D.; Neville, S.; Kociuba, K. Peritonitis Associated with a CDC Group EO-3 Organism. J. Clin. Microbiol. 1997, 35, 3338–3339. [Google Scholar] [CrossRef] [Green Version]
- Forsythe, S.J. Updates on the Cronobacter Genus. Annu. Rev. Food Sci. Technol. 2018, 9, 23–44. [Google Scholar] [CrossRef] [PubMed]
- Strysko, J.; Cope, J.; Martin, H.; Tarr, C.; Hise, K.; Collier, S.; Bowen, A. Food Safety and Invasive Cronobacter Infections during Early Infancy, 1961–2018. Emerg. Infect. Dis. 2020, 26, 857–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duar, R.M.; Lin, X.B.; Zheng, J.; Martino, M.E.; Grenier, T.; Perez-Munoz, M.E.; Leulier, F.; Ganzle, M.; Walter, J. Lifestyles in translocation: Evolution and natural history of the genus Lactobacillus. FEMS Microbiol. Rev. 2017, 41, 27–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kullar, R.; Goldenstein, E.; Johnson, S.; McFarland, L.V. Lactobacillus Bactermia and Probiotics: A Review. Microorganisms 2023, 11, 896. [Google Scholar] [CrossRef]
- Iovieno, A.; Lambiase, A.; Sacchetti, M.; Stampachiacchiere, B.; Micera, A.; Bonini, S. Preliminary evidence for the efficacy of probiotic eye-drop treatment in patients with vernal keratoconjunctivitis. Graefes Arch. Clin. Exp. Ophthalmol. 2007, 246, 435–441. [Google Scholar] [CrossRef]
- Hu, J.; Chen, F.; Kan, T.; Zhuang, H.; Zhang, J.; Han, X. Inhibition of Fusarium solani Infection in Murine Keratocytes by Lactobacillus salivarius ssp. salivarius JC1231 Culture In Vitro. Curr. Eye Res. 2017, 42, 1339–1347. [Google Scholar] [CrossRef]
- Kate, A.; Bagga, B.; Ramazonova, K.; Joshi, V.; Mohamed, A.; Sharma, S. Risk factors, clinical features and outcomes of Neisseria keratitis. Int. Ophthalmol. 2021, 41, 3361–3369. [Google Scholar] [CrossRef] [PubMed]
- Takada, S.; Fujiwara, S.; Inoue, T.; Kataoka, Y.; Hadano, Y.; Matsumoto, K.; Morino, K.; Shimizu, T. Meningococcemia in Adults: A Review of the Literature. Intern. Med. 2016, 55, 567–572. [Google Scholar] [CrossRef] [Green Version]
- Quillin, S.; Seifert, H. Neisseria gonorrhoeae host adaptation and pathogenesis. Nat. Rev. Microbiol. 2018, 16, 226–240. [Google Scholar] [CrossRef] [PubMed]
- Butler, L.; Shah, M.; Cottom, L.; Winter, A.; Lockington, D. Five-year review of ocular Neisseria gonorrhoeae infections presenting to ophthalmology departments in Greater Glasgow & Clyde, Scotland. Eye 2021, 36, 1442–1447. [Google Scholar]
- Kawashima, M.; Kawakita, T.; Den, S.; Tomitia, M.; Shimazaki, J. Surgical management of corneal perforation secondary to gonococcal keratoconjunctivitis. Eye 2009, 23, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Tipple, C.; Smith, A.; Bakowska, E.; Corbett, M. Corneal perforation requiring corneal grafting: A rare complication of gonococcal eye infection. Sex Transm. Infect. 2010, 86, 447–448. [Google Scholar] [CrossRef] [Green Version]
- Center for Disease Control and Prevention. Sexually Transmitted Diseases Treatment Guidelines. 2022. Available online: cdc.gov/std/treatment-guidelines/gonorrhea-adults.htm (accessed on 15 March 2022).
- Mochizuki, Y.; Ishikawa, H.; Sato, A.; Yamada, K.; Takesue, Y.; Gomi, F. Pasteurella multocida-induced endophthalmitis after a cat scratch. Am. J. Ophthalmol. Case Rep. 2020, 18, 100711. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Talati, M.; Mauger, T. Medical and surgical management of Pasteurella canis infectious keratitis. IDCases 2017, 9, 42–44. [Google Scholar] [CrossRef]
- Lion, C.; Conroy, M.; Carpentier, A.; Lozniewski, A. Antimicrobial susceptibilities of Pasteurella strains isolated from humans. Int. J. Antimicrob. Agents 2006, 27, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Koreishi, A.; Schechter, M.; Karp, C. Ocular Infections Caused by Providencia rettgeri. Ophthalmology 2006, 113, 1463–1466. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, C.M.; Brenner, F.W.; Miller, J.M. Classification, Identification, and Clinical Significance of Proteus, Providencia, and Morganella. Clin. Microbiol. Rev. 2000, 12, 534–546. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.; Balshi, A. First literature review of carbapenem-resistant Providencia. New Microbes New Infect. 2018, 25, 16–23. [Google Scholar] [CrossRef]
- Choi, W.; Ji, Y.; Yoon, K. A case of bilateral keratitis caused by Providencia alcalifaciens: A rarely encountered ocular pathogens. Int. Ophthalmol. 2018, 38, 1325–1328. [Google Scholar] [CrossRef]
- Gopaul, D.; Ellis, C.; Maki, A.; Joseph, M. Isolation of Rhodococcus rhodochrous from a Chronic Corneal Ulcer. Diagn. Microbiol. Infect. Dis. 1988, 10, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.L.; Goodfellow, M. Rhodococcus. In Bergey’s Manual of Systematics of Archaea and Bacteria, Online; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar]
- Bell, K.; Philip, J.; Aw, D.; Christofi, N. The genus Rhodococcus. J. Appl. Microbiol. 1998, 85, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.E.; Moore, J.E.; Jiru, X.; Moore, J.E.; Goodall, E.A.; Dooley, J.S.G.; Hayes, V.E.A.; Dartt, D.A.; Downes, C.S.; Moore, T.C.B. Ocular Pathogen or Commensal: A PCR-Based Study of Surface Bacterial Flora in Normal and Dry Eyes. Investig. Ophthalmol. Vis. Sci. 2007, 49, 5616–5623. [Google Scholar] [CrossRef] [PubMed]
- Cuello, O.; Caorlin, J.; Reviglio, V.; Carvajal, L.; Juarez, C.; de Guerra, E.; Luna, J. Rhodococcus globerulus keratitis after laser in situ keratomileusis. J. Cataract. Refract. Surg. 2001, 28, 2235–2237. [Google Scholar] [CrossRef]
- Lalitha, P.; Srinivasan, M.; Prajna, V. Rhodococcus ruber as a Cause of Keratitis. Cornea 2006, 25, 238–239. [Google Scholar] [CrossRef] [PubMed]
- Rummelt, W.; Naumann, K. Chronic postoperative endophthalmitis following cataract extraction and intraocular lens implantation. Report on nine patients. German J. Ophthalmol. 1993, 2, 419–425. [Google Scholar]
- Vosoghi, H.; Lyle, C.E.; Fleming, J.C. Recurrent Rhodococcus gordoniae Eyelid Infection After Combined Septoplasty and Blepharoplasty. Am. Soc. Ophthal. Plastic Recon. Surg. 2010, 26, 127–129. [Google Scholar] [CrossRef]
- Abidi, M.; Ledeboer, N.; Banerjee, A.; Hari, P. Morbidity and mortality attributable to Rothia bacteremia in neutropenic and nonneutropenic patients. Diagn. Microbiol. Infect. Dis. 2016, 85, 116–120. [Google Scholar] [CrossRef]
- Oie, S.; Mochizuki, K.; Ishida, K.; Nakayama, A.; Ohkusu, K. Case of late-onset bleb associated endophthalmitis caused by Rothia mucilaginosa. J. Infect. Chemother. 2016, 22, 645–647. [Google Scholar] [CrossRef]
- Alvarez-Ramos, P.; Del Moral-Ariza, A.; Alonso-Moroto, J.; Marin-Casanova, P.; Calandria-Amigueti, J.; Rodriguez-Inglesias, M.; Rodriguez de la Rua, E. First report of acute postoperative endophthalmitis caused by Rothia mucilaginosa after phacoemulsification. Infect. Dis. Rep. 2016, 8, 6–7. [Google Scholar] [CrossRef] [Green Version]
- Morley, A.; Tuft, S. Rothia dentocariosa Isolated from a Corneal Ulcer. Cornea 2006, 25, 1128–1129. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.; Jafri, M.; Arfeen, S.; Reidy, J.; Hariprasad, S.; Farooq, A. Atypical Keratitis Caused by Rothia Dentocariosa. Ocul. Immunol. Inflam. 2019, 29, 26–28. [Google Scholar] [CrossRef]
- O’Brian, T.; Maguire, M.; Fink, N.; Alfonso, E.; McDonnell, P. Efficacy of Ofloxacin Vs Cefazolin and Tobramycin in the Therapy for Bacterial Keratitis: Report from the Bacterial Keratitis Study Research Group. Arch. Ophthalmol. 1995, 113, 1257–1265. [Google Scholar]
- Ofloxacin Study Group. Ofloxacin Monotherapy for the Primary Treatment of Microbial Keratitis. A Double-masked, Randomized, Controlled Trial with Conventional Dual Therapy. Ophthalmology 1996, 104, 1902–1909. [Google Scholar]
- Low, L.; Nakamichi, K.; Akileswaran, L.; Lee, C.; Lee, A.; Moussa, G.; Murray, P.; Wallace, G.; Van Gelder, R.; Rauz, S.; et al. Deep metagenomic sequencing for endophthalmitis pathogen detection using a nanopore platform. Am. J. Ophthalmol. 2022, 242, 243–251. [Google Scholar] [CrossRef]
- Moshirfar, M.; Hopping, G.; Vaidyanathan, U.; Liu, H.; Somani, A.; Ronquillo, Y.; Hoopes, P. Biological Staining and Culturing in Infectious Keratitis: Controversy in Clinical Utility. Med. Hypothesis Dis. Innov. Ophthalmol. 2019, 8, 145–151. [Google Scholar]
- Park, J.; Lee, K.M.; Zhou, H.; Rabin, M.; Jwo, K.; Burton, W.B.; Gritz, D.C. Community Practice Patterns for Bacterial Corneal Ulcer Evaluation and Treatment. Eye Contact Lens 2015, 41, 12–18. [Google Scholar] [CrossRef]
- Delbeke, H.; Younas, S.; Casteels, I.; Joosens, M. Current knowledge on the human eye microbiome: A systematic review of available amplicon and metagenomic sequencing data. Acta Ophthalmol. 2021, 99, 16–25. [Google Scholar] [CrossRef]





| Gene | Sequences (5′-3′) | ||
|---|---|---|---|
| Clumping factor [10] | clf | Fwd | CGA TTG GCG TGG CTT CAG |
| Rev | GCC AGT AGC CAA TGT CAC | ||
| Fibronectin-binding protein B [10] | fnbA | Fwd | GCG GAG ATC AAA GAC AA |
| Rev | CCA TCT ATA GCT GTG TGG | ||
| Fibronectin-binding protein A [10] | fnbB | Fwd | GGA GAA GGA ATT AAG GCG |
| Rev | GCC GTC GCC TTG AGC GT | ||
| Gamma-hemolysin A, B. and C [107] | hlg | Fwd | GTC AYA GAG TCC ATA ATG CA TTT AA |
| Rev | CAC CAA ATG TAT AGC CTA AAG TG | ||
| Alpha-hemolysin [10] | hla | Fwd | GGT TTA GCC TGG CCT TC |
| Rev | CAT CAC GAA CTC GTT CG | ||
| Beta-hemolysin [10] | hlb | Fwd | GCC AAA GCC GAA TCT AAG |
| Rev | CGC ATA TAC ATC CCA TGG C | ||
| LukE-LukD [107] | lukE-lukD | Fwd | TGAAAAAGGTTCAAAGTTGATACGAG |
| Rev | TGTATTCGATAGCAAAAGCAGTGCA | ||
| Mec A [10] | mecA | Fwd | GTA GAA ATG ACT GAA CGT CCG ATA A |
| Rev | CCA ATT CCA CAT TGT TTC GGT CTA A | ||
| PVL [107] | lukS-PV-lukF-PV | Fwd | ATC ATT AGG TAA AAA TGT CTG GAC ATGATC CA |
| Rev | GCATCAASTGTATTGGATAGCAAAAGC | ||
| TSST-1 [10] | tst | Fwd | AAG CCC TTT GTT GCT TGC G |
| Rev | ATC GAA CTT TGG CCC ATA CTT T |
| Gene | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| clf | fnbA | fnbB | hla | hlb | mecA | tst | hlg | luk | PVL | |
| MRSA (48) | 100.0% | 95.8% | 77.1% | 85.4% | 75.0% | 100.0% | 0.0% | 10.4% | 85.4% | 37.5% |
| MSSA (81) | 97.5% | 97.5% | 58.0% | 80.2% | 79.0% | 0.0% | 6.2% | 24.7% | 71.6% | 14.8% |
| All (129) | 98.4% | 96.9% | 65.1% | 82.2% | 77.5% | 37.2% | 3.9% | 19.4% | 76.7% | 23.3% |
| Period of Study | Location | Gram-Positive | Bacillus | CoNS | S. aureus | S. epidermidis | Streptoccoci | S. pneumoniae | Gram-negative | Moraxella | Pseudomonas sp. | P. aeruginosa | Serratia | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Alexandrakis [126] | 1990–1998 | USA, Florida | 48.0% | 19.4% | 1.3% | 6.7% | 49.6% | 25.7% | 7.6% | |||||
| Schaefer [127] | 1997–1998 | Switzerland | 1% | 22% | 40% | 5% | 8% | 5% | 9% | 5% | ||||
| Leck [128] | 1999–2001 | India, Tamil Nadu | 0.9% | 24.7% | 2.1% | 46.80% | 26.4% | 14.9% | 14% | |||||
| Leck [128] | 1999–2001 | Ghana | 0.0% | 5.0% | 5.0% | 20.0% | 15.0% | 52.5% | ||||||
| Lam [129] | 1997–1998 | Hong Kong | 46.8% | 11.4% | 53.2% | 6.3% | ||||||||
| Bourcier [82] | 1998–1999 | France, Paris | 83.1% | 48.3% | 7.7% | 9.2% | 3.4% | 16.9% | 0.5% | 10.1% | 5.3% | |||
| Zhang [130] | 2001–2002 | China, Beijing | 67.62% | 2.16% | 6% | 15.83% | 7.91% | 32.38% | 17.99% | 0.71% | ||||
| 2003–2004 | China, Beijing | 59.28% | 0.71% | 8% | 12.14% | 7.14% | 40.72% | 22.15% | 2.14% | |||||
| Geethakumari [131] | 2007–2009 | India, Kerala | 9.09% | 15.9% | 26.14% | 26.14% | ||||||||
| Orlans [132] | 1999–2004 | UK, Oxford | 56.1% | 1% | 20.1% | 18.7% | 3.6% | 43.9% | 3.6% | 25.9% | 20.9% | 3.5% | ||
| 2004–2009 | UK, Oxford | 52.4% | 0% | 32.0% | 9.4% | 2.3% | 47.7% | 6.2% | 31.20% | 28.1% | 0% | |||
| Lichtinger [6] | 2000–2003 | Canada, Toronto | 81.4% | 40.3% | 20.0% | 16.3% | 19% | 3% | 7.2% | 3.2% | ||||
| 2004–2007 | Canada, Toronto | 74% | 33.7% | 5.7% | 20.0% | 26% | 5.1% | 15.6% | 3.8% | |||||
| 2008–2010 | Canada Toronto | 69% | 33.1% | 16.9% | 15.2% | 31% | 8.4% | 21.3% | 2.8% | |||||
| Tan [133] | 2004–2006 | UK, Manchester | 71.9% | 4.4% | 35.0% | 14.5% | 12.6% | 28.1% | 4.4% | 12.6% | 3.6% | |||
| 2007–2009 | UK, Manchester | 72.7% | 5.6% | 30.6% | 15.9% | 12.9% | 27.3% | 5.6% | 9.4% | 2.3% | ||||
| 2010–2012 | UK, Manchester | 68.0% | 1.9% | 23.9% | 15.6% | 17.9% | 32.3% | 4.8% | 12.9% | 3.5% | ||||
| 2013–2015 | UK, Manchester | 66.2% | 1.7% | 15.4% | 21.7% | 16.1% | 36.1% | 13.0% | 10.0% | 3.3% | ||||
| Lin [134] | 2006–2007 | India, Southeast | 37.0% | 21.6% | ||||||||||
| 2007–2008 | India, Southeast | 32.5% | 27.0% | |||||||||||
| 2008–2009 | India, Southeast | 35.5% | 24.5% | |||||||||||
| Ting [135] | 2008–2012 | UK, England | 65.8% | 5.8% | 21.1% | 17.4% | 16.8% | 34.2% | 10.0% | 16.3% | ||||
| 2013–2017 | UK, England | 74.7% | 8.6% | 34.3% | 13.1% | 10.6% | 25.3% | 10.6% | 7.8% | |||||
| Al-Dhahari [9] | 2011–2014 | Saudi Arabia | 91.4% | 61.4% | 11.6% | 47.2% | 7.8% | 2.2% | 6% | |||||
| Hsiao [136] | 2003–2007 | Taiwan | 46.2% | 8.3% | 15.50% | 9.40% | 4.4% | 53.8% | 26.1% | 5.7% | ||||
| 2008–2012 | Taiwan | 54.8% | 7.4% | 16.40% | 7.60% | 1.7% | 45.2% | 22.9% | 4.8% | |||||
| Gautam [137] | 2017–2018 | Nepal | 21% | 56.00% | 2.0% | 12.0% | ||||||||
| Sagerfors [35] | 2004–2014 | Sweden | 79.1% | 38.0% | 14.5% | 3.0% | 20.9% | 7% | 6.4% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Astley, R.A.; Mursalin, M.H.; Coburn, P.S.; Livingston, E.T.; Nightengale, J.W.; Bagaruka, E.; Hunt, J.J.; Callegan, M.C. Ocular Bacterial Infections: A Ten-Year Survey and Review of Causative Organisms Based on the Oklahoma Experience. Microorganisms 2023, 11, 1802. https://doi.org/10.3390/microorganisms11071802
Astley RA, Mursalin MH, Coburn PS, Livingston ET, Nightengale JW, Bagaruka E, Hunt JJ, Callegan MC. Ocular Bacterial Infections: A Ten-Year Survey and Review of Causative Organisms Based on the Oklahoma Experience. Microorganisms. 2023; 11(7):1802. https://doi.org/10.3390/microorganisms11071802
Chicago/Turabian StyleAstley, Roger A., Md Huzzatul Mursalin, Phillip S. Coburn, Erin T. Livingston, James W. Nightengale, Eddy Bagaruka, Jonathan J. Hunt, and Michelle C. Callegan. 2023. "Ocular Bacterial Infections: A Ten-Year Survey and Review of Causative Organisms Based on the Oklahoma Experience" Microorganisms 11, no. 7: 1802. https://doi.org/10.3390/microorganisms11071802