
Contribution of Manure-Spreading Operations to Bioaerosols and Antibiotic Resistance Genes’ Emission

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Manure Spreading
2.2. Sampling of Manure and Air
2.3. Processing of Manure and Air Samples
2.4. Quantification of Total Bacteria and Fecal Indicators Using Classic CFX qPCR
2.5. Quantification of ARGs Using the TAKARA Platform
2.6. Calculation of Emission Rates for Total Bacteria, Fecal Indicators, Aerococcus Phage, and ARGs
2.7. Statistical Analyses
3. Results
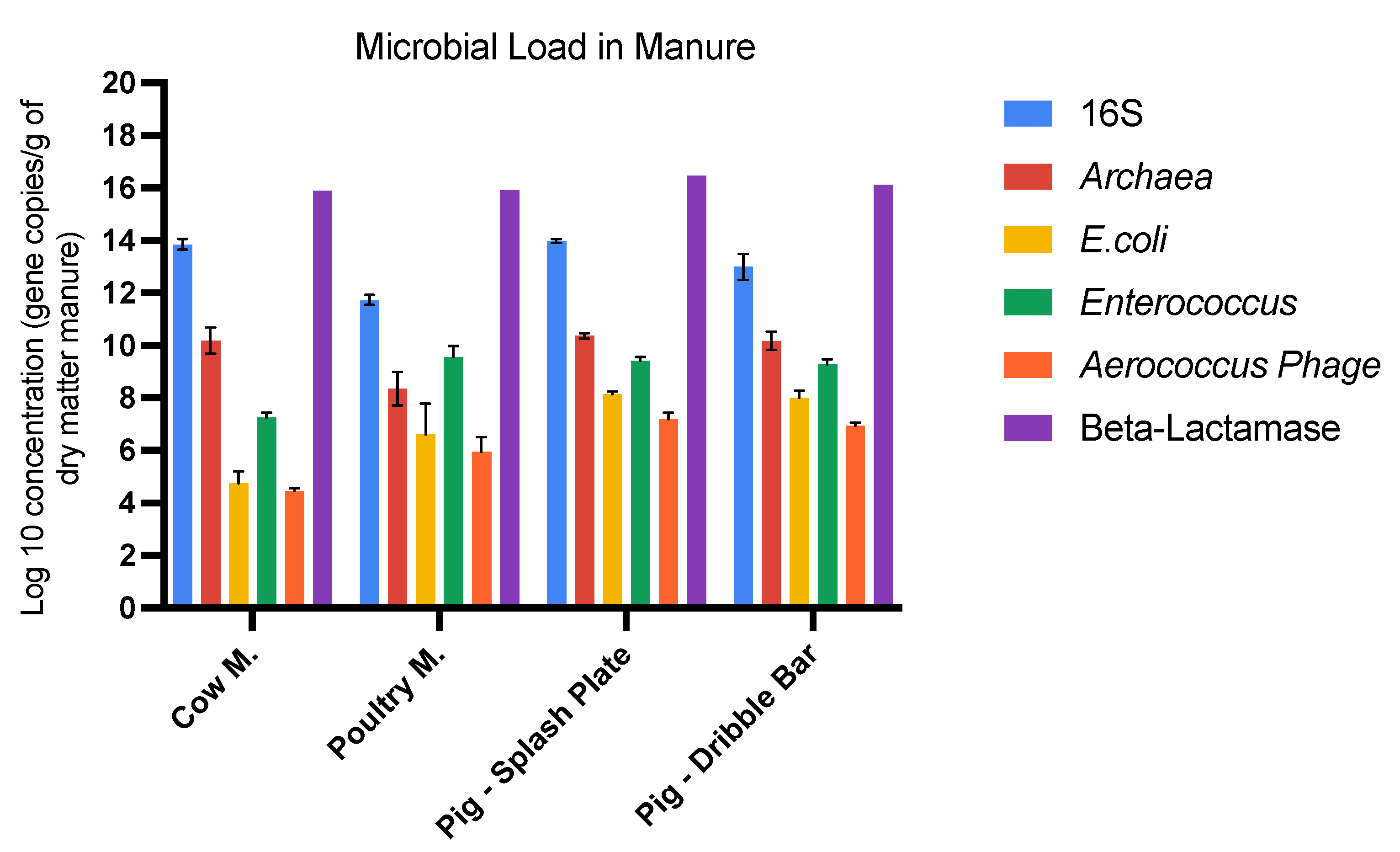
3.1. Bacteria, Archaea, Fecal Indicators, and ARGs in Manure
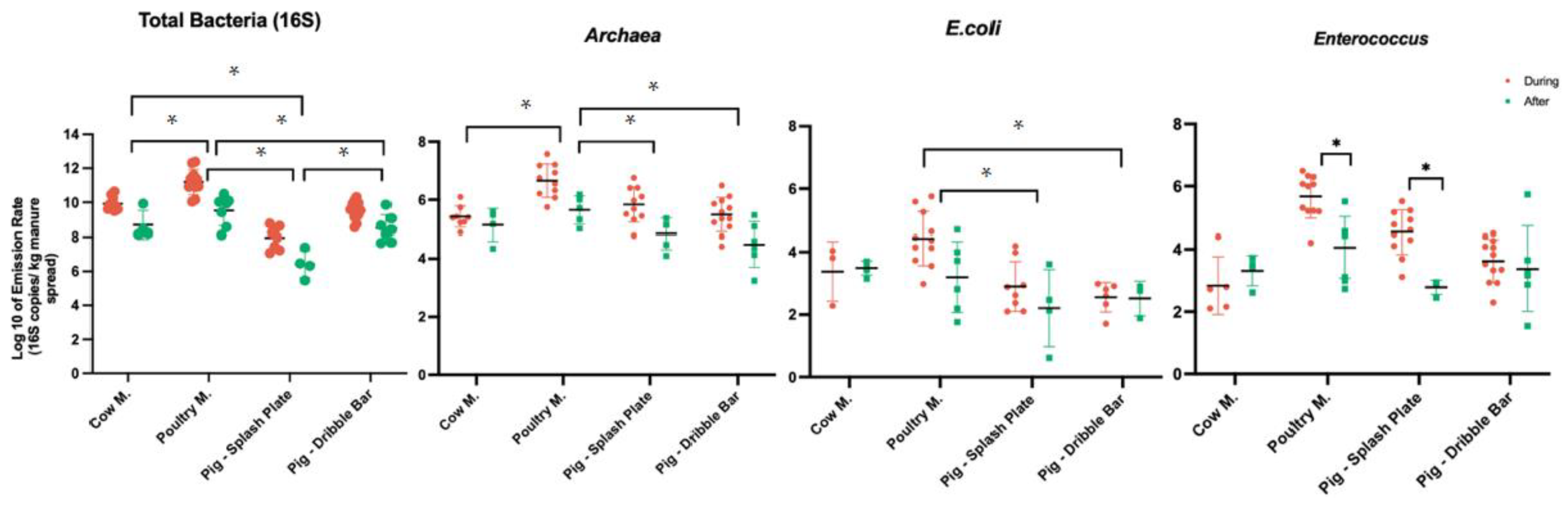
3.2. Airborne Emissions of Total Bacteria and Dust Particles
3.3. Airborne Emissions of Fecal Indicators
3.4. Antibiotic Resistance Genes’ (ARGs) Emission Rates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DeBaun, M.R.; Lai, C.; Sanchez, M.; Chen, M.J.; Goodnough, L.H.; Chang, A.; Bishop, J.A.; Gardner, M.J. Antibiotic Resistance: Still a Cause of Concern? OTA Int. 2021, 4, e104. [Google Scholar] [CrossRef]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- WHO Antimicrobial Resistance. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 6 May 2023).
- Kang, J.; Liu, Y.; Chen, X.; Xu, F.; Wang, H.; Xiong, W.; Li, X. Metagenomic Insights into the Antibiotic Resistomes of Typical Chinese Dairy Farm Environments. Front. Microbiol. 2022, 13, 3562. [Google Scholar] [CrossRef] [PubMed]
- Gerba, C.P.; Smith, J.E. Sources of Pathogenic Microorganisms and Their Fate during Land Application of Wastes. J. Environ. Qual. 2005, 34, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Han, W.; Chen, S.; Dong, W.; Qiao, M.; Hu, C.; Liu, B. Fifteen-Year Application of Manure and Chemical Fertilizers Differently Impacts Soil ARGs and Microbial Community Structure. Front. Microbiol. 2020, 11, 62. [Google Scholar] [CrossRef]
- Jahne, M.A.; Rogers, S.W.; Holsen, T.M.; Grimberg, S.J. Quantitative Microbial Risk Assessment of Bioaerosols from a Manure Application Site. Aerobiologia 2015, 31, 73–87. [Google Scholar] [CrossRef]
- Mceachran, A.D.; Blackwell, B.R.; Hanson, J.D.; Wooten, K.J.; Mayer, G.D.; Cox, S.B.; Smith, P.N. Supplemental Material: Antibiotics, Bacteria, and Antibiotic Resistance Genes: Aerial Transport from Cattle Feed Yards via Particulate Matter. Environ. Health Perspect. 2015, 123, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Jaffrezic, A.; Jardé, E.; Pourcher, A.-M.; Gourmelon, M.; Caprais, M.-P.; Heddadj, D.; Cottinet, P.; Bilal, M.; Derrien, M.; Marti, R.; et al. Microbial and Chemical Markers: Runoff Transfer in Animal Manure-Amended Soils. J. Environ. Qual. 2011, 40, 959–968. [Google Scholar] [CrossRef]
- Gibbs, S.G.; Green, C.F.; Tarwater, P.M.; Mota, L.C.; Mena, K.D.; Scarpino, P.V. Isolation of Antibiotic-Resistant Bacteria from the Air Plume Downwind of a Swine Confined or Concentrated Animal Feeding Operation. Environ. Health Perspect. 2006, 114, 1032–1037. [Google Scholar] [CrossRef] [Green Version]
- Ko, G.; Simmons, O.D.; Likirdopulos, C.A.; Worley-Davis, L.; Williams, M.; Sobsey, M.D. Investigation of Bioaerosols Released from Swine Farms Using Conventional and Alternative Waste Treatment and Management Technologies. Environ. Sci. Technol. 2008, 42, 8849–8857. [Google Scholar] [CrossRef]
- Gladding, T.L.; Rolph, C.A.; Gwyther, C.L.; Kinnersley, R.; Walsh, K.; Tyrrel, S. Concentration and Composition of Bioaerosol Emissions from Intensive Farms: Pig and Poultry Livestock. J. Environ. Manag. 2020, 272, 111052. [Google Scholar] [CrossRef]
- Brooks, J.P.; Tanner, B.D.; Gerba, C.P.; Pepper, I.L. The Measurement of Aerosolized Endotoxin from Land Application of Class B Biosolids in Southeast Arizona. Can. J. Microbiol. 2006, 52, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Manure Spreader Guide, Colerain, North Carolina. Available online: https://www.robertsonequipment.com/departments/our-service-department/kuhn-product-tips/manure-spreader-guide/ (accessed on 10 October 2022).
- Vaisala MX50G | Manure Spreaders MX50G | Manure Spreaders | Ground Drive—Towable/Self-Contained | Wallenstein Outdoor Power Equipment. Available online: https://www.wallensteinequipment.com/in/en/model/mx50g (accessed on 14 January 2023).
- Different Types Of Manure Spreaders and Calibration—FarmTech. Available online: https://www.farmtech.com.au/2022/03/18/different-types-of-manure-spreaders-and-calibration/ (accessed on 20 September 2022).
- Cheng, Y.S.; Irshad, H.; McFarland, A.R.; Su, W.C.; Zhou, Y.; Barringer, D. An Aerosol Wind Tunnel for Evaluation of Massive-Flow Air Samplers and Calibration of Snow White Sampler. Aerosol Sci. Technol. 2004, 38, 1099–1107. [Google Scholar] [CrossRef]
- Griffiths, W.D.; Stewart, I.W.; Futter, S.J.; Upton, S.L.; Mark, D. The Development of Sampling Methods for the Assessment of Indoor Bioaerosols. J. Aerosol Sci. 1997, 28, 437–457. [Google Scholar] [CrossRef]
- Bae, G.N.; Kim, J.B.; Woo, S.H.; Yook, S.J.; Park, H.K.; Yoon, H.H. Non-Powered Dust Removal Device Attached Underneath A Train Cabin In A Subway Tunnel. WIT Trans. Built Environ. 2015, 168, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Desbiens, V.; Brassard, P.; Baghdadi, M.; Létourneau, V.; Turgeon, N.; Duchaine, C.; Trivino Arevalo, A.; Godbout, S. Comparison of Air Contaminants Emissions from Two Pig Slurry Spreading Methods in a Controlled Environment—Technical Library of the CSBE-SCGAB. Available online: https://library.csbe-scgab.ca/all-publications/5968:comparison-of-air-contaminants-emissions-from-two-pig-slurry-spreading-methods-in-a-controlled-environment (accessed on 23 January 2023).
- Nijhuis, R.H.T.; Veldman, K.T.; Schelfaut, J.; Van Essen-Zandbergen, A.; Wessels, E.; Claas, E.C.J.; Gooskens, J. Detection of the Plasmid-Mediated Colistin-Resistance Gene Mcr-1 in Clinical Isolates and Stool Specimens Obtained from Hospitalized Patients Using a Newly Developed Real-Time PCR Assay. J. Antimicrob. Chemother. 2016, 71, 2344–2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stedtfeld, R.D.; Guo, X.; Stedtfeld, T.M.; Sheng, H.; Williams, M.R.; Hauschild, K.; Gunturu, S.; Tift, L.; Wang, F.; Howe, A.; et al. Primer Set 2.0 for Highly Parallel QPCR Array Targeting Antibiotic Resistance Genes and Mobile Genetic Elements. FEMS Microbiol. Ecol. 2018, 94, fiy130. [Google Scholar] [CrossRef]
- Bélanger Cayouette, A.; Ouradou, A.; Veillette, M.; Turgeon, N.; George, P.B.L.; Corbin, S.; Boulanger, C.; Duchaine, C.; Bédard, E. Effect of Odor-Treatment Biofilter on Bioaerosols from an Indoor Wastewater Treatment Plant. Atmosphere 2022, 13, 1723. [Google Scholar] [CrossRef]
- Brunner, E.; Domhof, S.; Langer, F. Nonparametric Analysis of Longitudinal Data in Factorial Experiments; Wiley Series in Probability and Statistics; J. Wiley: New York, NY, USA, 2001. [Google Scholar]
- Nikaeen, M.; Hatamzadeh, M.; Hasanzadeh, A.; Sahami, E.; Joodan, I. Bioaerosol Emissions Arising during Application of Municipal Solid-Waste Compost. Aerobiologia 2009, 25, 1–6. [Google Scholar] [CrossRef]
- Chien, Y.C.; Chen, C.J.; Lin, T.H.; Chen, S.H.; Chien, Y.C. Characteristics of Microbial Aerosols Released from Chicken and Swine Feces. J. Air Waste Manag. Assoc. 2011, 61, 882–889. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Duan, Y.; Awasthi, S.K.; Liu, T.; Zhang, Z.; Kim, S.H.; Pandey, A. Effect of Biochar on Emission, Maturity and Bacterial Dynamics during Sheep Manure Compositing. Renew. Energy 2020, 152, 421–429. [Google Scholar] [CrossRef]
- Dungan, R.S. BOARD-INVITED REVIEW: Fate and Transport of Bioaerosols Associated with Livestock Operations and Manures. J. Anim. Sci. 2010, 88, 3693–3706. [Google Scholar] [CrossRef] [PubMed]
- Cambra-López, M.; Aarnink, A.J.A.; Zhao, Y.; Calvet, S.; Torres, A.G. Airborne Particulate Matter from Livestock Production Systems: A Review of an Air Pollution Problem. Environ. Pollut. 2010, 158, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Liu, L.; Meng, Y.; Han, H.; Xiong, R.; Li, Y.; Chen, L. Archaeal and Bacterial Community Structures of Rural Household Biogas Digesters with Different Raw Materials in Qinghai Plateau. Biotechnol. Lett. 2021, 43, 1337–1348. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zamudio Cañas, E.M.; Zhu, Z.; Linville, J.L.; Chen, S.; He, Q. Robustness of Archaeal Populations in Anaerobic Co-Digestion of Dairy and Poultry Wastes. Bioresour. Technol. 2011, 102, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Lecours, P.B.; Veillette, M.; Marsolais, D.; Cormier, Y.; Kirychuk, S.; Duchaine, C. Archaea in Bioaerosols in Dairy Farms, Poultry Houses and Wastewater Treatment Plants and Their Role in Lung Inflammation. Chemical and Biological Hazards Prevention, Studies and Research Projects, Report R-845, IRSST, Montréal. Available online: https://numerique.banq.qc.ca/patrimoine/details/52327/2419597 (accessed on 10 March 2023).
- Aust, M.O.; Godlinski, F.; Travis, G.R.; Hao, X.; McAllister, T.A.; Leinweber, P.; Thiele-Bruhn, S. Distribution of Sulfamethazine, Chlortetracycline and Tylosin in Manure and Soil of Canadian Feedlots after Subtherapeutic Use in Cattle. Environ. Pollut. 2008, 156, 1243–1251. [Google Scholar] [CrossRef]
- Diarra, M.S.; Rempel, H.; Champagne, J.; Masson, L.; Pritchard, J.; Topp, E. Distribution of Antimicrobial Resistance and Virulence Genes in Enterococcus Spp. and Characterization of Isolates from Broiler Chickens. Appl. Environ. Microbiol. 2010, 76, 8033–8043. [Google Scholar] [CrossRef] [Green Version]
- Holman, D.B.; Chénier, M.R. Antimicrobial Use in Swine Production and Its Effect on the Swine Gut Microbiota and Antimicrobial Resistance. Can. J. Microbiol. 2015, 61, 785–798. [Google Scholar] [CrossRef]
- Xu, Y.; Li, H.; Shi, R.; Lv, J.; Li, B.; Yang, F.; Zheng, X.; Xu, J. Antibiotic Resistance Genes in Different Animal Manures and Their Derived Organic Fertilizer. Environ. Sci. Eur. 2020, 32, 102. [Google Scholar] [CrossRef]
- Wolters, B.; Ding, G.C.; Kreuzig, R.; Smalla, K. Full-Scale Mesophilic Biogas Plants Using Manure as C-Source: Bacterial Community Shifts along the Process Cause Changes in the Abundance of Resistance Genes and Mobile Genetic Elements. FEMS Microbiol. Ecol. 2016, 92, fiv163. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Li, Y.; Zhang, Y.; Zhang, H.; Guo, Z.; Liu, J. Deciphering of Microbial Diversity and Antibiotic Resistome of Bioaerosols in Swine Confinement Buildings. Sci. Total Environ. 2021, 781, 147056. [Google Scholar] [CrossRef]
- Jones, A.M.; Harrison, R.M. The Effects of Meteorological Factors on Atmospheric Bioaerosol Concentrations—A Review. Sci. Total Environ. 2004, 326, 151–180. [Google Scholar] [CrossRef]
- Girard, M.; Gasser, M.-O.; Lévesque, A.; Tremblay, M.-É. Impact de la Date d’épandage du Lisier de Porc sur le Rendement des Cultures Ainsi Que la Qualité du sol, de l’eau et de l’air. Available online: https://irda.blob.core.windows.net/media/5347/girard-et-al-2017-impact_de_la_date_depandage_du_lisier_de_porc_sur_le_rendement_des_cultures_ainsi_que_la_qualite_du_sol_de_leau_et_de_lair.pdf (accessed on 30 June 2023).
- Johnson, J.Y.; Thomas, J.E.; Graham, T.A.; Townshend, I.; Byrne, J.; Selinger, L.B.; Gannon, V.P. Prevalence of Escherichia Coli O157:H7 and Salmonella Spp. in Surface Waters of Southern Alberta and Its Relation to Manure Sources. Can. J. Microbiol. 2011, 49, 326–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shridhar, P.B.; Patel, I.R.; Gangiredla, J.; Noll, L.W.; Shi, X.; Bai, J.; Elkins, C.A.; Strockbine, N.A.; Nagaraja, T.G. Genetic Analysis of Virulence Potential of Escherichia Coli O104 Serotypes Isolated from Cattle Feces Using Whole Genome Sequencing. Front. Microbiol. 2018, 9, 341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manning, S.D. Deadly Diseases and Epidemics: Escherichia Coli Infections; Nuevos sistemas de comunicación e información; Infobase Publishing: New York, NY, USA, 2010; pp. 1–134. [Google Scholar]
- Oliver, J.P.; Gooch, C.A.; Lansing, S.; Schueler, J.; Hurst, J.J.; Sassoubre, L.; Crossette, E.M.; Aga, D.S. Invited Review: Fate of Antibiotic Residues, Antibiotic-Resistant Bacteria, and Antibiotic Resistance Genes in US Dairy Manure Management Systems. J. Dairy Sci. 2020, 103, 1051–1071. [Google Scholar] [CrossRef] [PubMed]
- Bach, H.J.; Tomanova, J.; Schloter, M.; Munch, J.C. Enumeration of Total Bacteria and Bacteria with Genes for Proteolytic Activity in Pure Cultures and in Environmental Samples by Quantitative PCR Mediated Amplification. J. Microbiol. Methods 2002, 49, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Malinen, E.; Kassinen, A.; Rinttilä, T.; Palva, A. Comparison of Real-Time PCR with SYBR Green I or 5′-Nuclease Assays and Dot-Blot Hybridization with RDNA-Targeted Oligonucleotide Probes in Quantification of Selected Faecal Bacteria. Microbiology 2003, 149, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Baker, G.C.; Smith, J.J.; Cowan, D.A. Review and Re-Analysis of Domain-Specific 16S Primers. J. Microbiol. Methods 2003, 55, 541–555. [Google Scholar] [CrossRef] [Green Version]
- Robb, F.T.; DasSarma, S.; Fleischmann, E.M. Archaea: A Laboratory Manual; Cold Spring Harbor Laboratory: New York, NY, USA, 1995; p. 217. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| (A) | ||||
|---|---|---|---|---|
| Gene Copies/g Dry Matter of Manure | ||||
| Cow Manure | Poultry Manure | Pig Slurry with Splash Plate | Pig Slurry with Dribble Bar | |
| Total bacteria | 7.61 × 1013 | 5.70 × 1011 | 9.29 × 1013 | 1.91 × 1013 |
| Enterococcus | 1.91 × 107 | 4.86 × 109 | 2.75 × 109 | 2.09 × 109 |
| E. coli | 7.65 × 104 | 1.19 × 108 | 1.43 × 108 | 1.19 × 108 |
| Archaea | 2.54 × 1010 | 4.49 × 108 | 2.39 × 1010 | 1.83 × 1010 |
| Aerococcus Phage | 1.45 × 104 | 1.63 × 106 | 1.74 × 107 | 9.06 × 106 |
| (B) | ||||
| Gene Copies/g Dry Matter of Manure | ||||
| Cow Manure | Poultry Manure | Pig Slurry with Splash Plate | Pig Slurry with Dribble Bar | |
| Aminoglycosides | 7.15 × 107 | 2.77 × 1015 | 4.84 × 109 | 6.63 × 1015 |
| Beta-Lactamase | 7.75 × 1015 | 8.24 × 1015 | 2.96 × 1016 | 1.29 × 1016 |
| Erythromycin | 2.77 × 1010 | 7.72 × 1015 | 6.72 × 1015 | 1.01 × 109 |
| MGE | 5.61 × 109 | 1.16 × 1016 | 4.34 × 1011 | 4.12 × 1016 |
| Tetracycline | 2.20 × 109 | 2.24 × 1015 | 2.31 × 1012 | 4.33 × 1015 |
| Sulfonamide | 1.52 × 1010 | 7.04 × 109 | 5.77 × 1011 | 2.75 × 1011 |
| Quinolones | 3.27 × 1010 | 1.61 × 108 | 1.66 × 1016 | 6.50 × 109 |
| Vancomycin | 1.40 × 1015 | 6.41 × 1015 | 8.13 × 1016 | 7.22 × 1016 |
| Absolute Number of Total ARGs in the Air during Spreading (Gene Copies/m3) | Absolute Number of Total ARGs in the Air after Spreading (Gene Copies/m3) | |
|---|---|---|
| Cow manure | 1.23 × 106 | 7.98 × 105 |
| Poultry manure | 4.91 × 106 | 1.31 × 106 |
| Pig slurry with splash plate | 1.23 × 106 | 1.25 × 105 |
| Pig slurry with dribble bar | 3.33 × 107 | 7.18 × 106 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baghdadi, M.; Brassard, P.; Godbout, S.; Létourneau, V.; Turgeon, N.; Rossi, F.; Lachance, É.; Veillette, M.; Gaucher, M.-L.; Duchaine, C. Contribution of Manure-Spreading Operations to Bioaerosols and Antibiotic Resistance Genes’ Emission. Microorganisms 2023, 11, 1797. https://doi.org/10.3390/microorganisms11071797
Baghdadi M, Brassard P, Godbout S, Létourneau V, Turgeon N, Rossi F, Lachance É, Veillette M, Gaucher M-L, Duchaine C. Contribution of Manure-Spreading Operations to Bioaerosols and Antibiotic Resistance Genes’ Emission. Microorganisms. 2023; 11(7):1797. https://doi.org/10.3390/microorganisms11071797
Chicago/Turabian StyleBaghdadi, Mahsa, Patrick Brassard, Stéphane Godbout, Valérie Létourneau, Nathalie Turgeon, Florent Rossi, Émie Lachance, Marc Veillette, Marie-Lou Gaucher, and Caroline Duchaine. 2023. "Contribution of Manure-Spreading Operations to Bioaerosols and Antibiotic Resistance Genes’ Emission" Microorganisms 11, no. 7: 1797. https://doi.org/10.3390/microorganisms11071797