
Deciphering the Association between Campylobacter Colonization and Microbiota Composition in the Intestine of Commercial Broilers

Abstract
:1. Introduction
2. Materials and Methods
2.1. Farm Selection
2.2. Sample Collection
2.3. Campylobacter Status Determination of Farms
2.4. DNA Extraction, Amplification, and Sequencing
2.5. Data Analysis
3. Results
3.1. Campylobacter Status of the Selected Farms
3.2. Microbial Diversity
3.2.1. 16S rRNA Gene Sequencing Outputs Overview
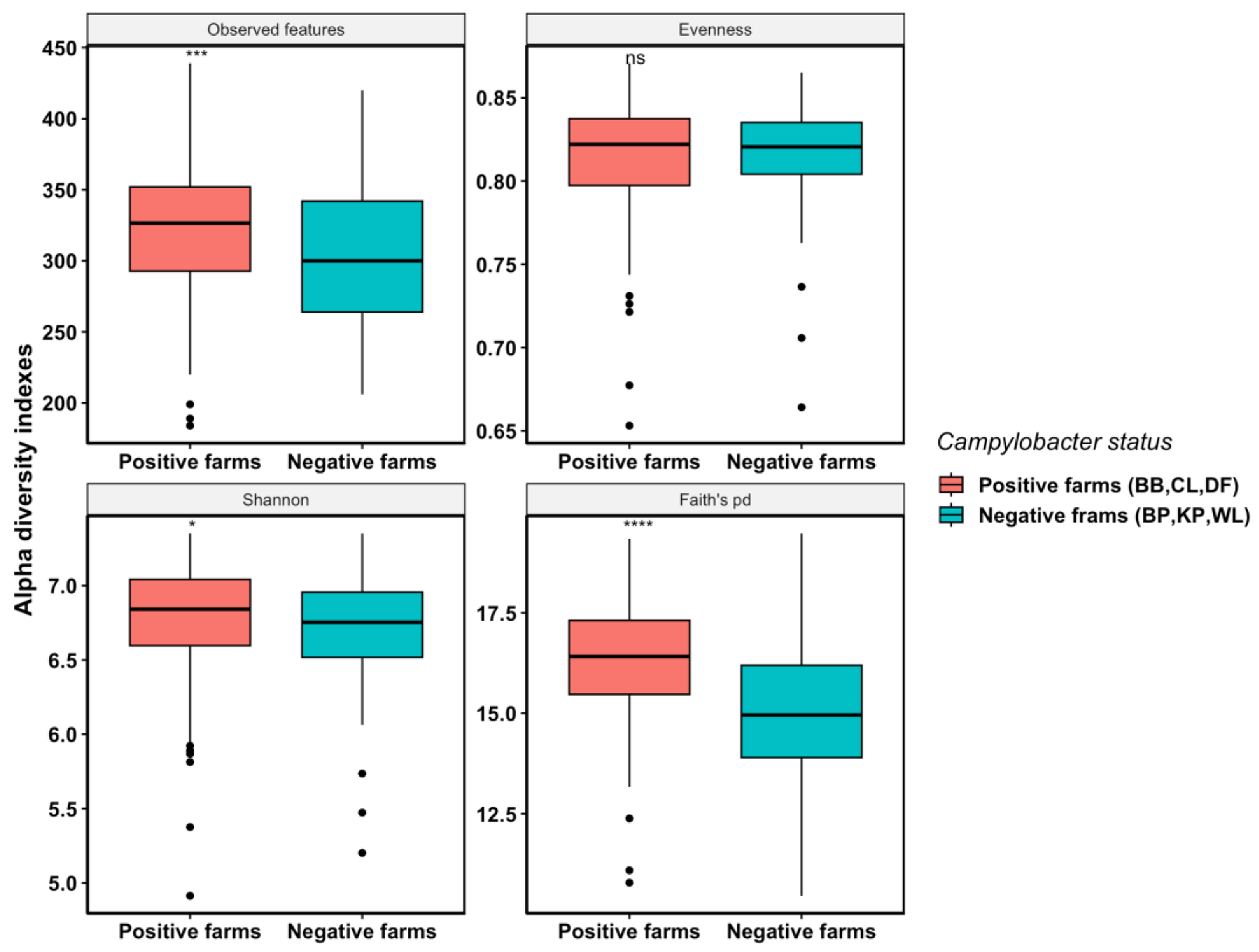
3.2.2. Campylobacter-Positive Farms Had Higher Cecal Microbiota Species Richness and Phylogenetic Diversity Compared with Campylobacter-Negative Farms
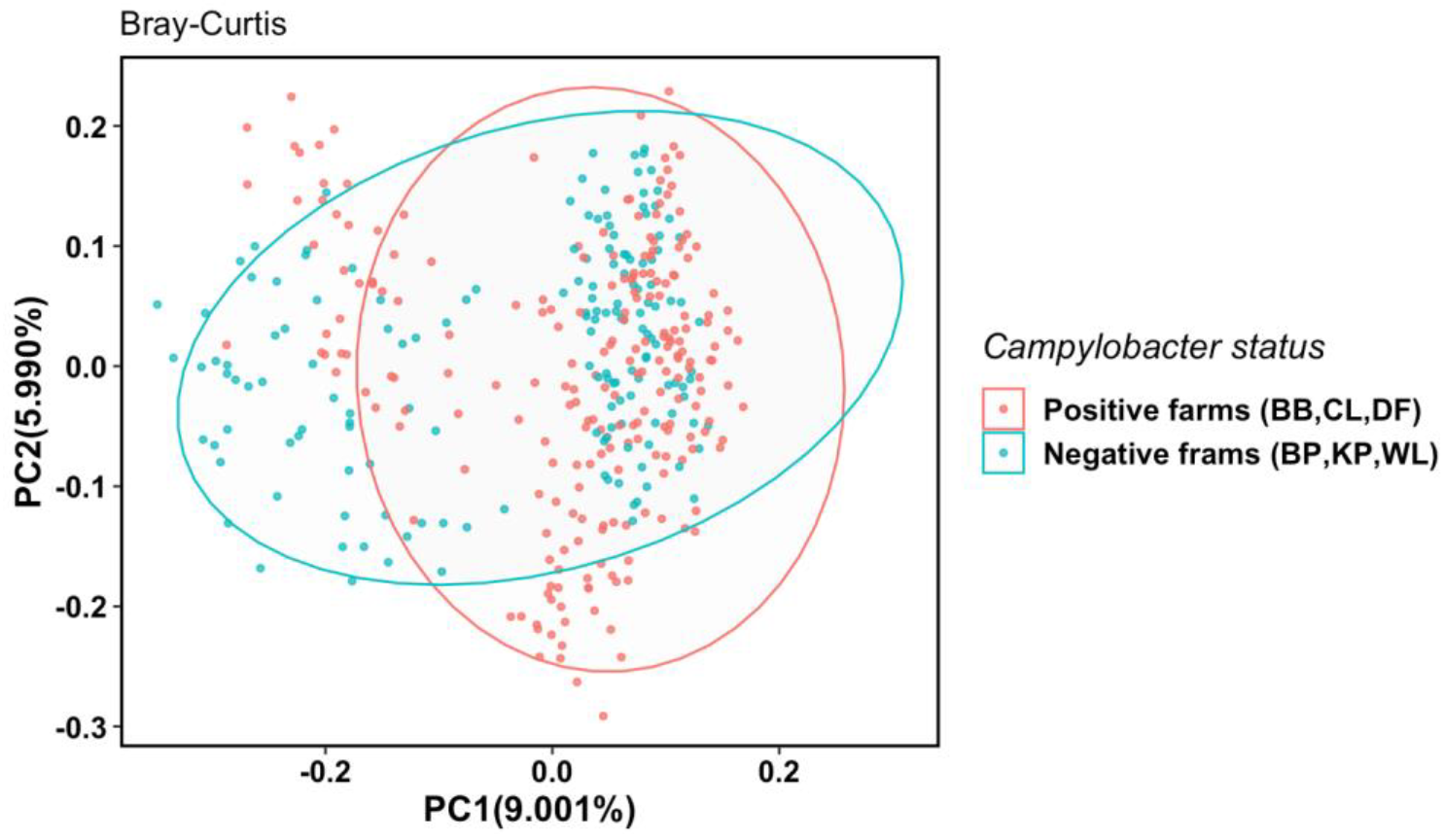
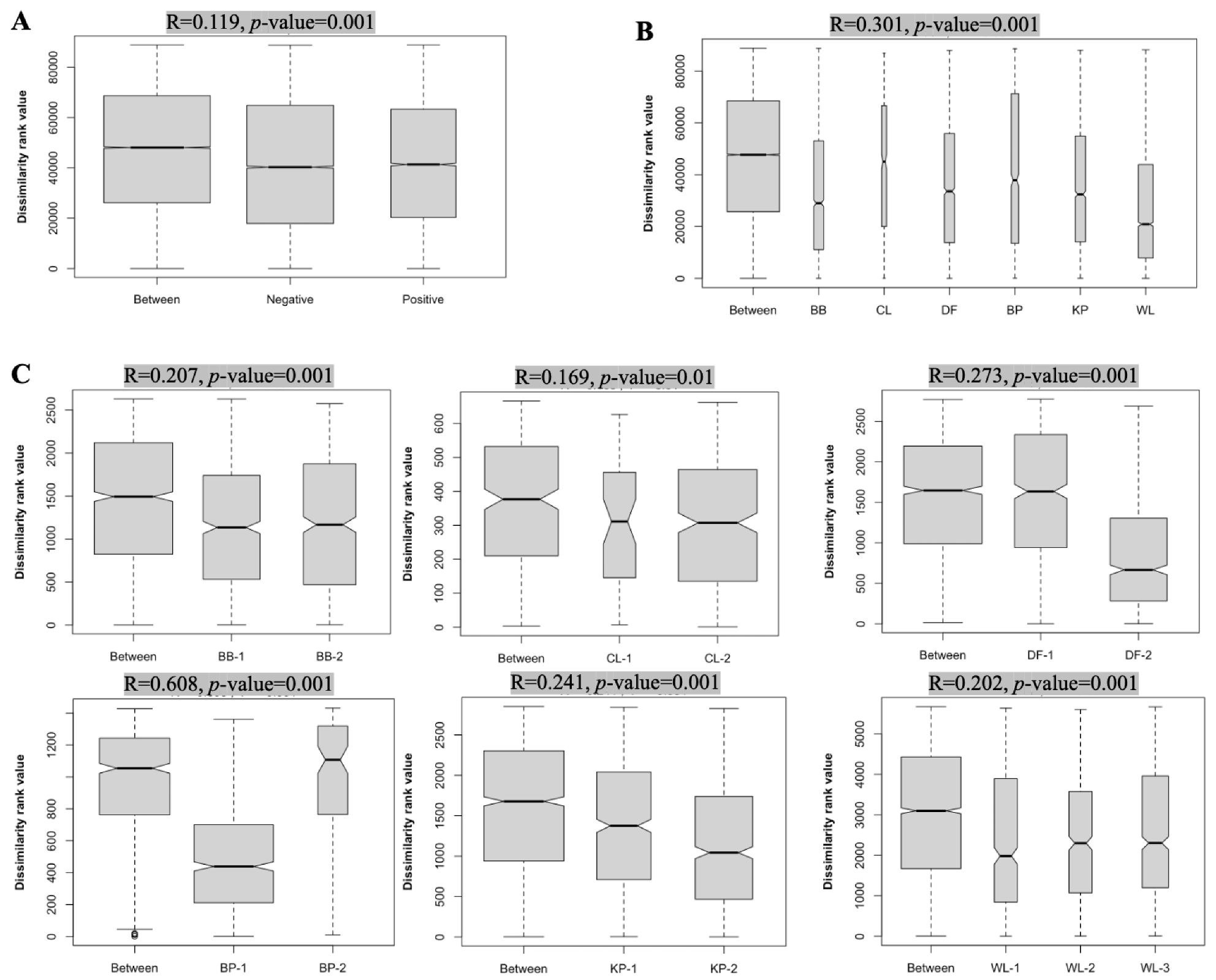
3.2.3. Multiple Factors Contribute to Cecal Microbiota Composition Differences among Farms
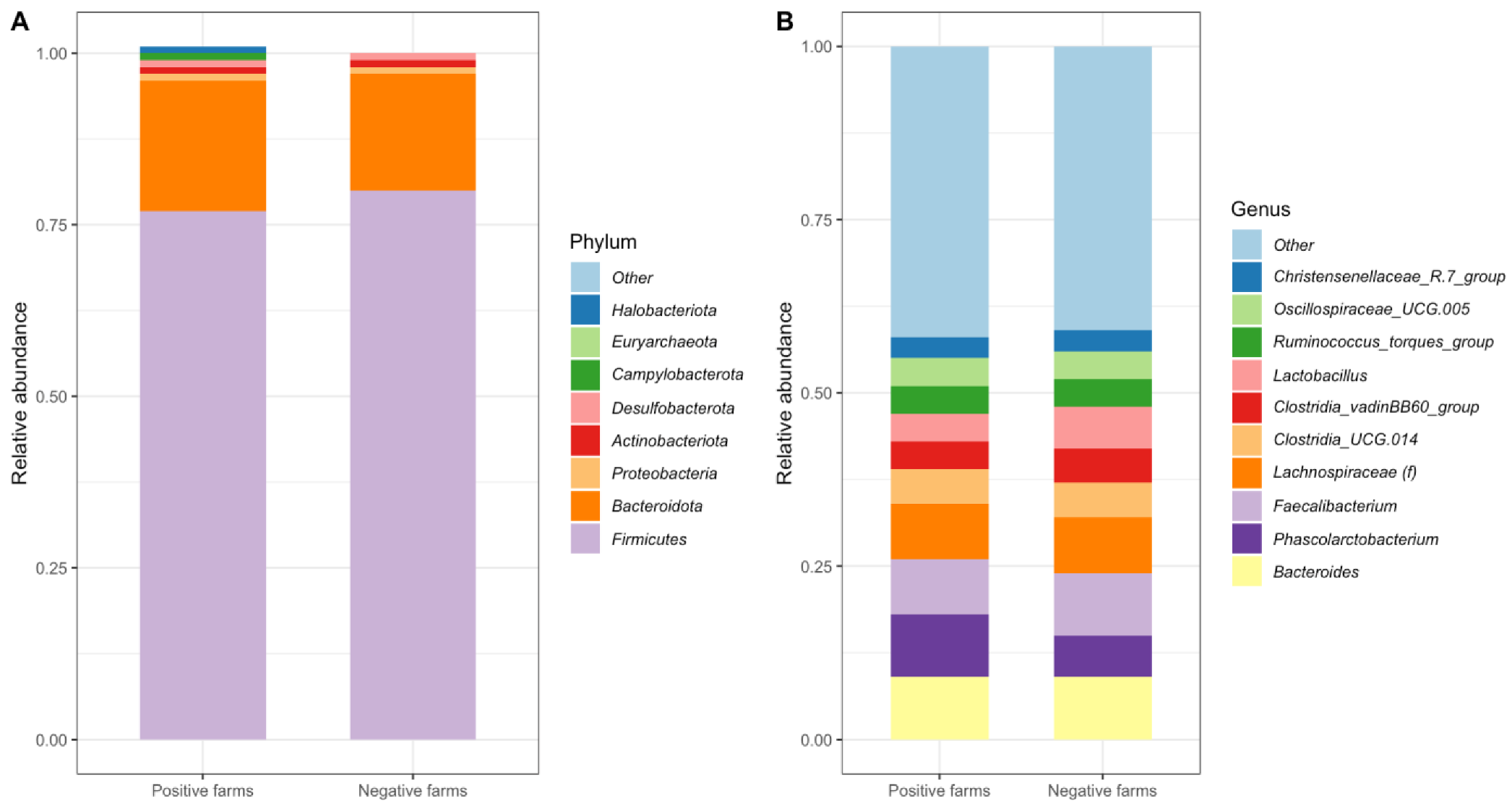
3.3. Cecal Microbiota Overall Was Dominated by Similar Taxa between Campylobacter-Positive and -Negative Farms
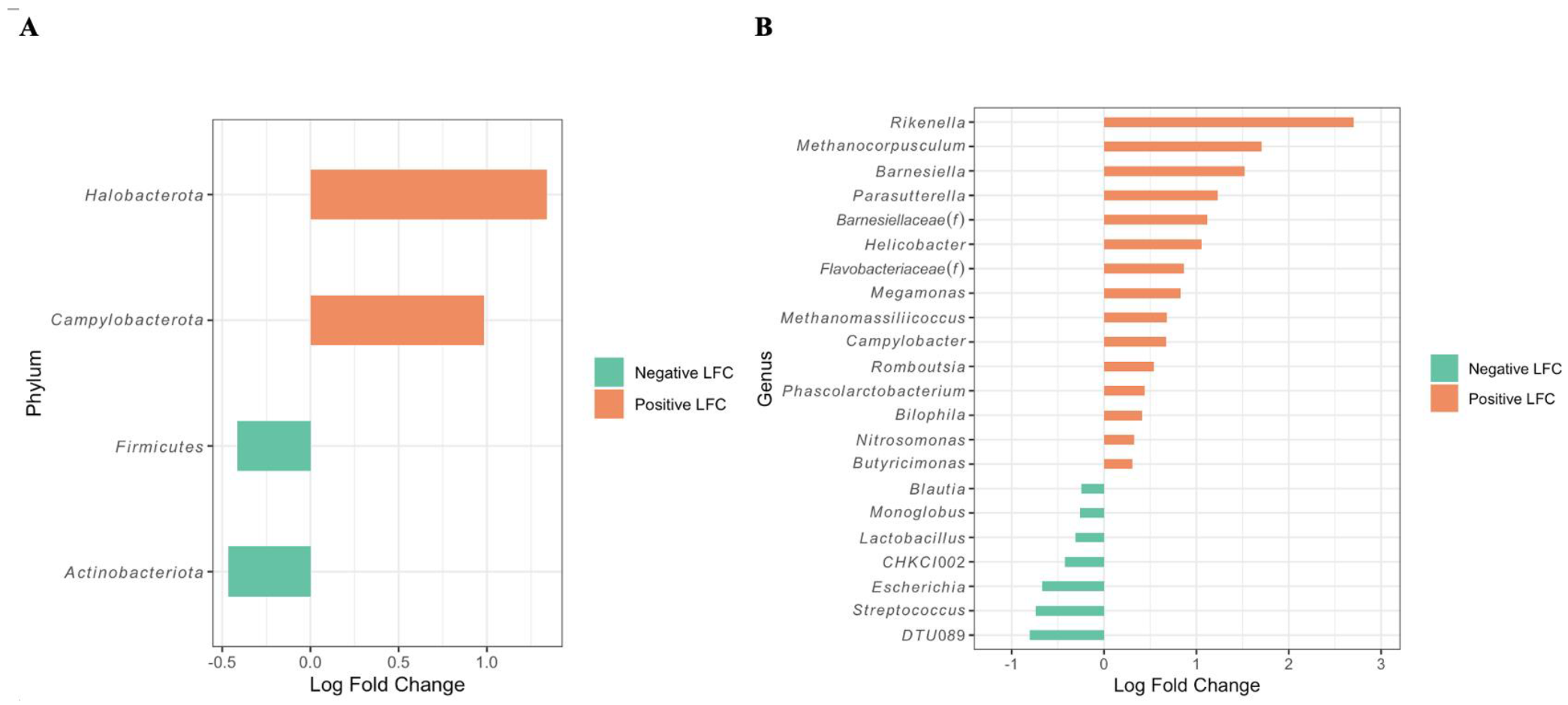
3.4. Differentially Abundant Taxa between Campylobacter-Positive and -Negative Farms as Revealed by ANCOM and Correlation Tests
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Havelaar, A.H.; Kirk, M.D.; Torgerson, P.R.; Gibb, H.J.; Hald, T.; Lake, R.J.; Praet, N.; Bellinger, D.C.; de Silva, N.R.; Gargouri, N.; et al. World Health Organization Global Estimates and Regional Comparisons of the Burden of Foodborne Disease in 2010. PLoS Med. 2015, 12, e1001923. [Google Scholar] [CrossRef] [Green Version]
- Questions and Answers|Campylobacter|CDC. Available online: https://www.cdc.gov/campylobacter/faq.html (accessed on 19 February 2023).
- Collins, J.P. Preliminary Incidence and Trends of Infections Caused by Pathogens Transmitted Commonly through Food—Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2016–2021. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 1260–1264. [Google Scholar] [CrossRef] [PubMed]
- Peterson, M.C. Clinical Aspects of Campylobacter jejuni Infections in Adults. West. J. Med. 1994, 161, 148–152. [Google Scholar]
- White, A.E.; Ciampa, N.; Chen, Y.; Kirk, M.; Nesbitt, A.; Bruce, B.B.; Walter, E.S. Characteristics of Campylobacter, Salmonella Infections and Acute Gastroenteritis in Older Adults in Australia, Canada, and the United States. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2019, 69, 1545–1552. [Google Scholar] [CrossRef] [PubMed]
- Facciolà, A.; Riso, R.; Avventuroso, E.; Visalli, G.; Delia, S.A.; Laganà, P. Campylobacter: From Microbiology to Prevention. J. Prev. Med. Hyg. 2017, 58, E79–E92. [Google Scholar] [PubMed]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [Green Version]
- Thépault, A.; Rose, V.; Queguiner, M.; Chemaly, M.; Rivoal, K. Dogs and Cats: Reservoirs for Highly Diverse Campylobacter jejuni and a Potential Source of Human Exposure. Anim. Open Access J. 2020, 10, 838. [Google Scholar] [CrossRef]
- Mughini Gras, L.; Smid, J.H.; Wagenaar, J.A.; de Boer, A.G.; Havelaar, A.H.; Friesema, I.H.M.; French, N.P.; Busani, L.; van Pelt, W. Risk Factors for Campylobacteriosis of Chicken, Ruminant, and Environmental Origin: A Combined Case-Control and Source Attribution Analysis. PLoS ONE 2012, 7, e42599. [Google Scholar] [CrossRef] [Green Version]
- Mughini-Gras, L.; Pijnacker, R.; Coipan, C.; Mulder, A.C.; Fernandes Veludo, A.; de Rijk, S.; van Hoek, A.H.A.M.; Buij, R.; Muskens, G.; Koene, M.; et al. Sources and Transmission Routes of Campylobacteriosis: A Combined Analysis of Genome and Exposure Data. J. Infect. 2021, 82, 216–226. [Google Scholar] [CrossRef]
- Li, M.; Havelaar, A.H.; Hoffmann, S.; Hald, T.; Kirk, M.D.; Torgerson, P.R.; Devleesschauwer, B. Global Disease Burden of Pathogens in Animal Source Foods, 2010. PLoS ONE 2019, 14, e0216545. [Google Scholar] [CrossRef]
- Sahin, O.; Kassem, I.I.; Shen, Z.; Lin, J.; Rajashekara, G.; Zhang, Q. Campylobacter in Poultry: Ecology and Potential Interventions. Avian Dis. 2015, 59, 185–200. [Google Scholar] [CrossRef]
- Zhang, Q.; Sahin, O. Campylobacteriosis. In Diseases of Poultry; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 754–769. ISBN 978-1-119-37119-9. [Google Scholar]
- Luangtongkum, T.; Morishita, T.Y.; Ison, A.J.; Huang, S.; McDermott, P.F.; Zhang, Q. Effect of Conventional and Organic Production Practices on the Prevalence and Antimicrobial Resistance of Campylobacter Spp. in Poultry. Appl. Environ. Microbiol. 2006, 72, 3600–3607. [Google Scholar] [CrossRef] [Green Version]
- Powell, L.F.; Lawes, J.R.; Clifton-Hadley, F.A.; Rodgers, J.; Harris, K.; Evans, S.J.; Vidal, A. The Prevalence of Campylobacter Spp. in Broiler Flocks and on Broiler Carcases, and the Risks Associated with Highly Contaminated Carcases. Epidemiol. Infect. 2012, 140, 2233–2246. [Google Scholar] [CrossRef] [Green Version]
- Schets, F.M.; Jacobs-Reitsma, W.F.; van der Plaats, R.Q.J.; Heer, L.K.-D.; van Hoek, A.H.A.M.; Hamidjaja, R.A.; de Roda Husman, A.M.; Blaak, H. Prevalence and Types of Campylobacter on Poultry Farms and in Their Direct Environment. J. Water Health 2017, 15, 849–862. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Jiang, Q.; Tang, H.; Wang, Z.; Yin, Y.; Ren, F.; Kong, L.; Jiao, X.; Huang, J. Characterization and Prevalence of Campylobacter Spp. from Broiler Chicken Rearing Period to the Slaughtering Process in Eastern China. Front. Vet. Sci. 2020, 7, 227. [Google Scholar] [CrossRef]
- Poudel, S.; Li, T.; Chen, S.; Zhang, X.; Cheng, W.-H.; Sukumaran, A.T.; Kiess, A.S.; Zhang, L. Prevalence, Antimicrobial Resistance, and Molecular Characterization of Campylobacter Isolated from Broilers and Broiler Meat Raised without Antibiotics. Microbiol. Spectr. 2022, 10, e00251-22. [Google Scholar] [CrossRef]
- Golden, C.E.; Mishra, A. Prevalence of Salmonella and Campylobacter Spp. in Alternative and Conventionally Produced Chicken in the United States: A Systematic Review and Meta-Analysis. J. Food Prot. 2020, 83, 1181–1197. [Google Scholar] [CrossRef]
- Walker, L.J.; Wallace, R.L.; Smith, J.J.; Graham, T.; Saputra, T.; Symes, S.; Stylianopoulos, A.; Polkinghorne, B.G.; Kirk, M.D.; Glass, K. Prevalence of Campylobacter coli and Campylobacter jejuni in Retail Chicken, Beef, Lamb, and Pork Products in Three Australian States. J. Food Prot. 2019, 82, 2126–2134. [Google Scholar] [CrossRef]
- Madden, R.H.; Moran, L.; Scates, P.; McBride, J.; Kelly, C. Prevalence of Campylobacter and Salmonella in Raw Chicken on Retail Sale in the Republic of Ireland. J. Food Prot. 2011, 74, 1912–1916. [Google Scholar] [CrossRef]
- Korsak, D.; Maćkiw, E.; Rożynek, E.; Żyłowska, M. Prevalence of Campylobacter Spp. in Retail Chicken, Turkey, Pork, and Beef Meat in Poland between 2009 and 2013. J. Food Prot. 2015, 78, 1024–1028. [Google Scholar] [CrossRef]
- Newell, D.G.; Elvers, K.T.; Dopfer, D.; Hansson, I.; Jones, P.; James, S.; Gittins, J.; Stern, N.J.; Davies, R.; Connerton, I.; et al. Biosecurity-Based Interventions and Strategies to Reduce Campylobacter Spp. on Poultry Farms. Appl. Environ. Microbiol. 2011, 77, 8605–8614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royden, A.; Christley, R.; Prendiville, A.; Williams, N.J. The Role of Biosecurity in the Control of Campylobacter: A Qualitative Study of the Attitudes and Perceptions of UK Broiler Farm Workers. Front. Vet. Sci. 2021, 8, 1552. [Google Scholar] [CrossRef] [PubMed]
- Ghareeb, K.; Awad, W.A.; Mohnl, M.; Schatzmayr, G.; Böhm, J. Control Strategies for Campylobacter Infection in Poultry Production. Worlds Poult. Sci. J. 2013, 69, 57–76. [Google Scholar] [CrossRef]
- Meunier, M.; Guyard-Nicodème, M.; Dory, D.; Chemaly, M. Control Strategies against Campylobacter at the Poultry Production Level: Biosecurity Measures, Feed Additives and Vaccination. J. Appl. Microbiol. 2016, 120, 1139–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Hakeem, W.G.; Fathima, S.; Shanmugasundaram, R.; Selvaraj, R.K. Campylobacter jejuni in Poultry: Pathogenesis and Control Strategies. Microorganisms 2022, 10, 2134. [Google Scholar] [CrossRef]
- Lu, T.; Marmion, M.; Ferone, M.; Wall, P.; Scannell, A.G.M. On Farm Interventions to Minimise Campylobacter Spp. Contamination in Chicken. Br. Poult. Sci. 2021, 62, 53–67. [Google Scholar] [CrossRef]
- Wales, A.D.; Vidal, A.B.; Davies, R.H.; Rodgers, J.D. Field Interventions against Colonization of Broilers by Campylobacter. Compr. Rev. Food Sci. Food Saf. 2019, 18, 167–188. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.; Kumar, S.; Oakley, B.; Kim, W.K. Chicken Gut Microbiota: Importance and Detection Technology. Front. Vet. Sci. 2018, 5, 254. [Google Scholar] [CrossRef] [Green Version]
- Rychlik, I. Composition and Function of Chicken Gut Microbiota. Anim. Open Access J. 2020, 10, 103. [Google Scholar] [CrossRef] [Green Version]
- Saint-Cyr, M.J.; Haddad, N.; Taminiau, B.; Poezevara, T.; Quesne, S.; Amelot, M.; Daube, G.; Chemaly, M.; Dousset, X.; Guyard-Nicodème, M. Use of the Potential Probiotic Strain Lactobacillus salivarius SMXD51 to Control Campylobacter jejuni in Broilers. Int. J. Food Microbiol. 2017, 247, 9–17. [Google Scholar] [CrossRef]
- Stern, N.J.; Svetoch, E.A.; Eruslanov, B.V.; Perelygin, V.V.; Mitsevich, E.V.; Mitsevich, I.P.; Pokhilenko, V.D.; Levchuk, V.P.; Svetoch, O.E.; Seal, B.S. Isolation of a Lactobacillus salivarius Strain and Purification of Its Bacteriocin, Which Is Inhibitory to Campylobacter jejuni in the Chicken Gastrointestinal System. Antimicrob. Agents Chemother. 2006, 50, 3111–3116. [Google Scholar] [CrossRef] [Green Version]
- Messaoudi, S.; Kergourlay, G.; Dalgalarrondo, M.; Choiset, Y.; Ferchichi, M.; Prévost, H.; Pilet, M.-F.; Chobert, J.-M.; Manai, M.; Dousset, X. Purification and Characterization of a New Bacteriocin Active against Campylobacter Produced by Lactobacillus salivarius SMXD51. Food Microbiol. 2012, 32, 129–134. [Google Scholar] [CrossRef]
- Helmy, Y.A.; Closs, G.; Jung, K.; Kathayat, D.; Vlasova, A.; Rajashekara, G. Effect of Probiotic E. coli Nissle 1917 Supplementation on the Growth Performance, Immune Responses, Intestinal Morphology, and Gut Microbes of Campylobacter jejuni Infected Chickens. Infect. Immun. 2022, 90, e00337-22. [Google Scholar] [CrossRef]
- Šimunović, K.; Sahin, O.; Erega, A.; Štefanič, P.; Zhang, Q.; Mandic Mulec, I.; Smole Možina, S.; Klančnik, A. Bacillus subtilis PS-216 Spores Supplemented in Broiler Chicken Drinking Water Reduce Campylobacter jejuni Colonization and Increases Weight Gain. Front. Microbiol. 2022, 13, 2551. [Google Scholar] [CrossRef]
- Cisek, A.A.; Binek, M. Chicken Intestinal Microbiota Function with a Special Emphasis on the Role of Probiotic Bacteria. Pol. J. Vet. Sci. 2014, 17, 385–394. [Google Scholar] [CrossRef] [Green Version]
- Guyard-Nicodème, M.; Keita, A.; Quesne, S.; Amelot, M.; Poezevara, T.; Le Berre, B.; Sánchez, J.; Vesseur, P.; Martín, Á.; Medel, P.; et al. Efficacy of Feed Additives against Campylobacter in Live Broilers during the Entire Rearing Period1. Poult. Sci. 2016, 95, 298–305. [Google Scholar] [CrossRef]
- Clavijo, V.; Flórez, M.J.V. The Gastrointestinal Microbiome and Its Association with the Control of Pathogens in Broiler Chicken Production: A Review. Poult. Sci. 2018, 97, 1006–1021. [Google Scholar] [CrossRef]
- Deng, W.; Dittoe, D.K.; Pavilidis, H.O.; Chaney, W.E.; Yang, Y.; Ricke, S.C. Current Perspectives and Potential of Probiotics to Limit Foodborne Campylobacter in Poultry. Front. Microbiol. 2020, 11, 583429. [Google Scholar] [CrossRef]
- Robyn, J.; Rasschaert, G.; Messens, W.; Pasmans, F.; Heyndrickx, M. Screening for Lactic Acid Bacteria Capable of Inhibiting Campylobacter jejuni in In Vitro Simulations of the Broiler Chicken Caecal Environment. Benef. Microbes 2012, 3, 299–308. [Google Scholar] [CrossRef]
- Robyn, J.; Rasschaert, G.; Hermans, D.; Pasmans, F.; Heyndrickx, M. In Vivo Broiler Experiments to Assess Anti-Campylobacter jejuni Activity of a Live Enterococcus Faecalis Strain. Poult. Sci. 2013, 92, 265–271. [Google Scholar] [CrossRef]
- Mortada, M.; Cosby, D.E.; Shanmugasundaram, R.; Selvaraj, R.K. In Vivo and In Vitro Assessment of Commercial Probiotic and Organic Acid Feed Additives in Broilers Challenged with Campylobacter coli. J. Appl. Poult. Res. 2020, 29, 435–446. [Google Scholar] [CrossRef]
- Ty, M.; Taha-Abdelaziz, K.; Demey, V.; Castex, M.; Sharif, S.; Parkinson, J. Performance of Distinct Microbial Based Solutions in a Campylobacter Infection Challenge Model in Poultry. Anim. Microbiome 2022, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Sahin, O.; Pang, J.; Tang, Y.; Adiguzel, M.C.; Wang, C.; Zhang, Q. A Longitudinal Study on Campylobacter in Conventional Commercial Broiler Flocks in the United States: Prevalence and Genetic Diversity. Avian Dis. 2023; submitted. [Google Scholar]
- Diaz Carrasco, J.M.; Casanova, N.A.; Fernández Miyakawa, M.E. Microbiota, Gut Health and Chicken Productivity: What Is the Connection? Microorganisms 2019, 7, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, J.J.; Bobeck, E.A.; Sylte, M.J.; Looft, T. Eggshell and Environmental Bacteria Contribute to the Intestinal Microbiota of Growing Chickens. J. Anim. Sci. Biotechnol. 2020, 11, 60. [Google Scholar] [CrossRef]
- Ding, J.; Dai, R.; Yang, L.; He, C.; Xu, K.; Liu, S.; Zhao, W.; Xiao, L.; Luo, L.; Zhang, Y.; et al. Inheritance and Establishment of Gut Microbiota in Chickens. Front. Microbiol. 2017, 8, 1967. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing Taxonomic Classification of Marker-Gene Amplicon Sequences with QIIME 2’s Q2-Feature-Classifier Plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Robeson, M.S.; O’Rourke, D.R.; Kaehler, B.D.; Ziemski, M.; Dillon, M.R.; Foster, J.T.; Bokulich, N.A. RESCRIPt: Reproducible Sequence Taxonomy Reference Database Management for the Masses. PLoS Comput. Biol. 2021, 17, e1009581. [Google Scholar]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’hara, B.G.L.S.; Solymos, P.M.H.S.; Wagner, H. Vegan: Community Ecology Package, R Package Version 1. 15-4; R Foundation for Statistical Computing: Vienna, Austria, 2009.
- Lin, H.; Peddada, S.D. Analysis of Compositions of Microbiomes with Bias Correction. Nat. Commun. 2020, 11, 3514. [Google Scholar] [CrossRef] [PubMed]
- Hertogs, K.; Haegeman, A.; Schaumont, D.; Gelaude, P.; De Zutter, L.; Dewulf, J.; Heyndrickx, M.; Rasschaert, G. Contamination Sources and Transmission Routes for Campylobacter on (Mixed) Broiler Farms in Belgium, and Comparison of the Gut Microbiota of Flocks Colonized and Uncolonized with Campylobacter. Pathogens 2021, 10, 66. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Zhou, Q.; Yuan, Z.; Fu, L.; Wen, C.; Yang, N.; Sun, C. Impact of the Gut Microecology on Campylobacter Presence Revealed by Comparisons of the Gut Microbiota from Chickens Raised on Litter or in Individual Cages. BMC Microbiol. 2021, 21, 290. [Google Scholar] [CrossRef] [PubMed]
- Connerton, P.L.; Richards, P.J.; Lafontaine, G.M.; O’Kane, P.M.; Ghaffar, N.; Cummings, N.J.; Smith, D.L.; Fish, N.M.; Connerton, I.F. The Effect of the Timing of Exposure to Campylobacter jejuni on the Gut Microbiome and Inflammatory Responses of Broiler Chickens. Microbiome 2018, 6, 88. [Google Scholar] [CrossRef]
- Chintoan-Uta, C.; Wisedchanwet, T.; Glendinning, L.; Bremner, A.; Psifidi, A.; Vervelde, L.; Watson, K.; Watson, M.; Stevens, M.P. Role of Cecal Microbiota in the Differential Resistance of Inbred Chicken Lines to Colonization by Campylobacter jejuni. Appl. Environ. Microbiol. 2020, 86, e02607-19. [Google Scholar] [CrossRef]
- Pan, D.; Yu, Z. Intestinal Microbiome of Poultry and Its Interaction with Host and Diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Patuzzi, I.; Orsini, M.; Cibin, V.; Petrin, S.; Mastrorilli, E.; Tiengo, A.; Gobbo, F.; Catania, S.; Barco, L.; Ricci, A.; et al. The Interplay between Campylobacter and the Caecal Microbial Community of Commercial Broiler Chickens over Time. Microorganisms 2021, 9, 221. [Google Scholar] [CrossRef]
- McKenna, A.; Ijaz, U.Z.; Kelly, C.; Linton, M.; Sloan, W.T.; Green, B.D.; Lavery, U.; Dorrell, N.; Wren, B.W.; Richmond, A.; et al. Impact of Industrial Production System Parameters on Chicken Microbiomes: Mechanisms to Improve Performance and Reduce Campylobacter. Microbiome 2020, 8, 128. [Google Scholar] [CrossRef]
- Sofka, D.; Pfeifer, A.; Gleiss, B.; Paulsen, P.; Hilbert, F. Changes within the Intestinal Flora of Broilers by Colonisation with Campylobacter jejuni. Berl. Münchener Tierärztliche Wochenschr. 2015, 128, 104–110. [Google Scholar]
- Kers, J.G.; Velkers, F.C.; Fischer, E.A.J.; Hermes, G.D.A.; Stegeman, J.A.; Smidt, H. Host and Environmental Factors Affecting the Intestinal Microbiota in Chickens. Front. Microbiol. 2018, 9, 235. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Morrison, M.; Yu, Z. Bacterial Census of Poultry Intestinal Microbiome. Poult. Sci. 2013, 92, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Wise, M.G.; Siragusa, G.R. Quantitative Analysis of the Intestinal Bacterial Community in One- to Three-Week-Old Commercially Reared Broiler Chickens Fed Conventional or Antibiotic-Free Vegetable-Based Diets. J. Appl. Microbiol. 2007, 102, 1138–1149. [Google Scholar] [CrossRef] [PubMed]
- Oakley, B.B.; Morales, C.A.; Line, J.; Berrang, M.E.; Meinersmann, R.J.; Tillman, G.E.; Wise, M.G.; Siragusa, G.R.; Hiett, K.L.; Seal, B.S. The Poultry-Associated Microbiome: Network Analysis and Farm-to-Fork Characterizations. PLoS ONE 2013, 8, e57190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.T.; Duncan, S.H.; Stams, A.J.M.; van Dijl, J.M.; Flint, H.J.; Harmsen, H.J.M. The Gut Anaerobe Faecalibacterium prausnitzii Uses an Extracellular Electron Shuttle to Grow at Oxic–Anoxic Interphases. ISME J. 2012, 6, 1578–1585. [Google Scholar] [CrossRef]
- Hao, Y.; Ji, Z.; Shen, Z.; Xue, Y.; Zhang, B.; Yu, D.; Liu, T.; Luo, D.; Xing, G.; Tang, J.; et al. Increase Dietary Fiber Intake Ameliorates Cecal Morphology and Drives Cecal Species-Specific of Short-Chain Fatty Acids in White Pekin Ducks. Front. Microbiol. 2022, 13, 853797. [Google Scholar] [CrossRef] [PubMed]
- Donohoe, D.R.; Garge, N.; Zhang, X.; Sun, W.; O’Connell, T.M.; Bunger, M.K.; Bultman, S.J. The Microbiome and Butyrate Regulate Energy Metabolism and Autophagy in the Mammalian Colon. Cell Metab. 2011, 13, 517–526. [Google Scholar] [CrossRef] [Green Version]
- Corrêa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A.R. Regulation of Immune Cell Function by Short-Chain Fatty Acids. Clin. Transl. Immunol. 2016, 5, e73. [Google Scholar] [CrossRef]
- Kang, R.; Wang, W.; Liu, Y.; Huang, S.; Xu, J.; Zhao, L.; Zhang, J.; Ji, C.; Wang, Z.; Hu, Y.; et al. Dietary Selenium Sources Alleviate Immune Challenge Induced by Salmonella Enteritidis Potentially through Improving the Host Immune Response and Gut Microbiota in Laying Hens. Front. Immunol. 2022, 13, 928865. [Google Scholar] [CrossRef]
- Balta, I.; Butucel, E.; Stef, L.; Pet, I.; Gradisteanu-Pircalabioru, G.; Chifiriuc, C.; Gundogdu, O.; McCleery, D.; Corcionivoschi, N. Anti-Campylobacter Probiotics: Latest Mechanistic Insights. Foodborne Pathog. Dis. 2022, 19, 693–703. [Google Scholar] [CrossRef]
- Taha-Abdelaziz, K.; Astill, J.; Kulkarni, R.R.; Read, L.R.; Najarian, A.; Farber, J.M.; Sharif, S. In Vitro Assessment of Immunomodulatory and Anti-Campylobacter Activities of Probiotic Lactobacilli. Sci. Rep. 2019, 9, 17903. [Google Scholar] [CrossRef] [Green Version]
- Messaoudi, S.; Kergourlay, G.; Rossero, A.; Ferchichi, M.; Prévost, H.; Drider, D.; Manai, M.; Dousset, X. Identification of Lactobacilli Residing in Chicken Ceca with Antagonism against Campylobacter. Int. Microbiol. 2011, 14, 103. [Google Scholar] [CrossRef]
- Perez-Perez, G.I.; Blaser, M.J. Campylobacter and Helicobacter. In Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; ISBN 978-0-9631172-1-2. [Google Scholar]
- Kaakoush, N.O.; Sodhi, N.; Chenu, J.W.; Cox, J.M.; Riordan, S.M.; Mitchell, H.M. The Interplay between Campylobacter and Helicobacter Species and Other Gastrointestinal Microbiota of Commercial Broiler Chickens. Gut Pathog. 2014, 6, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Guo, X.; Zhang, J.; Zhang, M.; Ou, Z.; Peng, Y. Phascolarctobacterium faecium Abundant Colonization in Human Gastrointestinal Tract. Exp. Ther. Med. 2017, 14, 3122–3126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heimesaat, M.M.; Weschka, D.; Mousavi, S.; Bereswill, S. Treatment with the Probiotic Product Aviguard® Alleviates Inflammatory Responses during Campylobacter jejuni-Induced Acute Enterocolitis in Mice. Int. J. Mol. Sci. 2021, 22, 6683. [Google Scholar] [CrossRef] [PubMed]
- Valeris-Chacin, R.; Pieters, M.; Hwang, H.; Johnson, T.J.; Singer, R.S. Association of Broiler Litter Microbiome Composition and Campylobacter Isolation. Front. Vet. Sci. 2021, 8, 654927. [Google Scholar] [CrossRef]
- Valeris-Chacin, R.; Weber, B.; Johnson, T.J.; Pieters, M.; Singer, R.S. Longitudinal Changes in Campylobacter and the Litter Microbiome throughout the Broiler Production Cycle. Appl. Environ. Microbiol. 2022, 88, e00667-22. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Farm ID * | Campylobacter Status (2012–2014) | Campylobacter Status (2018–2019) | No. Samples Used in This Study | No. Samples Tested Positive in This Study |
|---|---|---|---|---|
| BB | Positive | Positive | 80 | 76 |
| CL | Negative | Positive | 80 | 79 |
| DF | Negative | Positive | 80 | 80 |
| BP | Positive | Negative | 80 | 0 |
| KP | Positive | Negative | 80 | 0 |
| WL | Negative | Negative | 120 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pang, J.; Looft, T.; Zhang, Q.; Sahin, O. Deciphering the Association between Campylobacter Colonization and Microbiota Composition in the Intestine of Commercial Broilers. Microorganisms 2023, 11, 1724. https://doi.org/10.3390/microorganisms11071724
Pang J, Looft T, Zhang Q, Sahin O. Deciphering the Association between Campylobacter Colonization and Microbiota Composition in the Intestine of Commercial Broilers. Microorganisms. 2023; 11(7):1724. https://doi.org/10.3390/microorganisms11071724
Chicago/Turabian StylePang, Jinji, Torey Looft, Qijing Zhang, and Orhan Sahin. 2023. "Deciphering the Association between Campylobacter Colonization and Microbiota Composition in the Intestine of Commercial Broilers" Microorganisms 11, no. 7: 1724. https://doi.org/10.3390/microorganisms11071724