
The Effect of Three Complexes of Iodine with Amino Acids on Gene Expression of Model Antibiotic Resistant Microorganisms Escherichia coli ATCC BAA-196 and Staphylococcus aureus ATCC BAA-39
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Iodine-Containing Compounds
2.2. Test Strains and Growth Conditions
2.3. X-ray Diffractions Analysis
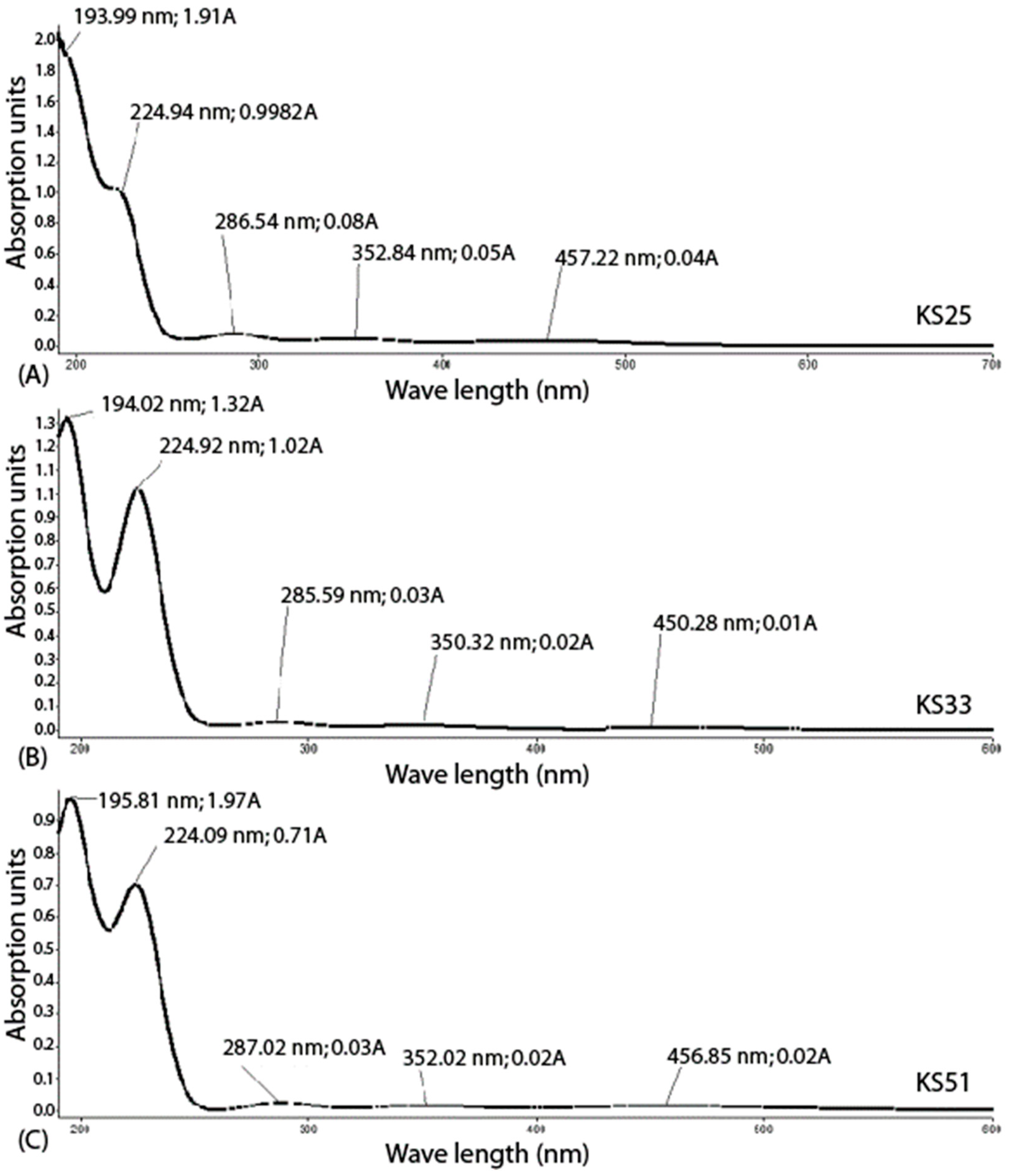
2.4. UV Spectroscopy
2.5. Determination of Minimal Bactericidal Concentrations of the Iodine Containing Complexes
2.6. Treatment of the Bacterial Cultures with the Complexes
2.7. RNA Library Preparation and Sequencing
2.8. Differential Gene Expression Analyses
2.9. Metabolic Pathway and Regulatory Network Analyses
2.10. Statistical Evaluations
3. Results
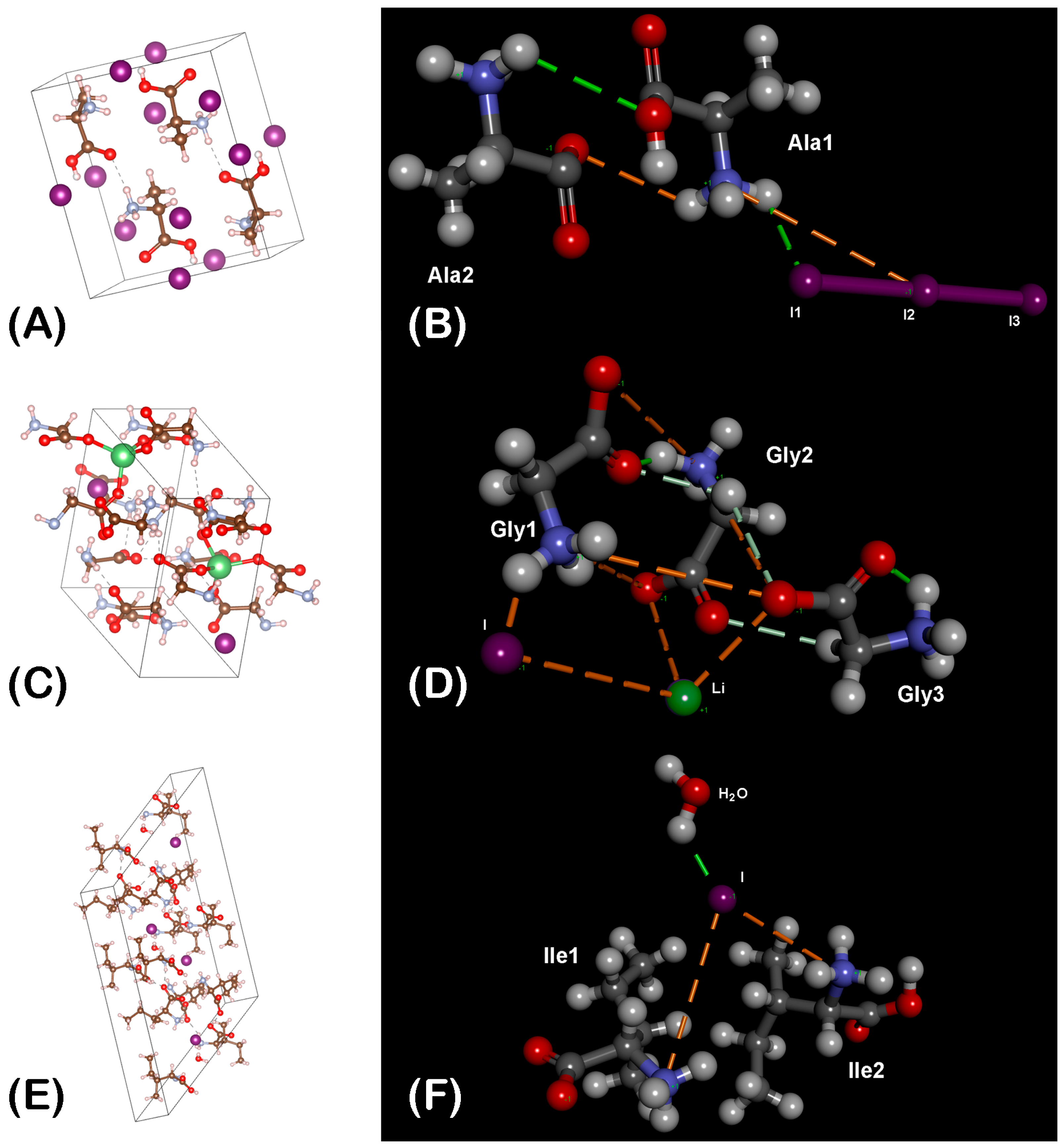
3.1. Prediction of Chemical Structures of the Synthesized Complexes
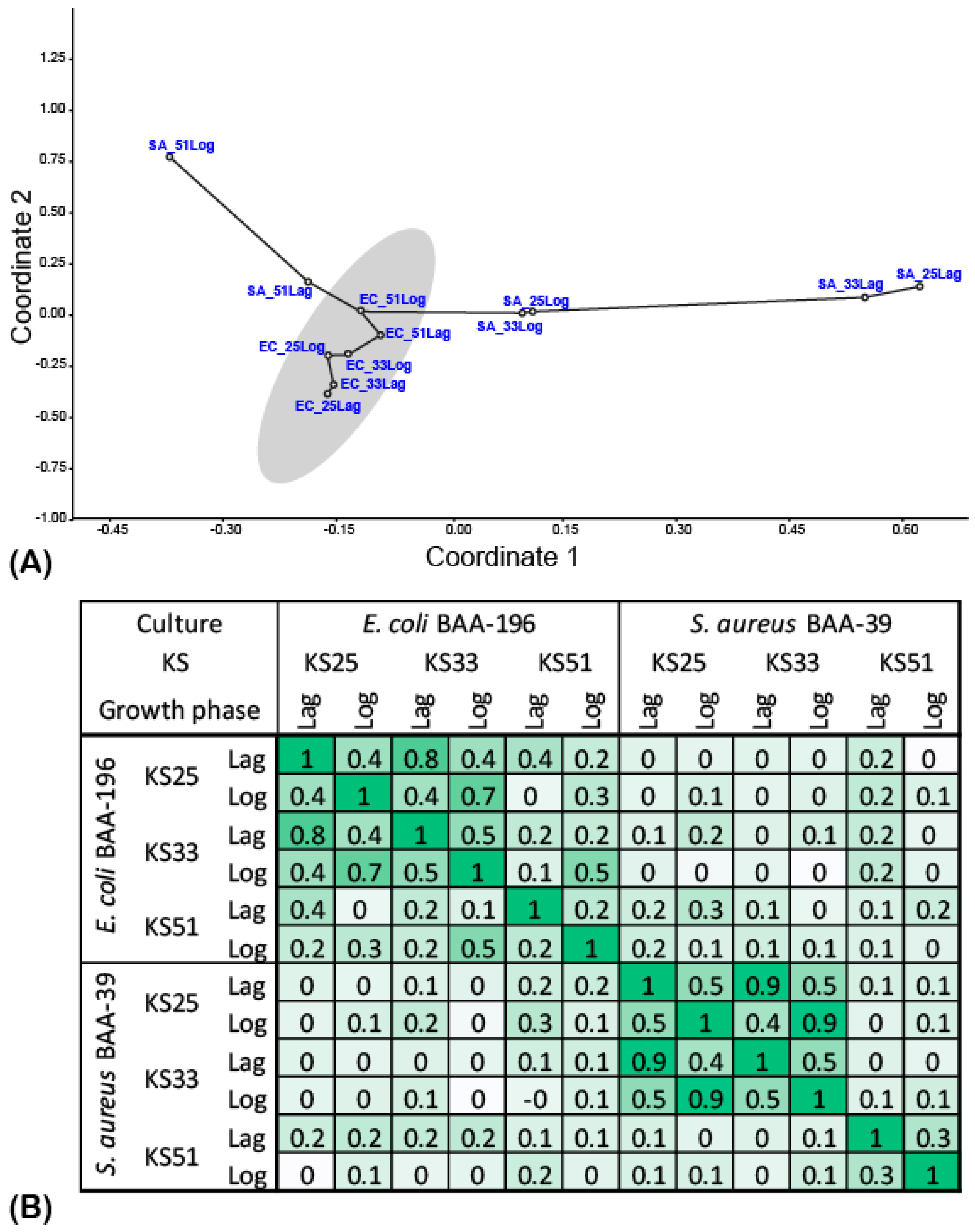
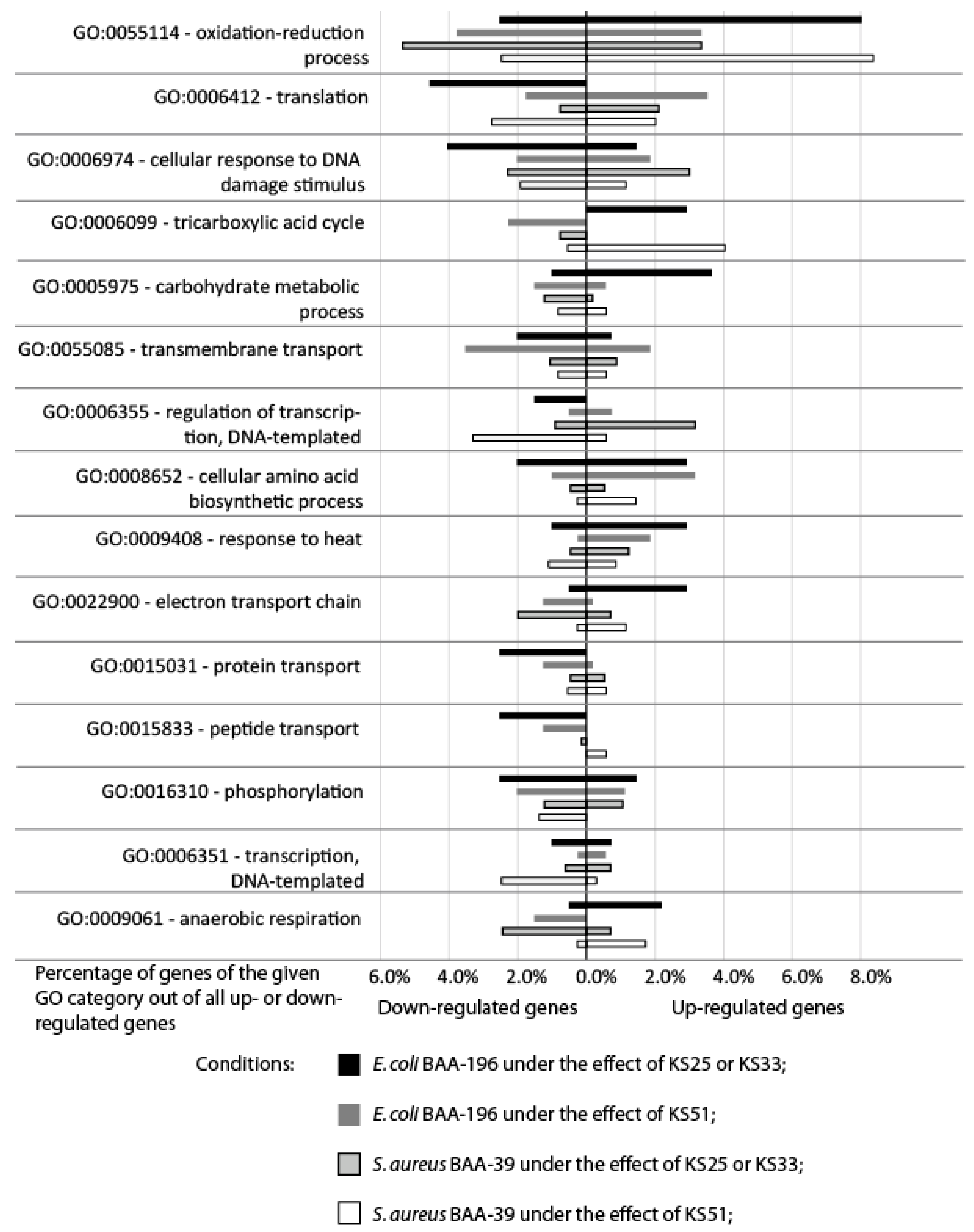
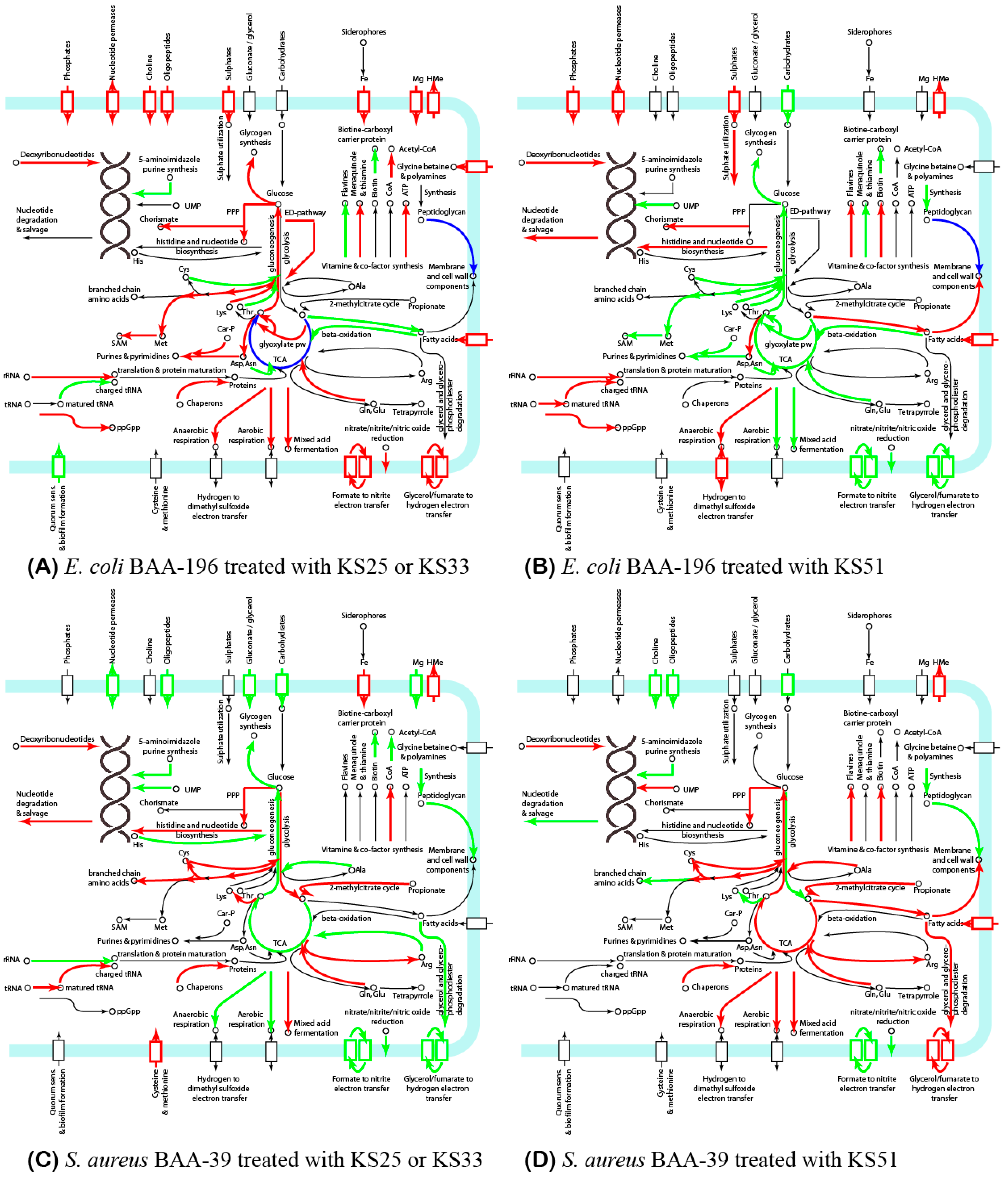
3.2. Comparison of Gene Expression Patterns in E. coli and S. aureus under the Effect of Iodine-Containing Complexes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Laxminarayan, R. Antibiotic effectiveness: Balancing conservation against innovation. Science 2014, 345, 1299–1301. [Google Scholar] [CrossRef] [Green Version]
- Årdal, C.; Balasegaram, M.; Laxminarayan, R.; McAdams, D.; Outterson, K.; Rex, J.H.; Sumpradit, N. Antibiotic development—Economic, regulatory and societal challenges. Nat. Rev. Microbiol. 2020, 18, 267–274. [Google Scholar] [CrossRef]
- Bush, K.; Courvalin, P.; Dantas, G.; Davies, J.; Eisenstein, B.; Huovinen, P.; Jacoby, G.A.; Kishony, R.; Kreiswirth, B.N.; Kutter, E.; et al. Tackling antibiotic resistance. Nat. Rev. Microbiol. 2011, 9, 894–896. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Jia, Y.; Yang, K.; Li, R.; Xiao, X.; Zhu, K.; Wang, Z. Metformin restores tetracyclines susceptibility against multidrug resistant bacteria. Adv. Sci. 2020, 7, 1902227. [Google Scholar] [CrossRef]
- McClure, N.S.; Day, T. A theoretical examination of the relative importance of evolution management and drug development for managing resistance. Proc. Biol. Sci. 2014, 281, 20141861. [Google Scholar] [CrossRef]
- Ilin, A.I.; Kulmanov, M.E.; Korotetskiy, I.S.; Islamov, R.A.; Akhmetova, G.K.; Lankina, M.V.; Reva, O.N. Genomic insight into mechanisms of reversion of antibiotic resistance in multidrug resistant Mycobacterium tuberculosis induced by a nanomolecular iodine-containing complex FS-1. Front. Cell Infect. Microbiol. 2017, 7, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reva, O.N.; Korotetskiy, I.S.; Joubert, M.; Shilov, S.V.; Jumagaziyeva, A.B.; Suldina, N.A.; Ilin, A.I. The effect of iodine-containing nano-micelles, FS-1, on antibiotic resistance, gene expression and epigenetic modifications in the genome of multidrug resistant MRSA strain Staphylococcus aureus ATCC BAA-39. Front. Microbiol. 2020, 11, 581660. [Google Scholar] [CrossRef] [PubMed]
- Korotetskiy, I.S.; Jumagaziyeva, A.B.; Shilov, S.V.; Kuznetsova, T.V.; Myrzabayeva, A.N.; Iskakbayeva, Z.A.; Ilin, A.I.; Joubert, M.; Taukobong, S.; Reva, O.N. Transcriptomics and methylomics study on the effect of iodine-containing drug FS-1 on Escherichia coli ATCC BAA-196. Future Microbiol. 2021, 16, 1063–1085. [Google Scholar]
- Korotetskiy, I.S.; Shilov, S.V.; Kuznetsova, T.V.; Ilin, A.I.; Joubert, M.; Taukobong, S.; Reva, O.N. Comparison of transcriptional responses and metabolic alterations in three multidrug-resistant model microorganisms, Staphylococcus aureus ATCC BAA-39, Escherichia coli ATCC BAA-196, and Acinetobacter baumannii ATCC BAA-1790, on exposure to iodine-containing nano-micelle drug FS-1. mSystems 2021, 6, e01293-20. [Google Scholar] [CrossRef]
- Abelson, M.; Capriotti, J.A.; Shapiro, A.; Lilyestrom, L. Iodine: An elemental force against infection. Rev. Ophthalmol. 2009, 15, 36–42. [Google Scholar]
- Pavlík, V.; Sojka, M.; Mazúrová, M.; Velebný, V. Dual role of iodine, silver, chlorhexidine and octenidine as antimicrobial and antiprotease agents. PLoS ONE 2019, 14, e0211055. [Google Scholar] [CrossRef]
- Islamov, R.; Kustova, T.; Nersesyan, A.; Ilin, A. Subchronic toxicity of the new iodine complex in dogs and rats. Front. Vet. Sci. 2020, 7, 184. [Google Scholar] [CrossRef] [Green Version]
- Murdoch, R.; Lagan, K.M. The role of povidone and cadexomer iodine in the management of acute and chronic wounds. Phys. Ther. Rev. 2013, 18, 207–216. [Google Scholar] [CrossRef]
- Wei, X.; Chen, X.; Wang, X.; Zheng, W.; Zhang, D.; Tian, D.; Jiang, S.; Ong, C.N.; He, G.; Qu, W. Occurrence of regulated and emerging iodinated DBPs in the Shanghai drinking water. PLoS ONE 2013, 8, e59677. [Google Scholar] [CrossRef] [Green Version]
- Navikaitė, V.; Danilovas, P.P.; Klimavičiūtė, R.; Bendoraitienė, J. The stability of water-soluble modified starch iodophors. Chem. Technol. 2013, 63, 34–41. [Google Scholar] [CrossRef]
- Willson, V.A. Determination of available chlorine in hypochlorite solutions by direct titration with sodium thiosulfate. Ind. Eng. Chem. Anal. Ed. 1935, 7, 44–45. [Google Scholar] [CrossRef]
- Bombi, G.G.; Sacchetto, G.A.; Maccà, C. Solubility equilibrium of silver (I) oxide in molten lithium nitrate+ potassium nitrate mixtures. J. Chem. Soc. Faraday Trans. Phys. Chem. Condens. Phases 1978, 74, 1–9. [Google Scholar] [CrossRef]
- Spek, A.L. PLATON SQUEEZE: A tool for the calculation of the disordered solvent contribution to the calculated structure factors. Acta Crystallogr. C Struct. Chem. 2015, 71 Pt 1, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Howden, B.P.; Beaume, M.; Harrison, P.F.; Hernandez, D.; Schrenzel, J.; Seemann, T.; Francois, P.; Stinear, T.P. Analysis of the small RNA transcriptional response in multidrug-resistant Staphylococcus aureus after antimicrobial exposure. Antimicrob. Agents Chemother. 2013, 57, 3864–3874. [Google Scholar] [CrossRef] [Green Version]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; the UGENE team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Karp, P.D.; Paley, S.; Romero, P. The Pathway Tools software. Bioinformatics 2002, 18 (Suppl. S1), S225–S232. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Moreira, B.; Vinuesa, P. GET_HOMOLOGUES, a versatile software package for scalable and robust microbial pangenome analysis. Appl. Environ. Microbiol. 2013, 79, 7696–7701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Yuldasheva, G.A.; Zhidomirov, G.M.; Leszczynski, J.; Ilin, A.I. The effect of the amphoteric properties of amino acids in the zwitterionic form on the structure of iodine complex compounds in aqueous solutions containing halogenides of alkaline metals and amino acids. J. Mol. Struct. 2013, 1033, 321–330. [Google Scholar] [CrossRef]
- Yuldasheva, G.A.; Zhidomirov, G.M.; Leszczynski, J.; Ilin, A.I. Iodine Containing Drugs: Complexes of Molecular Iodine and Tri-iodide with Bioorganic Ligands and Lithium Halogenides in Aqueous Solutions. In Practical Aspects of Computational Chemistry IV. Chapter 10; Leszczynski, J., Shukla, M., Eds.; Springer Science & Business Media: Berlin, Germany, 2016; pp. 279–301. [Google Scholar]
- Nagaoka, M.; Okuyama-Yoshida, N.; Yamabe, T. Origin of the transition state on the free energy surface: Intramolecular proton transfer reaction of glycine in aqueous solution. J. Phys. Chem. A 1998, 102, 8202–8208. [Google Scholar] [CrossRef]
- Ackerley, D.F.; Gonzalez, C.F.; Keyhan, M.; Blake, R., 2nd; Matin, A. Mechanism of chromate reduction by the Escherichia coli protein, NfsA, and the role of different chromate reductases in minimizing oxidative stress during chromate reduction. Environ. Microbiol. 2004, 6, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Chaffin, D.O.; Taylor, D.; Skerrett, S.J.; Rubens, C.E. Changes in the Staphylococcus aureus transcriptome during early adaptation to the lung. PLoS ONE 2012, 7, e41329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duval, V.; Lister, I.M. MarA, SoxS and Rob of Escherichia coli—Global regulators of multidrug resistance, virulence and stress response. Int. J. Biotechnol. Wellness Ind. 2013, 2, 101. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; He, Y.; Irwin, P.L.; Jin, T.; Shi, X. Antibacterial activity and mechanism of action of zinc oxide nanoparticles against Campylobacter jejuni. Appl. Environ. Microbiol. 2011, 77, 2325–2331. [Google Scholar] [CrossRef] [Green Version]
- Gurmu, D.; Lu, J.; Johnson, K.A.; Nordlund, P.; Holmgren, A.; Erlandsen, H. The crystal structure of the protein YhaK from Escherichia coli reveals a new subclass of redox sensitive enterobacterial bicupins. Proteins 2009, 74, 18–31. [Google Scholar] [CrossRef]
- Rai, S.; Rai, R.; Singh, P.K.; Rai, L.C. Alr2321, a multiple stress inducible glyoxalase I of Anabaena sp. PCC7120 detoxifies methylglyoxal and reactive species oxygen. Aquat. Toxicol. 2019, 214, 105238. [Google Scholar] [CrossRef] [PubMed]
- Masri, A.; Khan, N.A.; Zoqratt, M.Z.H.M.; Ayub, Q.; Anwar, A.; Rao, K.; Shah, M.R.; Siddiqui, R. Transcriptome analysis of Escherichia coli K1 after therapy with hesperidin conjugated with silver nanoparticles. BMC Microbiol. 2021, 21, 51. [Google Scholar] [CrossRef] [PubMed]
- Grossi, J.; Kohanoff, J.J.; English, N.J.; Bringa, E.M.; Del Pópolo, M.G. On the mechanism of the iodide-triiodide exchange reaction in a solid-state ionic liquid. J. Phys. Chem. 2017, 121, 6436–6441. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complex | Source of Iodine | Amino Acid |
|---|---|---|
| KS25 | I2 | Alanine |
| KS33 | I2 + LiI | Glycine |
| KS51 | I2 + LiI | Isoleucine |
| Test-Cultures | MBC Values (mg/mL), I2 Content Concentration per Total Concentration of the Substance | ||
|---|---|---|---|
| KS25 | KS33 | KS51 | |
| E. coli BAA-196 | 0.50/4.0 | 0.32/2.0 | 0.57/2.0 |
| S. aureus BAA-39 | 0.50/4.0 | 0.32/2.0 | 0.57/2.0 |
| Strain | Growth Phase | Treatment with the Complexes | Negative Control (NC) | ||
|---|---|---|---|---|---|
| KS25 | KS33 | KS51 | |||
| E. coli BAA-196 | Lag | 3 | 3 | 3 | 6 |
| Log | 3 | 3 | 3 | 6 | |
| S. aureus BAA-39 | Lag | 3 | 3 | 3 | 6 |
| Log | 6 | 6 | 3 | 9 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kenesheva, S.T.; Taukobong, S.; Shilov, S.V.; Kuznetsova, T.V.; Jumagaziyeva, A.B.; Karpenyuk, T.A.; Reva, O.N.; Ilin, A.I. The Effect of Three Complexes of Iodine with Amino Acids on Gene Expression of Model Antibiotic Resistant Microorganisms Escherichia coli ATCC BAA-196 and Staphylococcus aureus ATCC BAA-39. Microorganisms 2023, 11, 1705. https://doi.org/10.3390/microorganisms11071705
Kenesheva ST, Taukobong S, Shilov SV, Kuznetsova TV, Jumagaziyeva AB, Karpenyuk TA, Reva ON, Ilin AI. The Effect of Three Complexes of Iodine with Amino Acids on Gene Expression of Model Antibiotic Resistant Microorganisms Escherichia coli ATCC BAA-196 and Staphylococcus aureus ATCC BAA-39. Microorganisms. 2023; 11(7):1705. https://doi.org/10.3390/microorganisms11071705
Chicago/Turabian StyleKenesheva, Sabina T., Setshaba Taukobong, Sergey V. Shilov, Tatyana V. Kuznetsova, Ardak B. Jumagaziyeva, Tatyana A. Karpenyuk, Oleg N. Reva, and Aleksandr I. Ilin. 2023. "The Effect of Three Complexes of Iodine with Amino Acids on Gene Expression of Model Antibiotic Resistant Microorganisms Escherichia coli ATCC BAA-196 and Staphylococcus aureus ATCC BAA-39" Microorganisms 11, no. 7: 1705. https://doi.org/10.3390/microorganisms11071705