
Assessing the Feasibility of Employing a Combination of a Bacteriophage-Derived Endolysin and Spore Germinants to Treat Relapsing Clostridioides difficile Infection

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. C. difficile Spore Preparation
2.3. Cloning of LysCD6356 and Its EAD
2.4. Assessing the Lytic Activity of EAD
2.5. The Impact of pH on EAD Activity
2.6. The Impact of Calcium on EAD Activity
2.7. The Contribution of Divalent Metal Cations to EAD Activity
2.8. Impact of Spore Germinants and Calcium Chloride on the Lytic Activity of the EAD
2.9. C. difficile Spore Germination
2.10. Assessing the Sensitivity of Newly Germinated Spores to the EAD
2.10.1. Simultaneous Exposure to Spore Germinants and the EAD
2.10.2. Sequential Exposure to Spore Germinants and the EAD
2.11. Phase-Contrast Microscopy
2.12. Scanning Electron Microscopy
2.13. Statistical Analysis
3. Results
3.1. Production of Full-Length Recombinant Lysin of CD6356 and Its EAD Fragment from an E. coli Expression Host
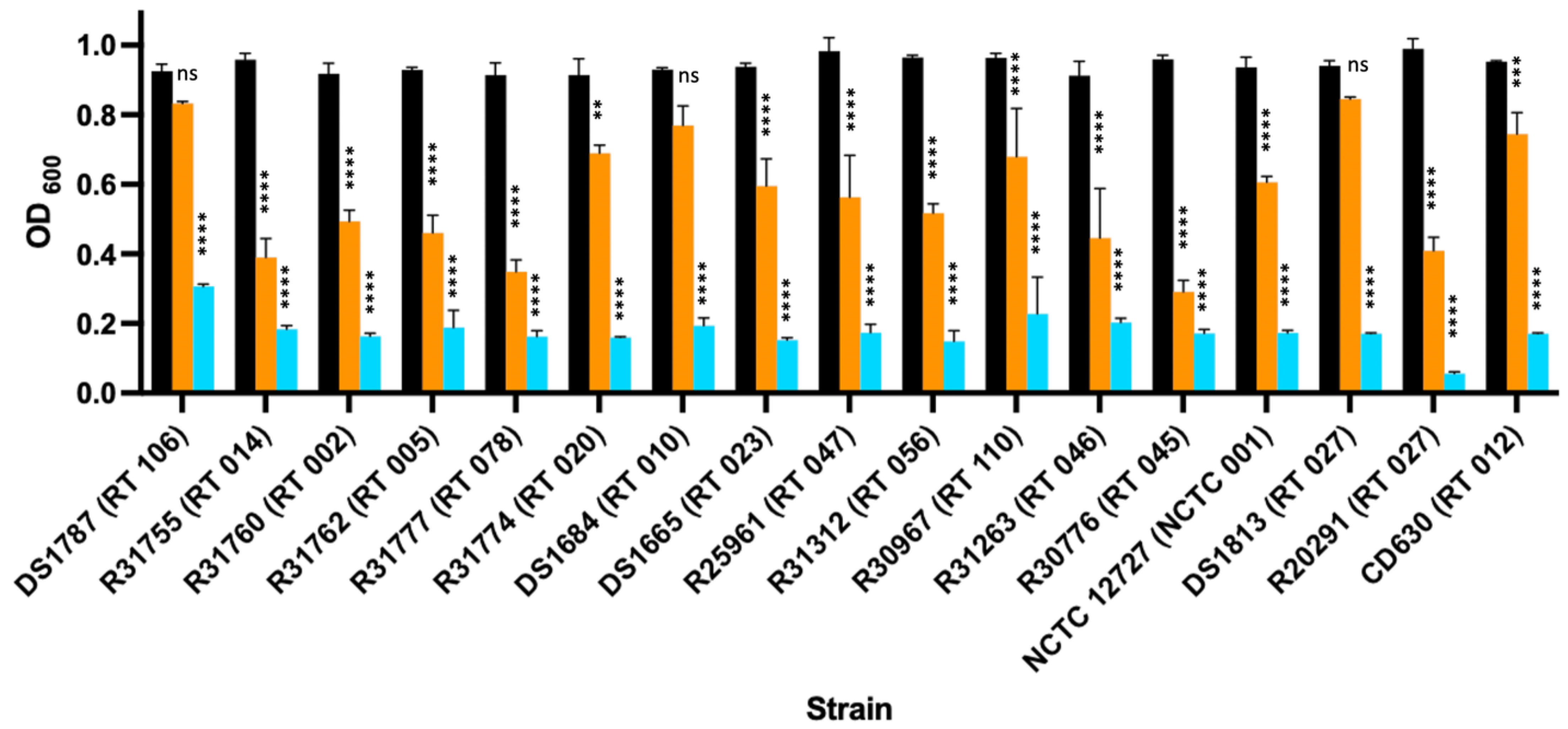
3.2. Enzymic Activity of LysCD6356 and Its EAD Fragment against Vegetative Isolates of C. difficile
3.3. Effect of pH on Lytic Activity
3.4. The Role of Divalent Cations in the Enzymic Activity of the EAD
3.5. The Impact of Calcium on the Enzymic Activity of the EAD of LysCD6356
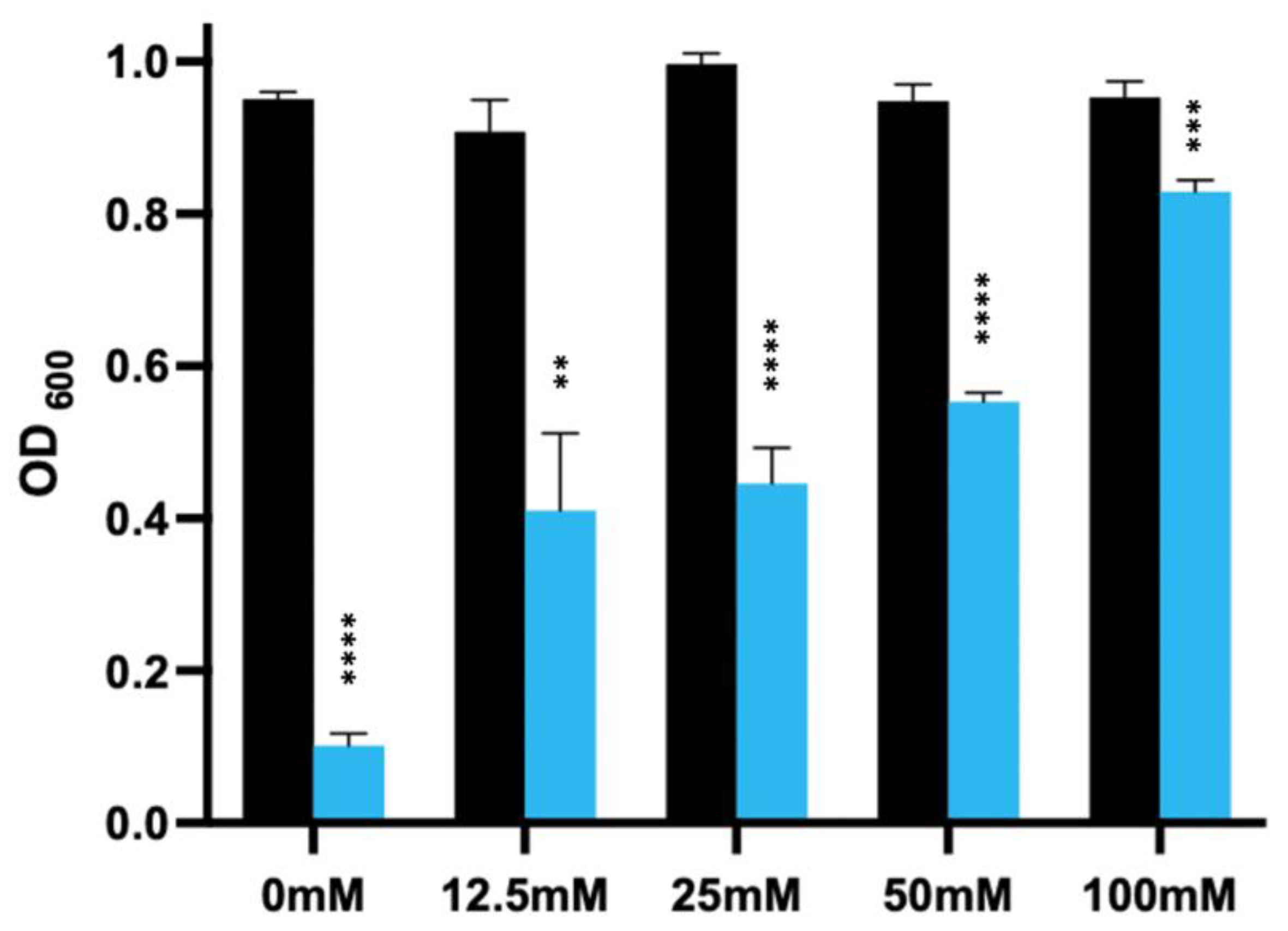
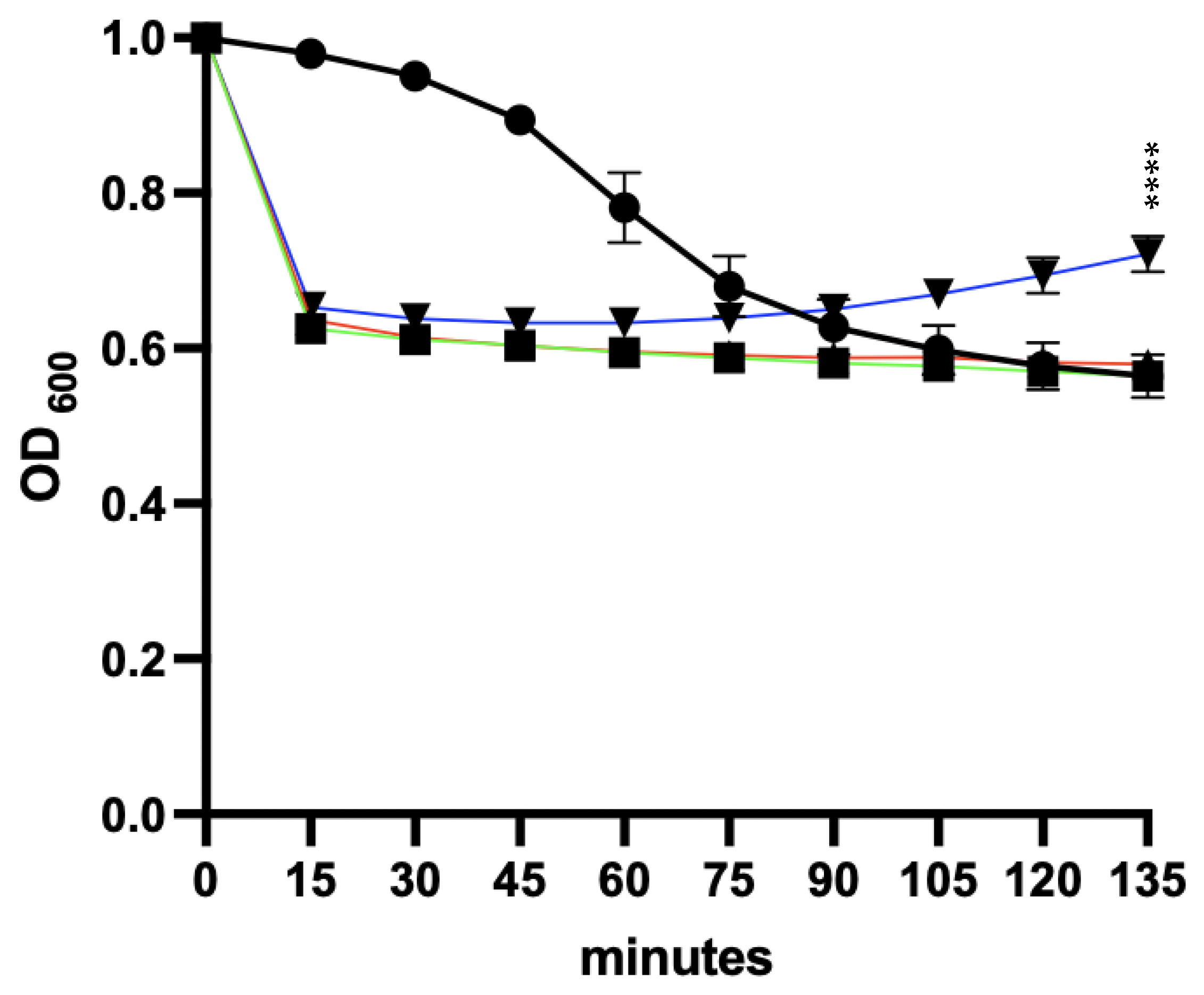
3.6. Effect of Spore Germinants and Calcium on the Activity of the EAD against Vegetative R20291
3.7. Characterisation of Bacterial Structure Using Scanning Electron Microscopy
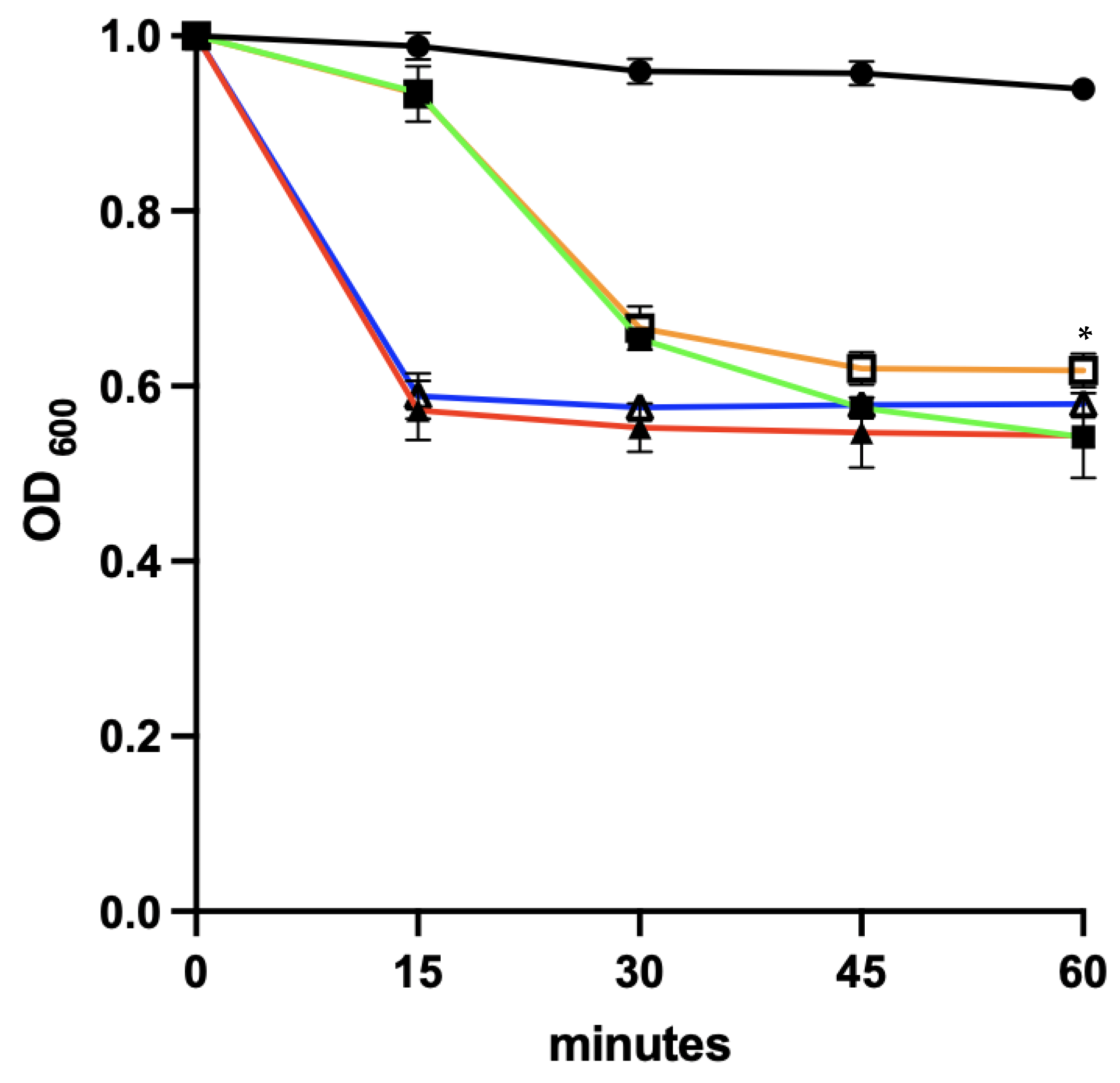
3.8. Spore Germination
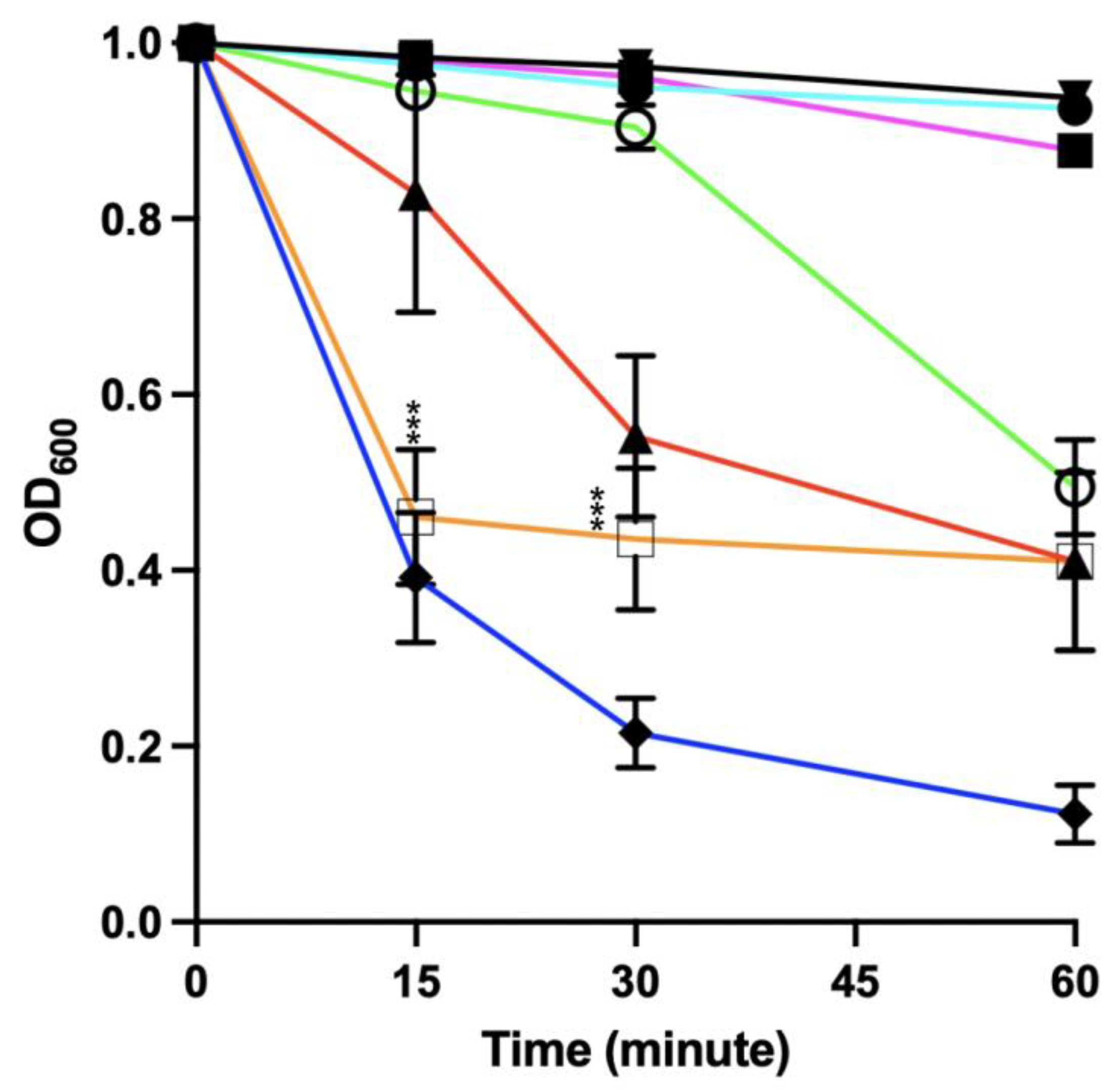
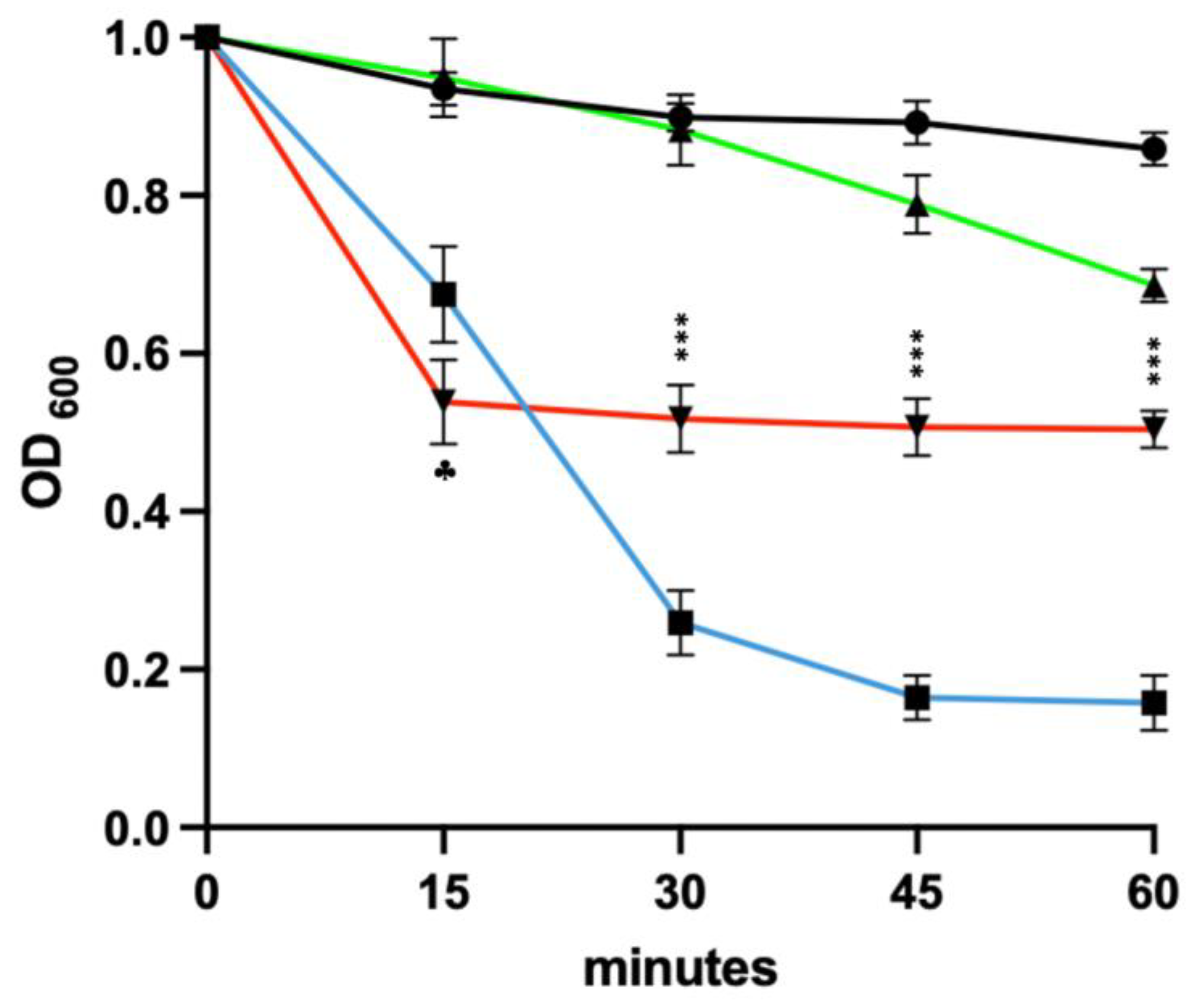
3.9. Sensitivity of Newly Germinated C. difficile to Lysis by the EAD
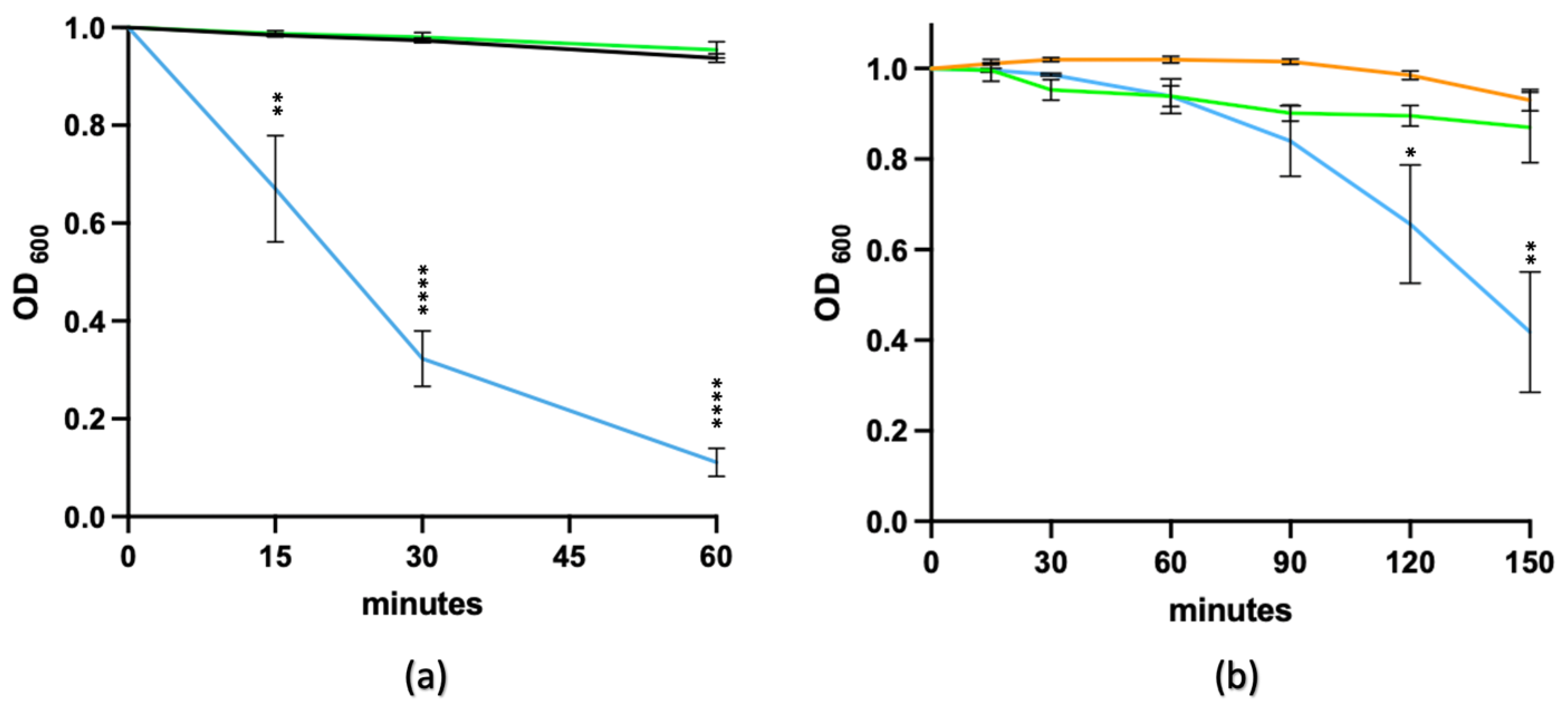
3.9.1. Co-Delivery of Germinants and the EAD
3.9.2. Germinants Followed by the EAD
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Voth, D.E.; Ballard, J.D. Clostridium difficile toxins: Mechanism of action and role in disease. Clin. Microbiol. Rev. 2005, 18, 247–263. [Google Scholar] [CrossRef] [Green Version]
- Dawson, L.F.; Valiente, E.; Donahue, E.H.; Birchenough, G.; Wren, B.W. Hypervirulent clostridium difficile pcr-ribotypes exhibit resistance to widely used disinfectants. PLoS ONE 2011, 6, e25754. [Google Scholar] [CrossRef] [Green Version]
- Frieden Tom. Antibiotic Resistance Threats in the United States 2013; CDC: Atlanta, GA, USA, 2013. [Google Scholar]
- Office for National Statistics. Deaths Involving Clostridium difficile, England and Wales 2012. 2014. Available online: http://www.ons.gov.uk/peoplepopulationandcommunity/birthsdeathsandmarriages/deaths/bulletins/deathsinvolvingclostridiumdifficileenglandandwales/2013-08-22 (accessed on 14 February 2023).
- Sorg, J.A.; Sonenshein, A.L. Inhibiting the initiation of Clostridium difficile spore germination using analogs of chenodeoxycholic acid, a bile acid. J. Bacteriol. 2010, 192, 4983–4990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuijper, E.J.; Barbut, F.; Brazier, J.S.; Kleinkauf, N.; Eckmanns, T.; Lambert, M.L.; Drudy, D.; Fitzpatrick, F.; Wiuff, C.; Brown, D.J.; et al. Update of Clostridium difficile Infection due to PCR Ribotype 027 in Europe, 2008. Eurosurveillance 2008, 13, 18942. [Google Scholar] [CrossRef] [PubMed]
- NICE Guideline. Clostridioides difficile Infection: Antimicrobial Prescribing. 2021. Available online: https://www.nice.org.uk/guidance/ng199/resources/clostridioides-difficile-infection-antimicrobial-prescribing-pdf-66142090546117 (accessed on 21 February 2023).
- Chilton, C.; Pickering, D.; Freeman, J. Microbiologic factors affecting Clostridium difficile recurrence. Clin. Microbiol. Infect. 2017, 24, 476–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Calle, D.; Guimarães Benevides, R.; Góes-Neto, A.; Billington, C. Bacteriophages as Alternatives to Antibiotics in Clinical Care. Antibiotics 2019, 8, 138. [Google Scholar] [CrossRef] [Green Version]
- Murray, E.; Draper, L.A.; Ross, R.P.; Hill, C. The Advantages and Challenges of Using Endolysins in a Clinical Setting. Viruses 2021, 13, 680. [Google Scholar] [CrossRef]
- Mayer, M.J.; Narbad, A.; Gasson, M.J. Molecular Characterization of a Clostridium difficile Bacteriophage and Its Cloned Biologically Active Endolysin. J. Bacteriol. 2008, 190, 6734–6740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, M.J.; Garefalaki, V.; Spoerl, R.; Narbad, A.; Meijers, R. Structure-Based Modification of a Clostridium difficile-Targeting Endolysin Affects Activity and Host Range. J. Bacteriol. 2011, 193, 5477–5486. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Euler, C.W.; Delaune, A.; Fischetti, V.A. Using a Novel Lysin to Help Control Clostridium difficile Infections. Antimicrob. Agents Chemother. 2015, 59, 7447–7457. [Google Scholar] [CrossRef] [Green Version]
- Peng, Z.; Wang, S.; Gide, M.; Zhu, D.; Patabendige, H.M.L.W.; Li, C.; Cai, J.; Sun, X. A novel bacteriophage lysin-human defensin fusion protein is effective in treatment of clostridioides difficile infection in mice. Front. Microbiol. 2019, 9, 3234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ecelebi, O.; Ebuyuk, F.; Pottage, T.; Ecrook, A.; Ehawkey, S.; Ecooper, C.; Bennett, A.; Esahin, M.; Ebaillie, L. The Use of Germinants to Potentiate the Sensitivity of Bacillus anthracis Spores to Peracetic Acid. Front. Microbiol. 2016, 7, 18. [Google Scholar] [CrossRef]
- Nerandzic, M.M.; Donskey, C.J. A Quaternary Ammonium Disinfectant Containing Germinants Reduces Clostridium difficile Spores on Surfaces by Inducing Susceptibility to Environmental Stressors. Open Forum Infect. Dis. 2016, 3, ofw196. [Google Scholar] [CrossRef] [PubMed]
- Nerandzic, M.M.; Donskey, C.J. Sensitizing Clostridium difficile Spores with Germinants on Skin and Environmental Surfaces Represents a New Strategy for Reducing Spores via Ambient Mechanisms. Pathog. Immun. 2017, 2, 404–421. [Google Scholar] [CrossRef] [Green Version]
- Horgan, M.; O’Sullivan, O.; Coffey, A.; Fitzgerald, G.F.; van Sinderen, D.; McAuliffe, O.; Ross, R.P. Genome analysis of the Clostridium difficile phage φCD6356, a temperate phage of the Siphoviridae family. Gene 2010, 462, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Alyousef, A. Identification and characterisation of lysin enzymes as potential therapeutics for the treatment of Clostridium difficile. Ph.D. Thesis, Cardiff University, Cardiff, UK, 2013. [Google Scholar]
- Budi, N.; Safdar, N.; Rose, W.E. Treatment issues in recurrent Clostridioides difficile infections and the possible role of germinants. FEMS Microbes 2020, 1, 1–8. [Google Scholar] [CrossRef]
- Public Health England. Clostridium Difficile Ribotyping Network (CDRN) for England and Northern Ireland. 2016. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/491253/CDRN_2013-15_Report.pdf (accessed on 10 January 2023).
- Herbert, R.; Hatcher, J.; Jauneikaite, E.; Gharbi, M.; D’arc, S.; Obaray, N.; Rickards, T.; Rebec, M.; Blandy, O.; Hope, R.; et al. Two-year analysis of Clostridium difficile ribotypes associated with increased severity. J. Hosp. Infect. 2019, 103, 388–394. [Google Scholar] [CrossRef] [Green Version]
- Sebaihia, M.; Wren, B.W.; Mullany, P.; Fairweather, N.F.; Minton, N.; Stabler, R.; Thomson, N.R.; Roberts, A.P.; Cerdeño-Tárraga, A.M.; Wang, H.; et al. The multidrug-resistant human pathogen Clostridium difficile has a highly mobile, mosaic genome. Nat. Genet. 2006, 38, 779–786. [Google Scholar] [CrossRef]
- Gonzales-Luna, A.J.; Carlson, T.J.; Dotson, K.M.; Poblete, K.; Costa, G.; Miranda, J.; Lancaster, C.; Walk, S.T.; Tupy, S.; Begum, K.; et al. PCR ribotypes of Clostridioides difficile across Texas from 2011 to 2018 including emergence of ribotype 255. Emerg. Microbes Infect. 2020, 9, 341–347. [Google Scholar] [CrossRef] [Green Version]
- Kociolek, L.K.; Gerding, D.N.; Hecht, D.W.; Ozer, E. Comparative genomics analysis of Clostridium difficile epidemic strain DH/NAP11/106. Microbes Infect. 2018, 20, 245–253. [Google Scholar] [CrossRef]
- Schmelcher, M.; Waldherr, F.; Loessner, M.J. Listeria bacteriophage peptidoglycan hydrolases feature high thermoresistance and reveal increased activity after divalent metal cation substitution. Appl. Microbiol. Biotechnol. 2011, 93, 633–643. [Google Scholar] [CrossRef] [Green Version]
- Alabdali, Y.A.J.; Oatley, P.; Kirk, J.A.; Fagan, R.P. A cortex-specific penicillin-binding protein contributes to heat resistance in Clostridioides difficile spores. Anaerobe 2021, 70, 102379. [Google Scholar] [CrossRef]
- Stabler, R.A.; He, M.; Dawson, L.; Martin, M.; Valiente, E.; Corton, C.; Lawley, T.D.; Sebaihia, M.; Quail, M.A.; Rose, G.; et al. Comparative genome and phenotypic analysis of Clostridium difficile 027 strains provides insight into the evolution of a hypervirulent bacterium. Genome Biol. 2009, 10, R102. [Google Scholar] [CrossRef] [Green Version]
- Pizarro-Guajardo, M.; Calderón-Romero, P.; Romero-Rodríguez, A.; Paredes-Sabja, D. Characterization of Exosporium Layer Variability of Clostridioides difficile Spores in the Epidemically Relevant Strain R20291. Front. Microbiol. 2020, 11, 1345. [Google Scholar] [CrossRef]
- Buckley, A.M.; Spencer, J.; Candlish, D.; Irvine, J.J.; Douce, G.R. Infection of hamsters with the UK Clostridium difficile ribotype 027 outbreak strain R20291. J. Med Microbiol. 2011, 60, 1174–1180. [Google Scholar] [CrossRef] [Green Version]
- Drudy, D.; Goorhuis, B.; Bakker, D.; Kyne, L.; van den Berg, R.; Fenelon, L.; Fanning, S.; Kuijper, E.J. Clindamycin Resistant Clone of Clostridium difficile PCR Ribotype 027, Europe. Emerg. Infect. Dis. 2008, 14, 1485–1487. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.I.; Akter, A.; Draper, L.A.; Ross, R.P.; Hill, C. Characterization of an Endolysin Targeting Clostridioides difficile That Affects Spore Outgrowth. Int. J. Mol. Sci. 2021, 22, 5690. [Google Scholar] [CrossRef] [PubMed]
- Setlow, P.; Wang, S.; Li, Y.-Q. Germination of Spores of the Orders Bacillales and Clostridiales. Annu. Rev. Microbiol. 2017, 71, 459–477. [Google Scholar] [CrossRef] [PubMed]
- Kochan, T.J.; Somers, M.J.; Kaiser, A.M.; Shoshiev, M.S.; Hagan, A.K.; Hastie, J.L.; Giordano, N.P.; Smith, A.D.; Schubert, A.M.; Jr, P.E.C.; et al. Intestinal calcium and bile salts facilitate germination of Clostridium difficile spores. PLoS Pathog. 2017, 13, e1006443. [Google Scholar] [CrossRef] [Green Version]
- Mehta, K.K.; Paskaleva, E.E.; Azizi-Ghannad, S.; Ley, D.J.; Page, M.A.; Dordick, J.S.; Kane, R.S. Characterization of AmiBA2446, a Novel Bacteriolytic Enzyme Active against Bacillus Species. Appl. Environ. Microbiol. 2013, 79, 5899–5906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlon, J.M.; Mitchell, S.J.; Bailey-Kellogg, C.; Griswold, K.E. Bioinformatics-driven discovery of novel Clostridioides difficile lysins and experimental comparison with highly active benchmarks. Biotechnol. Bioeng. 2021, 118, 2482–2492. [Google Scholar] [CrossRef]
- Mauro, V.P. Codon Optimization in the Production of Recombinant Biotherapeutics: Potential Risks and Considerations. Biodrugs 2018, 32, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Begley, M.; Gahan, C.G.; Hill, C. The interaction between bacteria and bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, R.; Sahu, G.K.; Das, J. Stress response in pathogenic bacteria. J. Biosci. 1996, 21, 149–160. [Google Scholar] [CrossRef]
- Oberkampf, M.; Hamiot, A.; Altamirano-Silva, P.; Bellés-Sancho, P.; Tremblay, Y.D.N.; DiBenedetto, N.; Seifert, R.; Soutourina, O.; Bry, L.; Dupuy, B.; et al. c-di-AMP signaling is required for bile salt resistance, osmotolerance, and long-term host colonization by Clostridioides difficile. Sci. Signal. 2022, 15, eabn8171. Available online: https://www.science.org (accessed on 5 February 2023). [CrossRef] [PubMed]
- Van Pijkeren, J.-P.; Barrangou, R. Genome Editing of Food-Grade Lactobacilli to Develop Therapeutic Probiotics. Microbiol. Spectr. 2017, 5, 389–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Ribotypes | Clinical Relevance | References |
|---|---|---|---|
| NCTC 12727 | 001 | Increased resistance to vancomycin | [21] |
| R31760 | 002 | The most common ribotype in England | [22] |
| R31762 | 005 | Third most common ribotype in England | [22] |
| DS1684 | 010 | ||
| CD630 | 012 | Came from an outbreak in Zurich, Switzerland; first strain to be genome sequence | [23] |
| R31755 | 014 | Fourth most common ribotype in England | [22] |
| R31774 | 020 | Fifth most prevalent ribotype in England | [22] |
| DS1665 | 023 | The 10th most common ribotype in England | [22] |
| DS1813/ R20291 | 027 | Hypervirulent, most common ribotype in Texas, USA. R20291 isolated in 2006 from an outbreak in Stoke Mandeville Hospital, England | [24] |
| R30776 | 045 | ||
| R31263 | 046 | ||
| R25961 | 047 | ||
| R31312 | 056 | ||
| R31777 | 078 | Sixth most common ribotype in England | [22] |
| DS1787 | 106 | The most common ribotype in the USA | [25] |
| R30967 | 110 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alyahya, K.; Baillie, L. Assessing the Feasibility of Employing a Combination of a Bacteriophage-Derived Endolysin and Spore Germinants to Treat Relapsing Clostridioides difficile Infection. Microorganisms 2023, 11, 1651. https://doi.org/10.3390/microorganisms11071651
Alyahya K, Baillie L. Assessing the Feasibility of Employing a Combination of a Bacteriophage-Derived Endolysin and Spore Germinants to Treat Relapsing Clostridioides difficile Infection. Microorganisms. 2023; 11(7):1651. https://doi.org/10.3390/microorganisms11071651
Chicago/Turabian StyleAlyahya, Khalid, and Les Baillie. 2023. "Assessing the Feasibility of Employing a Combination of a Bacteriophage-Derived Endolysin and Spore Germinants to Treat Relapsing Clostridioides difficile Infection" Microorganisms 11, no. 7: 1651. https://doi.org/10.3390/microorganisms11071651