1. Introduction
Microalgae are photosynthetic eukaryotic organisms inhabiting mostly aquatic environments, both freshwater and saltwater. One of their main features is the fixation of carbon dioxide during the process of photosynthesis while producing oxygen [
1,
2,
3]. Using carbon dioxide as a main source of energy is most common, however, some mixotrophs are capable of the occasional usage of organic carbon sources to produce energy [
1,
4]. Microalgae are present worldwide and they are able to adapt to more difficult environments [
5,
6]. They are able to survive in extreme conditions such as high temperatures, high salinity, the presence of high concentrations of metals and even combinations of these conditions, and more [
5]. Microalgal biomass is rich in valuable bioactive metabolites such as proteins, carbohydrates, lipids, polysaccharides, pigments, etc. As a rich source of fiber, vitamins and minerals, microalgae have found wide application in the food industry, especially as nutritional supplements and animal feed additives [
7,
8,
9]. The production of biologically active substances makes them great for potential use in the pharmaceutical industry [
9,
10,
11]. As the issue with fossil fuels as energy resources grows more important, the research has focused on using algae as a biofuel source, mostly the production of biodiesel from microalgae [
3,
12,
13,
14].
With a growing population, the demand for food production increases. This trend has seen the amount of food waste increase yearly [
15] (estimated at 1.3 billion tons annually) [
16,
17]. Food waste is either disposed into landfills or can be treated otherwise. One of the options is using food waste as feedstock in biotechnology processes [
17]. Food waste is rich in organic compounds such as carbohydrates, fats, nitrogenous compounds, vitamins, minerals, etc., which can be used as a source of organic carbon for microorganisms [
16].
The coffee industry is growing each year due to the enormous popularity of the beverage and coffee products. During the process of brewing coffee or making instant coffee, a large amount of waste in the form of spent coffee grounds (SCG) is generated [
18,
19,
20]. Spent coffee grounds are still rich in organic components such as polysaccharides, lignin and proteins, oils, and many smaller components that can include phenolic compounds, pigments, etc. [
21]. Depending on the type of coffee source, SCG can contain from 10 up to 20% of oil. One possible application of coffee oil is its conversion into biodiesel through the transesterification method [
22].
The food industry also produces a large number of other oil-based waste products among which waste frying oil (WFO) is important, mainly coming from fast-food chains and large restaurants [
23]. Not only is there a large amount of waste frying oil, but its disposal also poses an environmental problem as it is regularly poured into drains [
24,
25]. WFOs are highly available and relatively cheap, therefore they have been researched for biodiesel synthesis [
26,
27]. However, due to the chemical changes during the frying process, the quality of the biodiesel can be significantly influenced. This requires some pre-treatment of the feedstock and detailed control of the process [
25,
28].
The aim of this study was to test the ability of selected algal strains to grow mixotrophically in a basic Bold’s Basal Medium (BBM) with added food waste oils as an organic carbon source. This experimental work was conducted as a pilot study for subsequent research, mainly for yeast:microalgae co-cultivation experiments [
29]. To the best of our knowledge, the present study is the first work focused on the cultivation of microalgae on oil-based food waste materials. Coffee oil was used in this study for a comparison with waste frying oil and also for the possibility of testing microalgae for the absorption of valuable substances from these oils. In our previous experiments with yeast, the high capacity of yeast to absorb valuable substances from wastes such as tocopherol from coffee oil was confirmed.
3. Results
In this study, we present the first published results that examined the possible growth of microalgae on oil waste substrates. Based on our previous experiments [
29], cultivations were conducted to determine whether the microalgae cultures were able to grow in simple BBM with the addition of different amounts of waste frying oil and coffee oil. The main goal was to determine whether the studied strains were able to utilize waste lipid substrates, and thus to incorporate some molecules from used oils to the biomass. We aimed to verify whether mixotrophy growth could be used in co-cultivation experiments of microalgae with heterotrophic microorganisms such as red yeasts to produce a combined yeast:microalgae enriched biomass that could be used as a food and feed additive [
7]. In the experiments, the growth rate of microalgae in the control media was monitored and compared to the media with added oils. In the case where the microalgae culture was not able to utilize the added oils, it was necessary to monitor whether the presence of the oil had a negative effect on the viability of the microalgae, or whether it inhibited photosynthesis by obstructing the light availability and thus the absorption of light by cells. Different concentrations of the tested oils were then used to identify the limits of tolerance of the microalgae to the oil, as this parameter is very important for subsequent co-cultivation experiments.
3.1. Desmodesmus Armatus CCALA 439
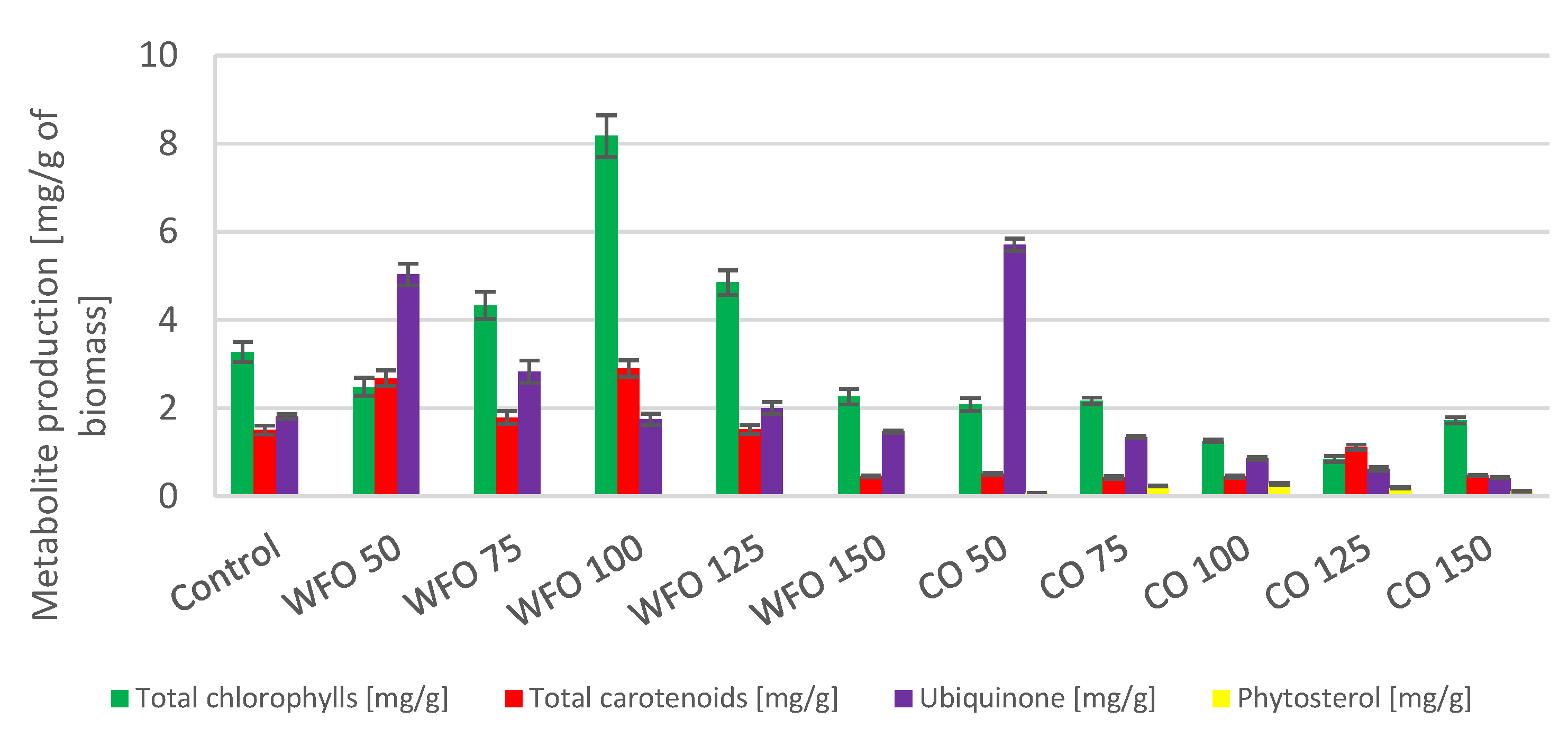
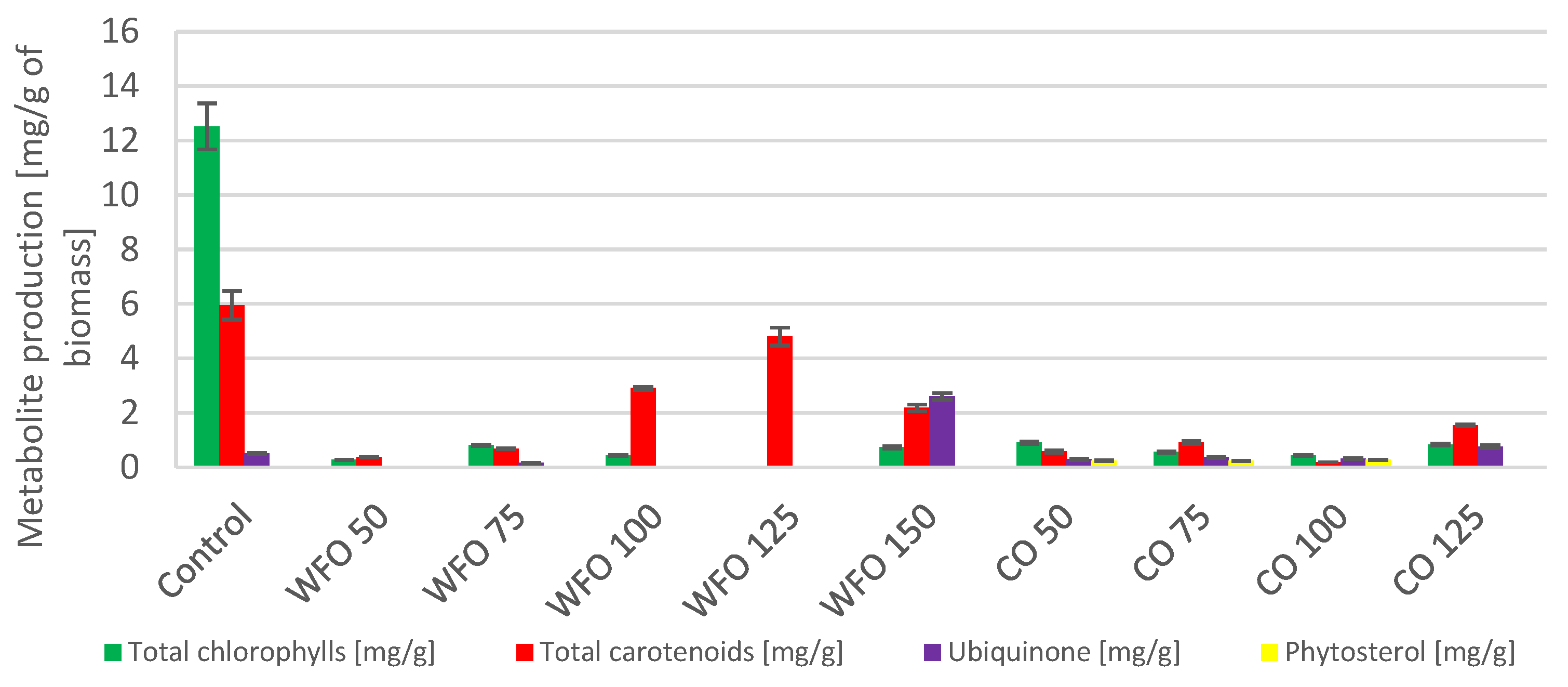
Cultivation of
Desmodesmus armatus with the addition of waste frying oil showed a higher production of total chlorophylls compared to the control except for WFO 50 and WFO 150. The highest production of total chlorophylls was obtained with WFO 100 (8.171 ± 0.475 mg/g). The production of total carotenoids did not seem to depend on the concentration of oil present in the medium. Apart from the highest concentration, WFO 150, the production of carotenoids was higher compared to the control. The best production of carotenoid pigments (
Figure 1) was found in sample WFO 100 (2.901 ± 0.188 mg/g). In the case of ubiquinone, the production in sample WFO 50 was greatly increased (5.031 ± 0.246 mg/g) compared to the control, and an observable trend of lowering its concentration with increasing concentrations of oil in the medium was present. Only in sample WFO 150 was a lower production observed than in the control. Overall, sample WFO 100 showed a higher production of targeted metabolites compared to the control. In sample WFO 150, the production decreased considerably compared to the control.
Regarding the samples cultivated with the addition of coffee oil, the production of metabolites was decreased compared to the control and samples cultivated with WFO. Except for ubiquinone, all metabolite production was lower than the control. The greatest production could be seen in sample CO 50 (5.708 ± 0.138 mg/g), which was also higher than in sample WFO 50. With higher concentrations of CO, the production of ubiquinone decreased more than in the WFO samples. Within the samples cultivated in the presence of coffee oil, the production of total carotenoids was significantly higher in the sample CO 125. The best production of total chlorophylls was in sample CO 75 (2.167 ± 0.075 mg/g). The samples cultivated on coffee oil showed a slight production of sterols, with the best production in CO 100 (0.284 ± 0.022 mg/g).
The cultivation in the presence of oil showed a positive effect on the production of biomass with the exception of sample CO 150, where the yield was lower than the control. The biomass yield in samples cultivated on WFO was slightly higher overall than in the samples cultivated on CO, with sample WFO 150 displaying the greatest production of biomass (1.134 ± 0.099 g/L) (
Table 3). The Shapiro–Wilk test for normality determination was applied to analyze the data for biomass production. This parameter did not show a normal distribution, therefore the Kruskal–Wallis method was applied (
p = 0.8804).
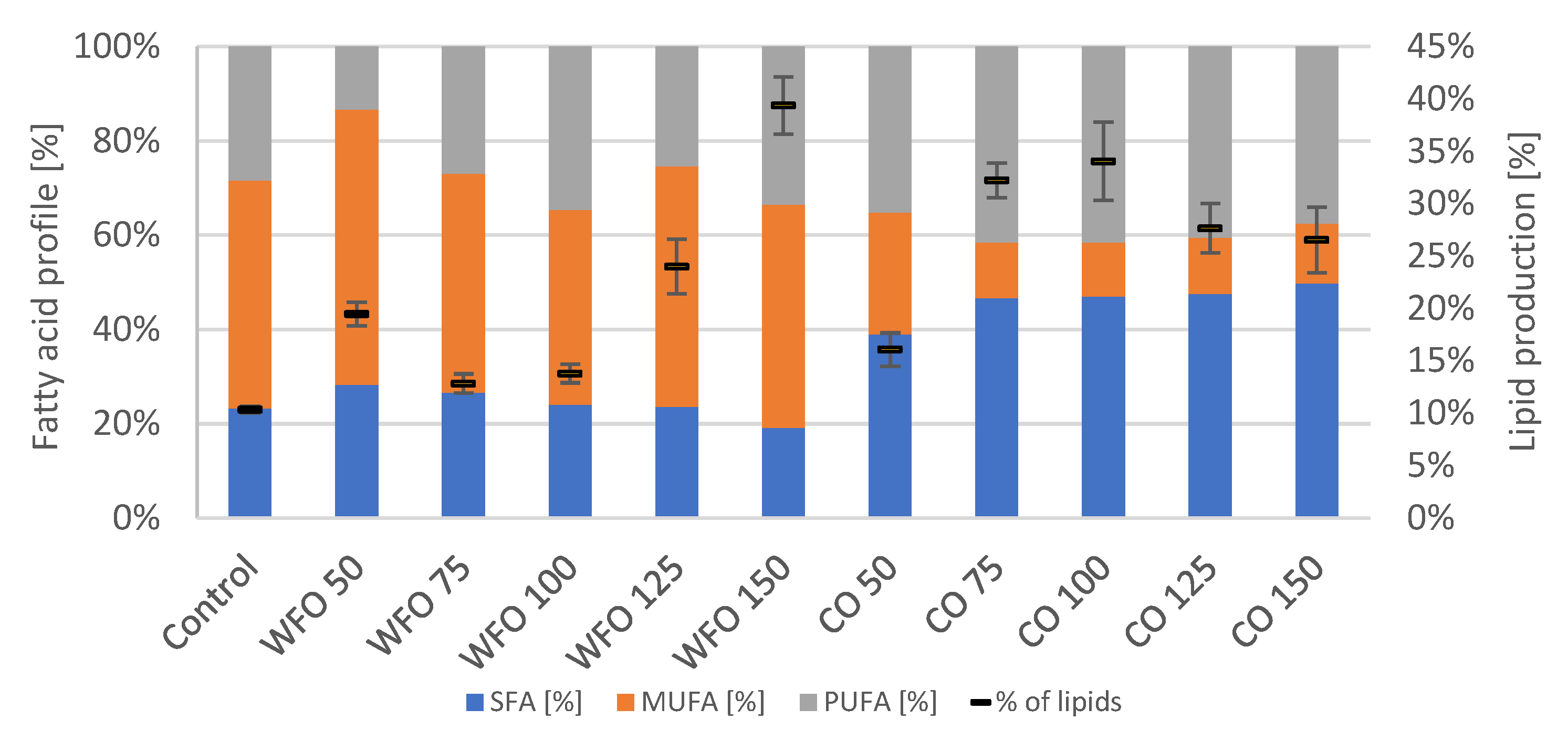
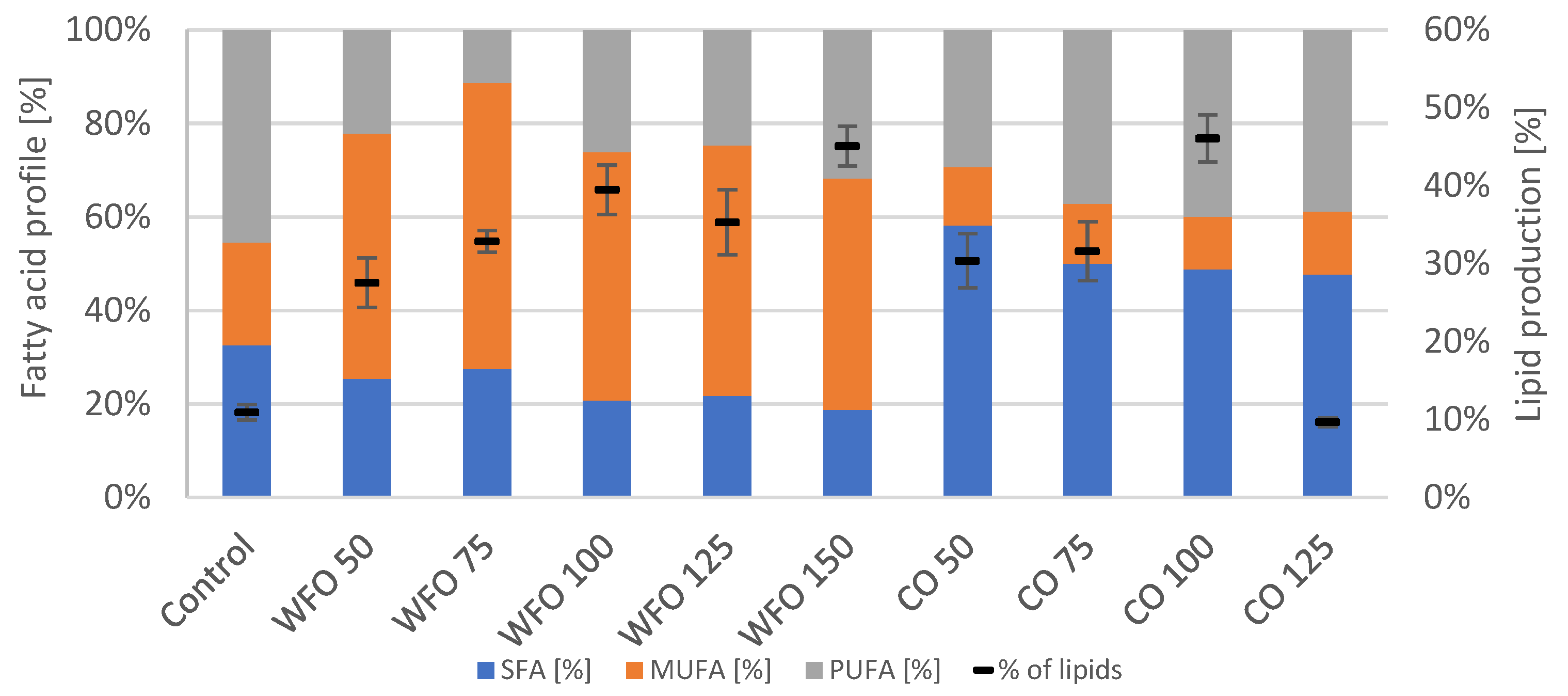
The GC analysis of lipid production showed (
Figure 2) that cultivation with the addition of oils did increase the production of total lipids compared to the control. For WFO, the lipid production increased with the higher concentration of oil with the exception of WFO 50. The highest lipid production was recorded in sample WFO 150 (39.38 ± 2.74%). The profile of fatty acids was not significantly different among the samples cultivated on WFO and the control, which had the highest representation of monounsaturated fatty acids (MUFA) (above 40%); the highest content was in sample WFO 50 (58.39%). The content of SFA ranged from about 19% to 28%. The least amount of polyunsaturated fatty acids (PUFA) was in sample WFO 50 (13.36%).
The samples cultivated with coffee oil also showed higher lipid production than the control with the lowest production in sample CO 50, then increased with higher concentrations ranging from 16 to 34%. The fatty acid composition in samples cultivated with coffee oil did not differ significantly among each other with the slight exception of sample CO 50. The samples cultivated on coffee oil showed the highest representation of saturated fatty acids (SFA) (above 40%) and the lowest for MUFA (approx. 11–13%).
The Shapiro–Wilk test for normality determination was also applied to analyze the data for lipid production. This parameter also did not show a normal distribution, therefore, the Kruskal–Wallis method was applied for lipid production (p = 0.2932).
3.2. Scenedesmus Acutus CCALA 904
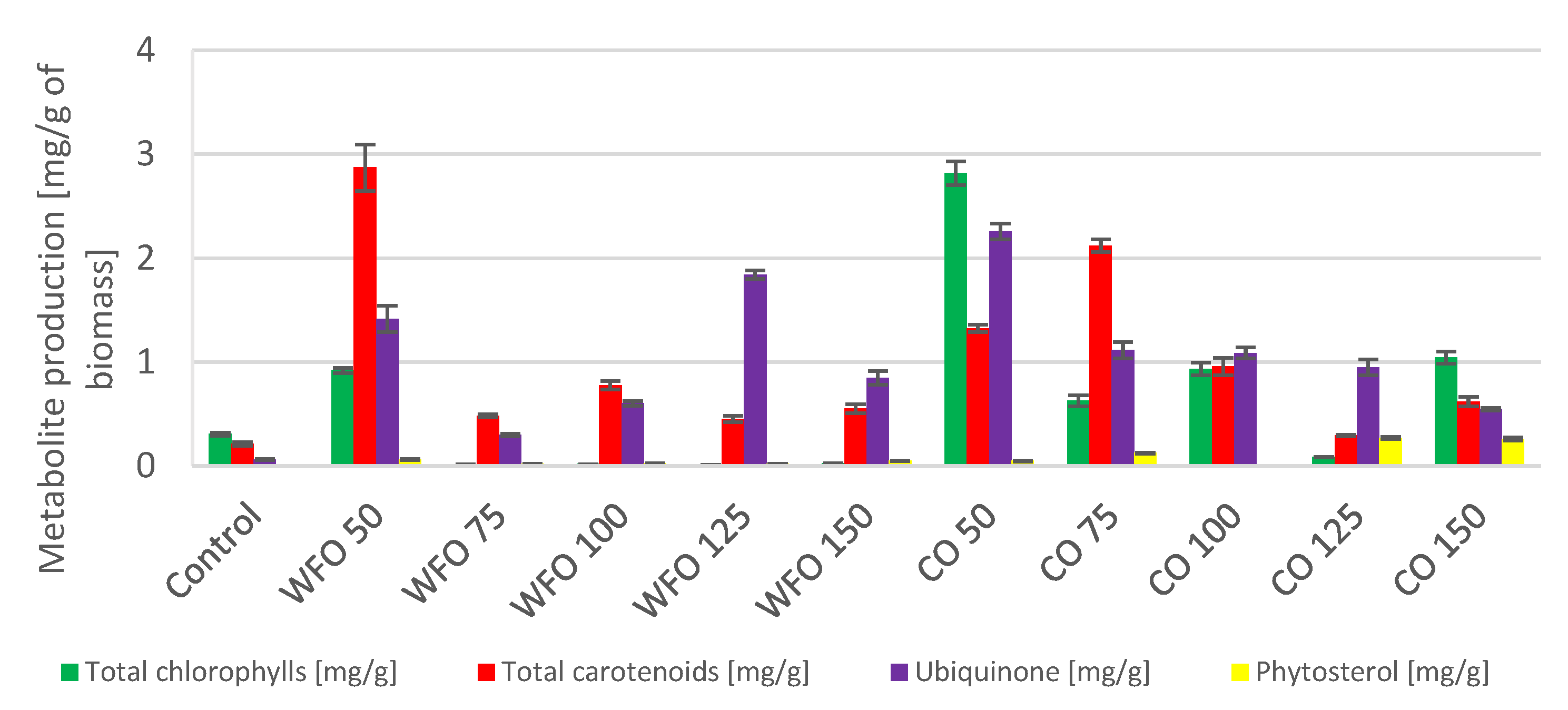
Microalgae cultivated on frying oil displayed a decrease in the production of total chlorophylls, except for WFO 50. This sample showed the greatest production of chlorophylls (0.920 ± 0.025 mg/g) as well as total carotenoids (2.872 ± 0.224 mg/g). The total carotenoid production decreased with higher concentrations of oil but these productions were higher than that of the control. A considerable increase (
Figure 3) was observed in the production of ubiquinone, where the best production was in sample WFO 125 (1.842 ± 0.040 mg/g). The production of certain metabolites was higher in samples cultivated with added oil compared to the control, however, the productions of chlorophylls and carotenoids were drastically decreased, possibly by the degradation of the mentioned metabolites.
In the case of coffee oil, the addition did not have such a negative effect on the production of total chlorophylls as waste frying oil. Therefore, the production of metabolites was also increased compared to the control. The production of chlorophylls was the highest in sample CO 50 (2.818 ± 0.112 mg/g), and the production in other samples was also rather high compared to the control, with the exception of sample CO 125. The production of total carotenoids was predominantly higher for the three lowest concentrations of coffee oil. The highest carotenoid production was seen in sample CO 75 (2.121 ± 0.062 mg/g). For ubiquinone, the best production was found in sample CO 50 (2.257 ± 0.076 mg/g) and decreased as concentrations of oil in the medium was increased. With the addition of coffee oil in the medium, the production of sterols was slightly increased, and the highest values were measured in sample CO 125 (0.270 ± 0.010 mg/g). The coffee oil did not have a negative effect on the production of chlorophylls and other carotenoids (pigments), while the frying oil showed a negative effect with the exception of culture sample WFO 50.
In the case of algae
Scenedesmus acutus, the addition of oil had a positive effect on the production of biomass for both WFO and CO. The greatest production was achieved by samples WFO 100 (1.190 ± 0.085 g/L) and CO 150 (1.214 ± 0.149 g/L) (
Table 4). Data from strain
Scenedesmus acutus were subjected to the Shapiro–Wilk test, which did not show a normal data distribution. The Kruskal–Wallis test was then applied for biomass production (
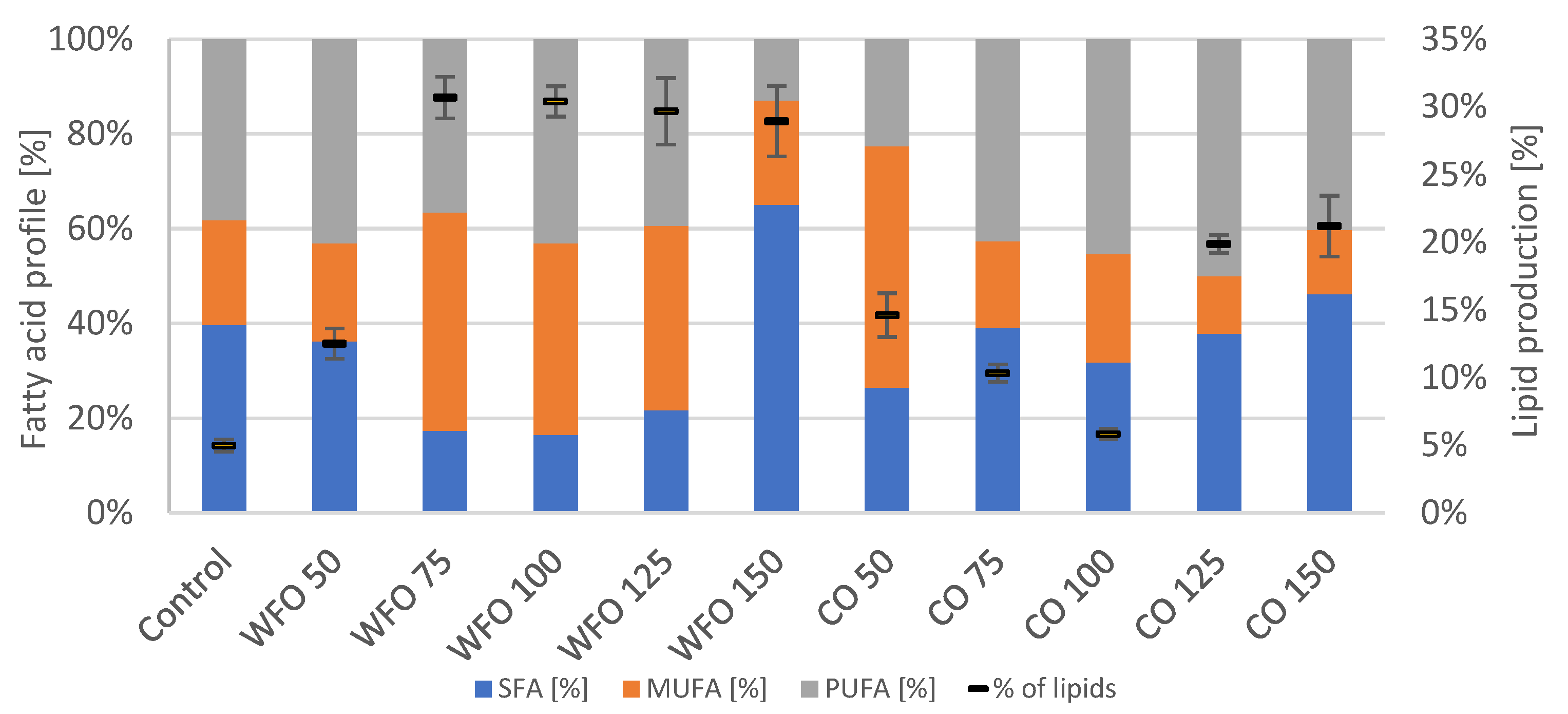
p = 0.2606). Analysis of the total amount of lipids in the biomass showed an increased production of lipids in all samples cultivated with oil compared to the control. The samples cultivated on waste frying oil showed production ranging from approx. 29% to 31%, with the exception of sample WFO 50 (12.5 ± 1.11%). The samples cultivated on coffee oil showed a decrease in the production of lipids with an increasing concentration of oil until sample CO 100, then increased rapidly in the case of the two highest concentrations of oil. The highest lipid content (
Figure 4) was found in sample CO 150 (21.19 ± 2.25%). In comparison with the waste frying oil, the samples cultivated on coffee oil did not reach that high amount of lipids. The data for lipid production were subjected to the Shapiro–Wilk test, which did not show a normal distribution of data. The Kruskal–Wallis test was then applied (
p = 0.4297).
The fatty acid composition did not seem to follow a united trend for all samples (
Figure 4). However, with the exception of CO 50, the samples cultivated on coffee oil produced the least amount of MUFA (approx. 12–23%), while the highest production varied between SFA and PUFA.
3.3. Chlamydomonas Reindhartii CCALA 973
Microalgae
Chlamydomonas reindhartii displayed a severe formation of emulsion (
Figure S1). A significant decrease in the production of total chlorophylls as well as other metabolites was observed in this strain (
Figure 5). Among the samples cultivated on waste frying oil, sample WFO 75 showed the greatest production of total chlorophylls (0.812 ± 0.034 mg/g). The production of carotenoids was higher in the samples also containing a higher amount of WFO in the medium, with the best production in sample WFO 125 (4.802 ± 0.327 mg/g). The only significant production of ubiquinone was seen in sample WFO 150 (2.607 ± 0.132 mg/g), which was also higher compared to the control. Samples cultivated on coffee oil showed similar results, especially a significant decrease in the production of metabolites (pigments). Among the samples, the highest production of total chlorophylls was in sample CO 50 (0.907 ± 0.053 mg/g). Sample CO 125 achieved the highest production of ubiquinone (0.764 ± 0.060 mg/g). Additionally, a slight production of sterols could be seen in the samples cultivated on coffee oil. The production of chlorophylls and carotenoids was severely decreased, possibly due to light obstruction during cultivation.
Microalgae samples cultivated on both WFO and CO displayed an increase in the biomass production compared to the control. The biomass production differed in individual samples and was not dependent on parameters of the cultivation. Sample WFO 75 displayed the greatest biomass yield (1.376 ± 0.096 g/L) (
Table 5) within the samples cultivated on WFO. Overall, the highest biomass production was found in sample CO 75 (1.496 ± 0.182 g/L). The biomass production data were subjected to the Shapiro–Wilk test, which determined a normal data distribution. Therefore, the ANOVA method was performed (
p = 0.000791).
Unlike the HPLC analysis, the GC analysis (
Figure 6) showed an increased production of lipids in all samples cultivated in the presence of oil with the exception of sample CO 125 (9.67 ± 0.54%), which was slightly lower compared to the control (10.94 ± 0.99%). The highest production of lipids could be seen in sample CO 100 (46.07 ± 3.03%) and WFO 150 (45.09 ± 2.54%). The production of lipids in samples cultivated on waste frying oil showed an increase at higher concentrations of oil in the medium. Regarding the representation of fatty acids in the lipidic part, the content of each group seemed to be similar among the samples cultivated on the same type of oil (
Figure 6). The samples cultivated on waste frying oil showed a very high production of MUFA (above 49%). The Shapiro–Wilk test for lipid production did not show a normal distribution of data. The Kruskal–Wallis test was performed (
p = 0.0212).
The highest level of MUFA was observed in sample WFO 75 (61.14%). The least represented group was either the SFA or PUFA. The lowest production of PUFA was also seen in sample WFO 75 (11.34%). Samples cultivated on coffee oil had the highest SFA (approx. 47–59%) and lowest MUFA production (up to 14%). This fatty acid profile was more similar to the control than the profile of the samples cultivated on waste frying oil.
3.4. Anabaena Torulosa CCALA 3
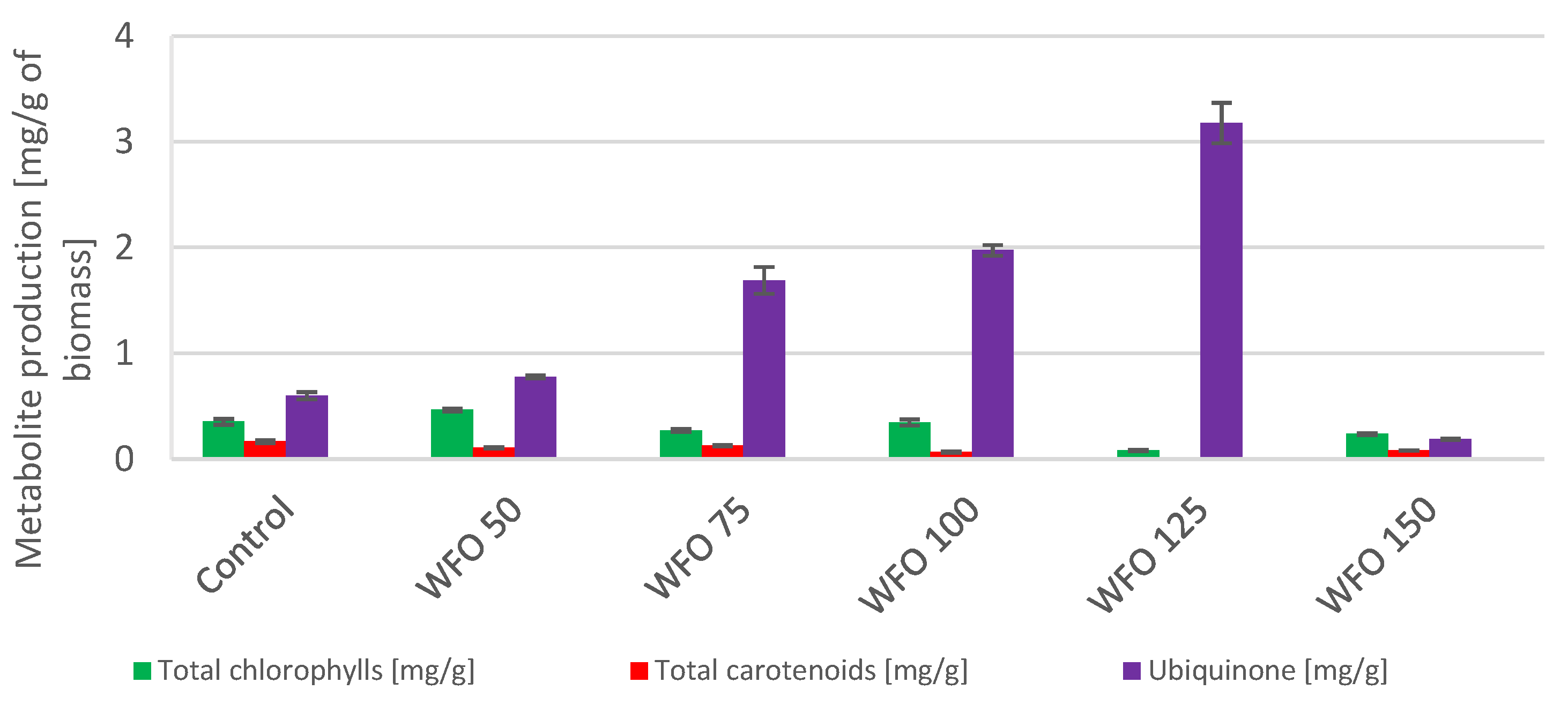
The presence of coffee oil in the media showed a negative effect on the viability of cultures of the strain
Anabaena torulosa and caused their death. Cyanobacteria samples cultivated on WFO displayed a decrease in the production of chlorophylls (
Figure 7) with the exception of sample WFO 50 (0.464 ± 0.015 mg/g), whose yield was higher compared to the control. Regarding the total carotenoids, the highest production among the samples cultivated in the presence of oil was found in sample WFO 75 (0.126 ± 0.006 mg/g), which was, however, lower compared to the control. The production of ubiquinone displayed a higher yield in samples cultivated on oil compared to the control and increased with higher concentrations of oil in the medium. Sample WFO 125 displayed the greatest ubiquinone production (3.177 ± 0.191 mg/g) with the exception of sample WFO 150 (0.187 ± 0.008 mg/g), where the production was decreased significantly.
Concerning this strain, the addition of oil did not have any significant effect on the production of biomass. Sample WFO 50 displayed the greatest biomass yield (0.520 ± 0.026 g/L) (
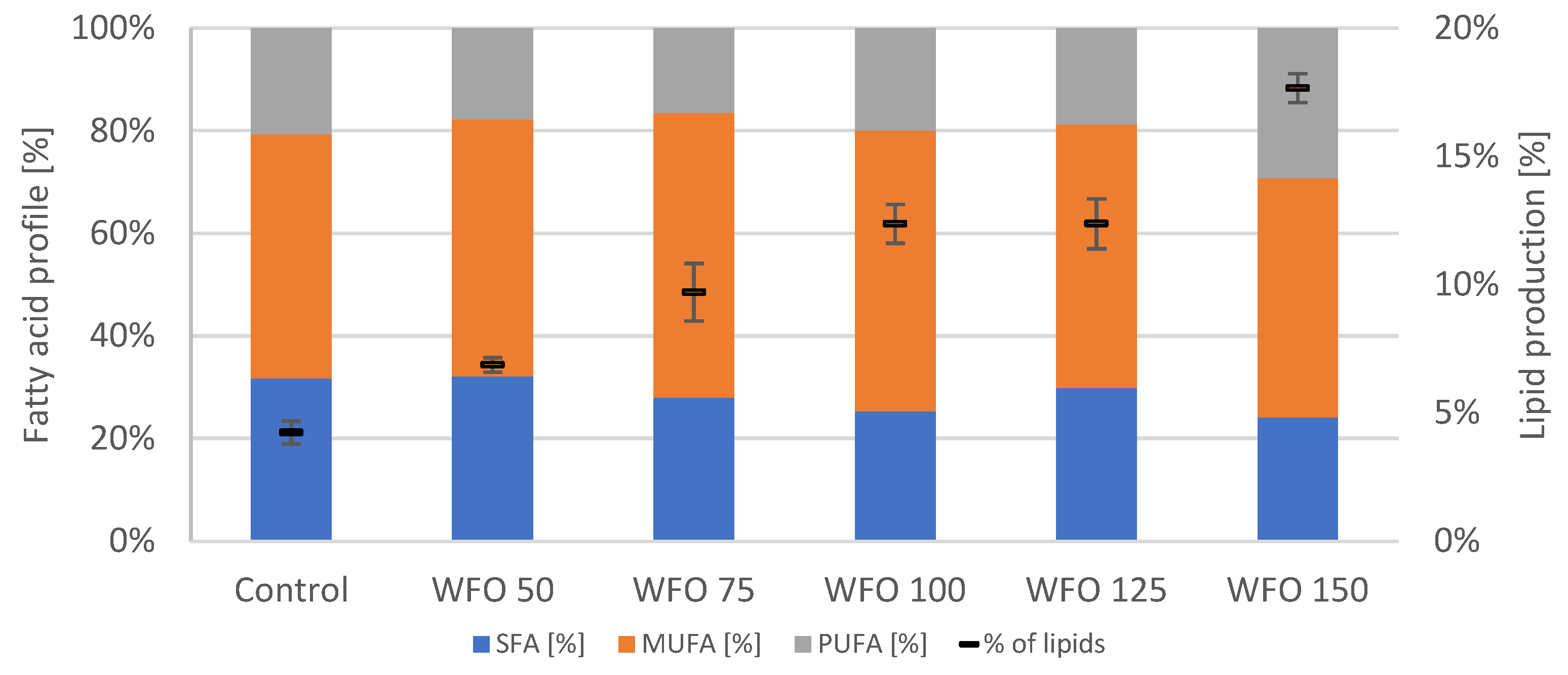
Table 6). The production of lipids (
Figure 3) grew with increasing concentrations of oil in the medium, with the best production in sample WFO 150 at 17.66 ± 0.57%. The profile of fatty acids did not significantly differ from the control (
Figure 8). The highest portion were MUFA ranged from approx. 47% to 56%, while the PUFA were represented the least, ranging from approx. 16% to 30%.
Triplicates were performed on this strain, however, throughout the tests, many of the cultures did not survive. Therefore, from the obtained data, we were not able to perform the ANOVA analysis. These tests will be repeated in the near future.
4. Discussion
In the present work, some microalgae were cultivated in the presence of different concentrations of either waste frying oil or coffee oil. The main focus of this experiment was to determine the viability and growth of the cultures in the presence of an oil-based nutrition source in the media, as this experiment was conducted as a pilot study for further experiments. Furthermore, the effect of the oil on the production of some specific algae metabolites was analyzed. The presence of oil in the medium caused the creation of emulsion with different severity. With the more severe creation of an emulsion in the medium, the obstruction of light absorption was also more severe. As a consequence, a negative effect on the production of pigments was observed, and simultaneously, an increase in the production of lipids was observed. Additionally, the darker color of coffee oil could have negative effect on the light transmission into the medium.
Cultivation of the microalgae Desmodesmus armatus CCALA 439 resulted in a decreased production of total chlorophylls and carotenoids in media with added coffee oil. The production of ubiquinone was the highest for the samples cultivated in the presence of the lowest concentrations of both waste frying oil and coffee oil (WFO 50, CO 50), with the greatest production in sample CO 50 (5.708 ± 0.138 mg/g). A significant increase in the production of total chlorophylls was observed in sample WFO 100 (8.717 ± 0.475 mg/g). Regarding the production of total carotenoids concerned, a noticeable increase was observed in samples WFO 50 and WFO 100, with the greatest production in sample WFO 100 (2.901 ± 0.788 mg/g). For the samples cultivated in the presence of coffee oil in the media, the production of the metabolites was mostly decreased compared to the control. As above-mentioned, with the decreased production of pigments in the samples cultivated on coffee oil, the production of lipids did increase. A noticeable increase in the production of lipids in the samples cultivated on WFO was also observed to have the greatest production of lipids, as shown in sample WFO 150 (39.38 ± 2.74%). The explanation is probably complex and except for the influence on photosynthesis includes at least (i) the oxidation of nutrients to acetyl-CoA as a precursor of both isoprenoids and lipids and (ii) the incorporation of lipids from media into the cell biomass.
The microalgae
Scenedesmus acutus CCALA 904 showed a production of lower amounts of total chlorophylls and carotenoids in samples cultivated on waste frying oil compared to the samples cultivated on coffee oil. The greatest production of total carotenoids could be seen in sample WFO 50 (2.872 ± 0.224 mg/g). For sample CO 50, the highest production of total chlorophylls (2.818 ± 0.112 mg/g) and ubiquinone (2.257 ± 0.076 mg/g) could be observed. Within the samples cultivated on coffee oil, the production of phytosterols was noticed, with the highest production in sample CO 125 (0.270 ± 0.010 mg/g). The production of metabolites was the opposite to the previous algal strain, which is probably a result of a different formation of an emulsion within the samples. The production of lipids followed this trend, where the production of lipids was higher in samples with a lower production of pigments, in this case, the samples cultivated on waste frying oil. This result is similar to the previous example and is also similar to the metabolic changes of some carotenogenic yeasts grown on fat media [
30]. In
S. acutus, the greatest production was observed in sample WFO 75, with a lipid yield above 30%.
In samples of microalgae Chlamydomonas reindhartii CCALA 973, the creation of an emulsion was very strong, regardless of the used oil, resulting in a significant decrease in the production of pigments. The greatest production of total carotenoids was observed in sample WFO 125 (4.802 ± 0.327 mg/g). For the metabolite ubiquinone, the greatest production could be seen in the sample with the highest concentration of waste frying oil, WFO 150 (2.607 ± 0.132 mg/g). Among the samples cultivated with oil in the media, the greatest production was observed in sample CO 50 (0.907 ± 0.053 mg/g), which was, however, several times lower than the production of total chlorophylls in the control sample. Samples seemed to be showing an increase in the production of lipids with increasing concentrations of oil in the media. The greatest productions of lipids were observed in samples WFO 150 (45.09 ± 2.54%) and CO 100 (46.07 ± 3.03%).
In the case of cyanobacteria Anabaena torulosa, the samples cultivated with coffee oil were not viable, regardless of the concentration of coffee oil used in the media. A significant increase in the production of ubiquinone was observed with increasing concentrations of waste frying oil, with the greatest production in sample WFO 125 (3.177 ± 0.191 mg/g). The production of ubiquinone could be considered as an adaptation mechanism supporting cell energy metabolism. The greatest production of total chlorophylls was observed in sample WFO 50 (0.464 ± 0.015 mg/g), which was also higher than the production of the control. The production of chlorophylls in other samples was decreased. The production of total carotenoids was also decreased compared to the control, with the greatest production in sample WFO 75 (0.126 ± 0.006 mg/g). The production of lipids increased with increasing concentrations of oil in the media.
Nowadays, lipid production by microorganisms is a rather attractive area of research and is mostly focused on microbial lipid production from organic waste and its further use (e.g., for biodiesel). Microalgae, as oleaginous microorganisms, have also been cultivated on carbon waste to produce a higher lipid content [
32]. In [
33], volatile fatty acids (VFAs) from low-cost organic waste were used as a majority carbon source for the cultivation of
Chlorella vulgaris, however, the lipid production was not as high as desired. VFAs were also used as a carbon source for marine oleaginous microorganisms [
34], where two freshwater algae,
A. protothecoides and
C. sorokiniana, were tested for their lipid production, and 28.97 and 33.79% of their cellular compartment lipids could be used as biodiesel feedstock, respectively. In another study [
35], the selected carbon source was washing water rich in glycerol, where an increase of 25% in microalgae production was obtained from the cultivation of
C. vulgaris. Other problems were also addressed such as the low oil content in microalgae cells and the higher costs of microalgae cultivation. The presentation of in situ technologies eliminates the need to extract the oil from biomass, therefore reducing the biodiesel production costs [
36]. Although further research is required, other microorganisms have also been studied for their possibility to degrade and utilize waste. One of the researched fields is petroleum degradation by microorganisms (bacteria, fungus, …). Their ability to adapt to the environment and use the waste substrate as a carbon source is a very exciting opportunity for future research [
37,
38].
The microorganisms studied in this work did not show a united trend in the production of various metabolites. The production did not seem to be dependent on the used waste oil type or on the concentration of the oil in the media. The main influence on the production of metabolites and the metabolic activity as a whole seemed to be the formation of the emulsion and its severity. These effects should be further studied because there is a lack of literary data with which the results of this study can be compared.
Experiments introduced in the present study confirmed the ability of microalgae cultures to withstand the presence of either waste frying oil or coffee oil in the medium. The original metabolic strategy originated in our previous co-cultivation experiments of red yeasts and microalgae. A number of surprising, interesting, and new data regarding the production of intracellular metabolites were gained from the results. To the best of our knowledge, this study is the first to have focused on the use of oil-based waste substrates in the cultivation of microalgae. These results will provide the necessary information for further experiments and possibly new approaches in the cultivation of microalgae using waste food lipid substrates as a carbon source.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}