
Probiotics, Prebiotics, and Synbiotics Utilization in Crayfish Aquaculture and Factors Affecting Gut Microbiota

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Pro-, Pre-, and Synbiotics
3. Main Pre-, Pro-, and Synbiotics Substances Administered in Crayfish
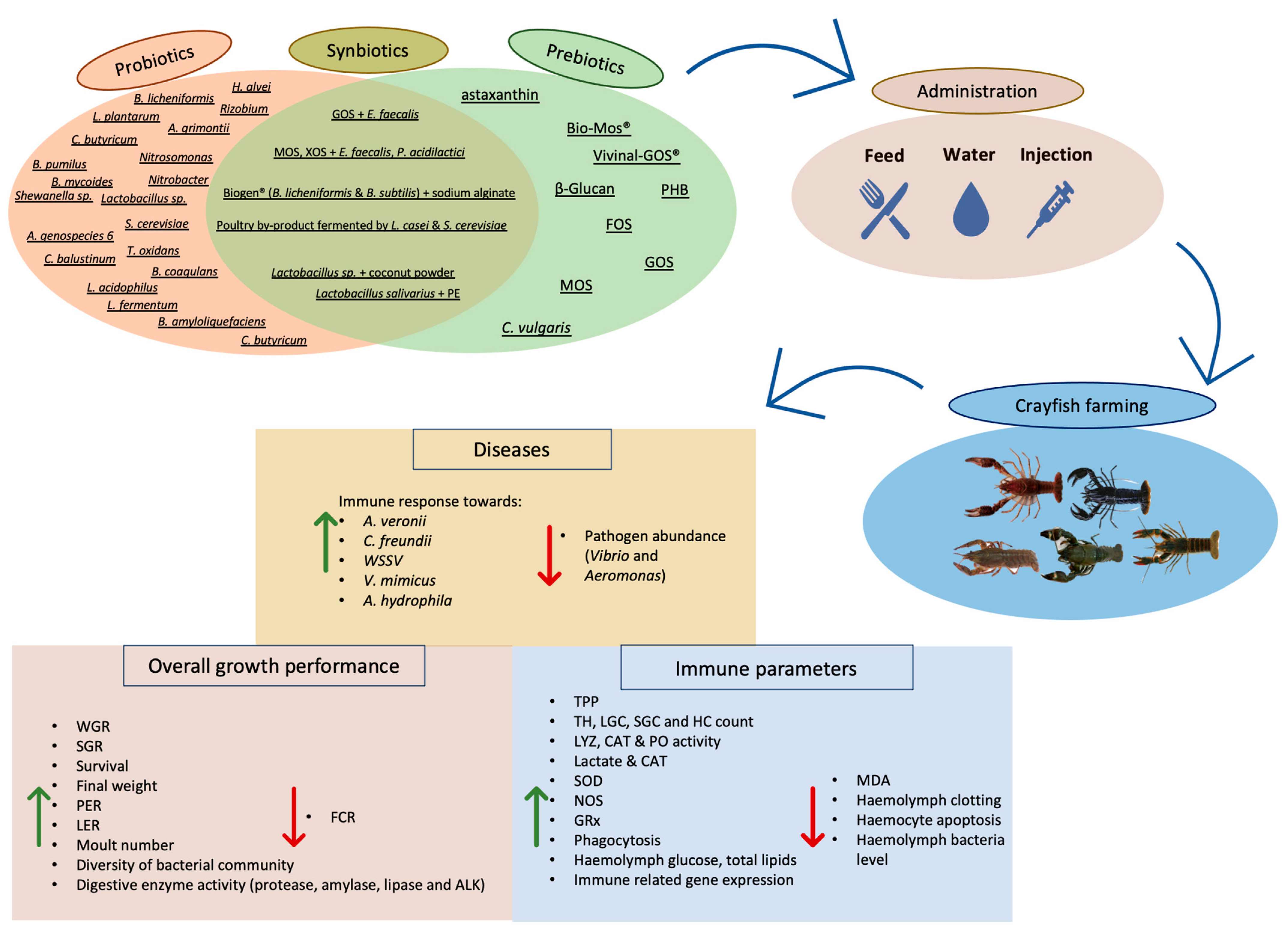
3.1. Probiotics Administration
3.2. Prebiotics Administration
3.3. Synbiotics Administration
3.4. Synopsis of Pro-, Pre-, and Synbiotics Administration and Limitations
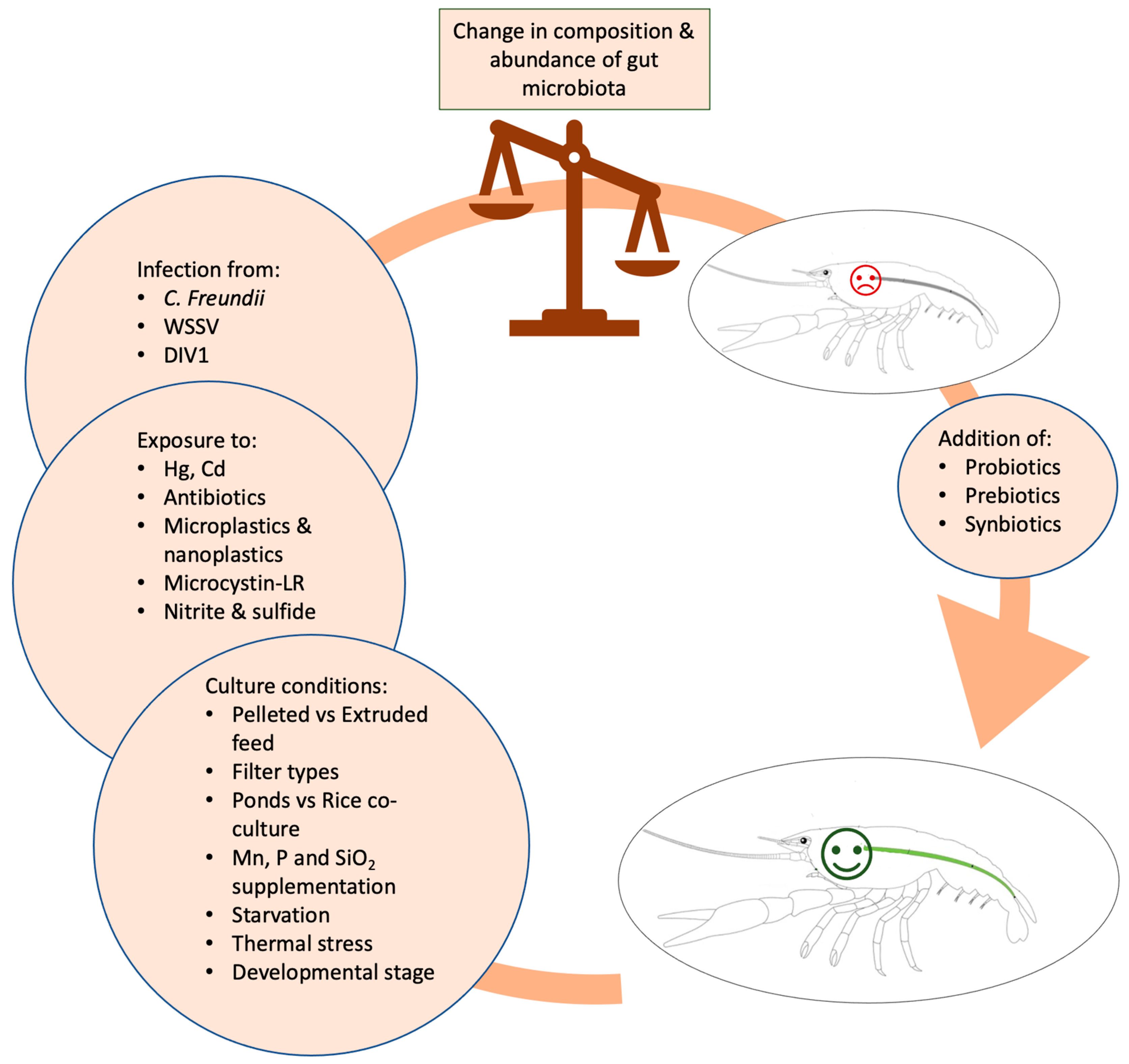
4. Main Factors Affecting Crayfish Microbiota Abundances and Composition
4.1. P. Clarkii
4.2. Cherax Genus
4.3. Other Genera
4.4. Overview of Factors Affecting Microbial Composition and Diversity and Limitations Existing
5. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020, in: Sustainability in Action. Rome, 2020. Available online: https://www.fao.org/documents/card/en/c/ca9229en (accessed on 9 April 2023).
- Edwards, P.; Zhang, W.; Belton, B.; Little, D.C. Misunderstandings, myths and mantras in aquaculture: Its contribution to world food supplies has been systematically over reported. Mar. Policy 2019, 106, 103547. [Google Scholar] [CrossRef]
- European Commission. The State of World Fisheries and Aquaculture (SOFIA) 2022. 2023. Available online: https://knowledge4policy.ec.europa.eu/publication/state-world-fisheries-aquaculture-sofia-2022_en (accessed on 9 April 2023).
- Subedi, B.; Shrestha, A. A review: Application of probiotics in aquaculture. Int. J. For. 2020, 4, 52–60. [Google Scholar]
- Rohani, M.F.; Islam, S.M.; Hossain, M.K.; Ferdous, Z.; Siddik, M.A.; Nuruzzaman, M.; Padeniya, U.; Brown, C.; Shahjahan, M. Probiotics, prebiotics and synbiotics improved the functionality of aquafeed: Upgrading growth, reproduction, immunity and disease resistance in fish. Fish Shellfish Immunol. 2022, 120, 569–589. [Google Scholar] [CrossRef]
- Tuan, T.N.; Duc, P.M.; Hatai, K. Overview of the use of probiotics in aquaculture. IJFA 2013, 3, 89–97. [Google Scholar]
- Garlock, T.; Asche, F.; Anderson, J.; Bjørndal, T.; Kumar, G.; Lorenzen, K.; Ropicki, A.; Smith, M.D.; Tveterås, R. A global blue revolution: Aquaculture growth across regions, species, and countries. Rev. Fish. Sci. Aquac. 2020, 28, 107–116. [Google Scholar] [CrossRef]
- Nakajima, T.; Hudson, M.J.; Uchiyama, J.; Makibayashi, K.; Zhang, J. Common carp aquaculture in Neolithic China dates back 8000 years. Nat. Ecol. Evol. 2019, 3, 1415–1418. [Google Scholar] [CrossRef]
- Alvanou, M.V.; Papadopoulos, D.K.; Lattos, A.; Georgoulis, I.; Feidantsis, K.; Apostolidis, A.P.; Michaelidis, B.; Giantsis, I.A. Biology, distribution, conservation status and stocking perspective of freshwater crayfish in Greece: An updated review. Aquac. Res. 2022, 53, 5115–5128. [Google Scholar] [CrossRef]
- Rubel, H.; Woods, W.; Pérez, D.; Meyer, A.; Felde, Z.; Zielcke, S.; Lidy, C. A Strategic Approach to Sustainable Shrimp Production in Thailand the Case for Improved Economics and Sustainability Carolin Lanfer 2. A Strategic Approach to Sustainable Shrimp Production in Thailand. Available online: https://media-publications.bcg.com/BCG-A-Strategic-Approach-to-Sustainable-Shrimp-Production-Thailand-July-2019.pdf (accessed on 9 April 2023).
- Wang, Q.; Ding, H.; Tao, Z.; Ma, D. Crayfish (Procambarus clarkii) cultivation in China: A decade of unprecedented development. In Aquaculture in China: Success Stories and Modern Trends; Gui, J.F., Tang, Q., Li, Z., Liu, J., De Silva, S.S., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2018; Volume 4, pp. 363–377. [Google Scholar]
- Shui, Y.; Kui, Y.; Zhu, G.Y.; Xu, Z.H. Egg incubation and plasma proteome alterations in response to brood care in maternal crayfish (Procambarus clarkii). Aquaculture 2021, 542, 736888. [Google Scholar] [CrossRef]
- Alvanou, M.V.; Kyriakoudi, A.; Makri, V.; Lattos, A.; Feidantsis, K.; Papadopoulos, D.; Georgoulis, I.; Apostolidis, A.P.; Michaelidis, B.; Mourtzinos, I.; et al. Effects of dietary substitution of fishmeal by black soldier fly (Hermetia illucens) meal on growth performance, whole-body chemical composition, and fatty acid profile of Pontastacus leptodactylus juveniles under captivity. Front. Physiol. 2023, 14, 501. [Google Scholar] [CrossRef]
- Ackefors, H.E. Freshwater crayfish farming technology in the 1990s: A European and global perspective. Fish Fish. 2000, 1, 337–359. [Google Scholar] [CrossRef]
- Savini, D.; Occhipinti-Ambrogi, A.; Marchini, A.; Tricarico, E.; Gherardi, F.; Olenin, S.; Gollasch, S. The top 27 animal alien species introduced into Europe for aquaculture and related activities. J. Appl. Ichthyol. 2010, 26, 1–7. [Google Scholar] [CrossRef]
- Holdich, D.M. Biology of Freshwater Crayfish; Blackwell Science: Oxford, UK, 2002. [Google Scholar]
- McClain, W.R. Crayfish aquaculture. Fish. Aquac. 2020, 9, 259. [Google Scholar]
- Stentiford, G.D.; Neil, D.M.; Peeler, E.J.; Shields, J.D.; Small, H.J.; Flegel, T.W.; Vlak, J.M.; Jones, B.; Morado, F.; Moss, S.; et al. Disease will limit future food supply from the global crustacean fishery and aquaculture sectors. J. Invertebr. Pathol. 2012, 110, 141–157. [Google Scholar] [CrossRef]
- Rajeev, R.; Adithya, K.; Kiran, G.S.; Selvin, J. Healthy microbiome: A key to successful and sustainable shrimp aquaculture. Rev. Aquac. 2021, 13, 238–258. [Google Scholar] [CrossRef]
- Thornber, K.; Verner-Jeffreys, D.; Hinchliffe, S.; Rahman, M.M.; Bass, D.; Tyler, C.R. Evaluating antimicrobial resistance in the global shrimp industry. Rev. Aquac. 2020, 12, 966–986. [Google Scholar] [CrossRef]
- CDC. Antibiotic Resistance Threats in the United States, 2019, 2019 AR Threats Report. CDC: U.S. Department of Health and Human Services; 2019. Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report-508.pdf (accessed on 25 April 2023).
- Relman, D.A.; Lipsitch, M. Microbiome as a tool and a target in the effort to address antimicrobial resistance. Proc. Natl. Acad. Sci. USA 2018, 115, 12902–12910. [Google Scholar] [CrossRef]
- Xiong, J.B.; Nie, L.; Chen, J. Current understanding on the roles of gut microbiota in fish disease and immunity. Zool. Res. 2019, 40, 70. [Google Scholar]
- Newman, A.M.; Arshad, M. The role of probiotics, prebiotics and synbiotics in combating multidrug-resistant organisms. Clin. Ther. 2020, 42, 1637–1648. [Google Scholar] [CrossRef]
- García-Márquez, J.; Tapia-Paniagua, S.; Moriñigo, M.Á.; Arijo, S. Probiotics for Controlling Infectious Diseases. In Probiotics in Aquacutlure; Austin, B., Sharifuzzaman, S., Eds.; Springer: Cham, Switzerland, 2022; pp. 103–129. [Google Scholar]
- Eissa, E.S.H.; Ahmed, R.A.; Abd Elghany, N.A.; Elfeky, A.; Saadony, S.; Ahmed, N.H.; Sakr, S.E.S.; Dayrit, G.B.; Tolenada, C.P.S.; Atienza, A.A.C.; et al. Potential Symbiotic Effects of β-1, 3 Glucan, and Fructooligosaccharides on the Growth Performance, Immune Response, Redox Status, and Resistance of Pacific White Shrimp, Litopenaeus vannamei to Fusarium solani Infection. Fishes 2023, 8, 105. [Google Scholar] [CrossRef]
- Sehnal, L.; Brammer-Robbins, E.; Wormington, A.M.; Blaha, L.; Bisesi, J.; Larkin, I.; Martyniuk, C.J.; Simonin, M.; Adamovsky, O. Microbiome composition and function in aquatic vertebrates: Small organisms making big impacts on aquatic animal health. Front. Microbiol. 2021, 12, 358. [Google Scholar] [CrossRef] [PubMed]
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Micha, R.; Wallace, S. Effects on coronary heart disease of increasing polyunsaturated fat in place of saturated fat: A systematic review and meta-analysis of randomized controlled trials. PLoS Med. 2010, 7, e1000252. [Google Scholar] [CrossRef]
- Willett, W.C. Trans fatty acids and cardiovascular disease—Epidemiological data. Atheroscler. Suppl. 2006, 7, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Mondal, K.; Haque, S. A review on application of probiotic, prebiotic and synbiotic for sustainable development of aquaculture. Growth 2017, 14, 15. [Google Scholar]
- Amenyogbe, E.; Chen, G.; Wang, Z.; Huang, J.; Huang, B.; Li, H. The exploitation of probiotics, prebiotics and synbiotics in aquaculture: Present study, limitations and future directions.: A review. Aquac. Int. 2020, 28, 1017–1041. [Google Scholar] [CrossRef]
- Butt, U.D.; Lin, N.; Akhter, N.; Siddiqui, T.; Li, S.; Wu, B. Overview of the latest developments in the role of probiotics, prebiotics and synbiotics in shrimp aquaculture. Fish Shellfish Immunol. 2021, 114, 263–281. [Google Scholar] [CrossRef]
- Interaminense, J.A.; Vogeley, J.L.; Gouveia, C.K.; Portela, R.S.; Oliveira, J.P.; Silva, S.M.; Coimbra, M.R.M.; Peixoto, S.M.; Soares, R.B.; Buarque, D.S.; et al. Effects of dietary Bacillus subtilis and Shewanella algae in expression profile of immune-related genes from hemolymph of Litopenaeus vannamei challenged with Vibrio parahaemolyticus. Fish Shellfish Immunol. 2019, 86, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Roy, S.; Meena, D.K.; Sarkar, U.K. Application of probiotics in shrimp aquaculture: Importance, mechanisms of action, and methods of administration. Rev. Fish. Sci. Aquac. 2016, 24, 342–368. [Google Scholar] [CrossRef]
- Ringø, E.; Doan, H.; Van Lee, S.; Song, S.K. Lactic acid bacteria in shellfish: Possibilities and challenges. Rev. Fish. Sci. Aquac. 2020, 28, 139–169. [Google Scholar] [CrossRef]
- Denev, S.; Beev, G.; Staykov, Y.; Moutafchieva, R. Microbial ecology of the gastrointestinal tract of fish and the potential application of probiotics and prebiotics in finfish aquaculture. Int. Aquat. Res. 2009, 1, 1. [Google Scholar]
- Cabello, F.C. Heavy use of prophylactic antibiotics in aquaculture: A growing problem for human and animal health and for the environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef]
- Tanwar, J.; Das, S.; Fatima, Z.; Hameed, S. Multidrug resistance: An emerging crisis. Interdiscip. Perspect. Infect. Dis. 2014, 2014, 541340. [Google Scholar] [CrossRef]
- Rekecki, A.; Dierckens, K.; Laureau, S.; Boon, N.; Bossier, P.; Van Den Broeck, W. Effect of germ-free rearing environment on gut development of larval sea bass (Dicentrarchus labrax L.). Aquaculture 2009, 293, 8–15. [Google Scholar] [CrossRef]
- Maynard, C.L.; Elson, C.O.; Hatton, R.D.; Weaver, C.T. Reciprocal interactions of the intestinal microbiota and immune system. Nature 2012, 489, 231–241. [Google Scholar] [CrossRef]
- Madani, N.S.H.; Adorian, T.J.; Farsani, H.G.; Hoseinifar, S.H. The effects of dietary probiotic Bacilli (Bacillus subtilis and Bacillus licheniformis) on growth performance, feed efficiency, body composition and immune parameters of whiteleg shrimp (Litopenaeus vannamei) postlarvae. Aquac. Res. 2018, 49, 1926–1933. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Khanongnuch, C.; Kanpiengjai, A.; Unban, K.; Van Van Doan, H.; Hoseinifar, S.H.; Khanongnuch, C.; Kanpiengjai, A.; Unban, K.; et al. Host-associated probiotics boosted mucosal and serum immunity, disease resistance and growth performance of Nile tilapia (Oreochromis niloticus). Aquaculture 2018, 491, 94–100. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Abdel-Daim, M.M.; Van Doan, H. Probiotic application for sustainable aquaculture. Rev. Aquac. 2019, 11, 907–924. [Google Scholar] [CrossRef]
- Selim, K.M.; Reda, R.M. Improvement of immunity and disease resistance in the Nile tilapia, Oreochromis niloticus, by dietary supplementation with Bacillus amyloliquefaciens. Fish Shellfish Immunol. 2015, 44, 496–503. [Google Scholar] [CrossRef]
- Ramos, M.A.; Batista, S.; Pires, M.A.; Silva, A.P.; Pereira, L.F.; Saavedra, M.J.; Ozório, R.O.A.; Rema, P. Dietary probiotic supplementation improves growth and the intestinal morphology of Nile tilapia. Animal 2017, 11, 1259–1269. [Google Scholar] [CrossRef]
- Burr, G.; Hume, M.; Neill, W.H.; Gatlin III, D.M. Effects of prebiotics on nutrient digestibility of a soybean-meal-based diet by red drum Sciaenops ocellatus (Linnaeus). Aquac. Res. 2008, 39, 1680–1686. [Google Scholar]
- Gómez, G.D.; Balcázar, J.L. A review on the interactions between gut microbiota and innate immunity of fish. FEMS Immunol. Med. Microbiol. 2008, 52, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, I.; Oliva-Teles, A.; Enes, P. Prebiotics as functional ingredients: Focus on Mediterranean fish aquaculture. Rev. Aquac. 2018, 10, 800–832. [Google Scholar] [CrossRef]
- Dawood, M.A.; Koshio, S.; Ishikawa, M.; El-Sabagh, M.; Yokoyama, S.; Wang, W.L.; Yukum, Z.; Olivier, A. Physiological response, blood chemistry profile and mucus secretion of red sea bream (Pagrus major) fed diets supplemented with Lactobacillus rhamnosus under low salinity stress. Fish Physiol. Biochem. 2017, 43, 179–192. [Google Scholar] [CrossRef]
- Martínez Cruz, P.; Ibáñez, A.L.; Monroy Hermosillo, O.A.; Ramírez Saad, H.C. Use of probiotics in aquaculture. ISRN Microbiol. 2012, 2012, 1–13. [Google Scholar] [CrossRef]
- Hai, N.V. The use of probiotics in aquaculture. J. Appl. Microbiol. 2015, 119, 917–935. [Google Scholar] [CrossRef]
- Akrami, R.; Nasri-Tajan, M.; Jahedi, A.; Jahedi, M.; Razeghi Mansour, M.; Jafarpour, S.A. Effects of dietary synbiotic on growth, survival, lactobacillus bacterial count, blood indices and immunity of beluga (Huso huso Linnaeus, 1754) juvenile. Aquac. Nutr. 2015, 21, 952–959. [Google Scholar] [CrossRef]
- Van Doan, H.; Soltani, M.; Ringø, E. In vitro antagonistic effect and in vivo protective efficacy of Gram-positive probiotics versus Gram-negative bacterial pathogens in finfish and shellfish. Aquaculture 2021, 540, 736581. [Google Scholar] [CrossRef]
- Feckaninova, A.; Koscova, J.; Mudronova, D.; Popelka, P.; Toropilova, J. The use of probiotic bacteria against Aeromonas infections in salmonid aquaculture. Aquaculture 2017, 469, 1–8. [Google Scholar] [CrossRef]
- Ina-Salwany, M.Y.; Al-saari, N.; Mohamad, A.; Mursidi, F.A.; Mohd-Aris, A.; Amal, M.N.A.; Kasai, H.; Mino, S.; Sawabe, T.; Zamri-Saad, M. Vibriosis in fish: A review on disease development and prevention. J. Aquat. Anim. Health 2019, 31, 2–22. [Google Scholar] [CrossRef]
- Lyapparaj, P.; Maruthiah, T.; Ramasubburayan, R.; Prakash, S.; Kumar, C.; Immanuel, G.; Palavesam, H. Optimization of bacteriocin production by Lactobacillus sp. MSU3IR against shrimp bacterial pathogens. Aquatic Biosyst. 2013, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Jastaniah, S.D.S.; Hafsan, H.; Tseng, C.J.; Karim, Y.S.; Hamza, M.U.; Hameed, N.M.; Al-Zubaidi, S.H.; Almotlaq, S.S.K.; Yasin, G.; Iswanto, A.; et al. Effects of Dietary Pectin and Lactobacillus salivarius ATCC 11741 on Growth Performance, Immunocompetence, Gut Microbiota, Antioxidant Capacity, and Disease Resistance in Narrow-Clawed Crayfish, Postantacus leptodactylus. Aquac. Nutr. 2022, 2022, 13. [Google Scholar] [CrossRef]
- Foysal, M.J.; Fotedar, R.; Siddik, M.A.; Tay, A. Lactobacillus acidophilus and L. plantarum improve health status, modulate gut microbiota and innate immune response of marron (Cherax cainii). Sci. Rep. 2020, 10, 5916. [Google Scholar] [CrossRef]
- Soltani, M.; Ghosh, K.; Hoseinifar, S.H.; Kumar, V.; Lymbery, A.L.; Roy, S.; Ringø, E. Genus bacillus, promising probiotics in aquaculture: Aquatic animal origin, bio-active components, bioremediation and efficacy in fish and shellfish. Rev. Fish. Sci. Aquac. 2019, 27, 331–379. [Google Scholar] [CrossRef]
- Hayakijkosol, O.; Owens, L.; Picard, J. Case report of bacterial infections in a redclaw crayfish (Cherax quadricarinatus) hatchery. Aquaculture 2017, 475, 1–7. [Google Scholar] [CrossRef]
- Valipour, A.; Nedaei, S.; Noori, A.; Khanipour, A.A.; Hoseinifar, S.H. Dietary Lactobacillus plantarum affected on some immune parameters, air-exposure stress response, intestinal microbiota, digestive enzyme activity and performance of narrow clawed crayfish (Astacus leptodactylus, Eschscholtz). Aquaculture 2019, 504, 121–130. [Google Scholar] [CrossRef]
- Xu, L.; Yuan, J.; Chen, X.; Zhang, S.; Xie, M.; Chen, C.; Wu, Z. Screening of intestinal probiotics and the effects of feeding probiotics on the digestive enzyme activity, immune, intestinal flora and WSSV resistance of Procambarus clarkii. Aquaculture 2021, 540, 736748. [Google Scholar] [CrossRef]
- Zhu, L.; Kong, Y.; Chang, X.; Feng, J.; Wang, X.; Hou, L.; Zhao, X.; Pei, C.; Kong, X. Effects of two fish-derived probiotics on growth performance, innate immune response, intestinal health, and disease resistance of Procambarus clarkii. Aquaculture 2023, 562, 738765. [Google Scholar] [CrossRef]
- Han, M.; Gao, T.; Liu, G.; Zhu, C.; Zhang, T.; Sun, M.; Li, J.; Ji, F.; Si, Q.; Jiang, Q. The effect of a polystyrene nanoplastic on the intestinal microbes and oxidative stress defense of the freshwater crayfish, Procambarus clarkii. Sci. Total Environ. 2022, 833, 155722. [Google Scholar] [CrossRef]
- Foysal, M.J.; Chua, E.G.; Gupta, S.K.; Lamichhane, B.; Tay, C.Y.; Fotedar, R. Bacillus mycoides supplemented diet modulates the health status, gut microbiota and innate immune response of freshwater crayfish marron (Cherax cainii). JAFST 2020, 262, 114408. [Google Scholar] [CrossRef]
- Ambas, I.; Suriawan, A.; Fotedar, R. Immunological responses of customised probiotics-fed marron, Cherax tenuimanus, (Smith 1912) when challenged with Vibrio mimicus. Fish Shellfish Immunol. 2013, 35, 262–270. [Google Scholar] [CrossRef]
- Lai, Y.; Luo, M.; Zhu, F. Dietary Bacillus amyloliquefaciens enhance survival of white spot syndrome virus infected crayfish. Fish Shellfish Immunol. 2020, 102, 161–168. [Google Scholar] [CrossRef]
- Foysal, M.J.; Fotedar, R.; Siddik, M.A.; Chaklader, M.R.; Tay, A. Lactobacillus plantarum in black soldier fly (Hermetica illucens) meal modulates gut health and immunity of freshwater crayfish (Cherax cainii). Fish Shellfish Immunol. 2021, 108, 42–52. [Google Scholar] [CrossRef]
- Javier, V.S.; Guadalupe, D.R.M.; Mauricio, G.M.; Andrea, B.A.; Alberto, R.O.D. Effect of live food enriched with the probiotic Spomune©, on the survival and growth of Cambarellus montezumae (Saussure, 1857) under controlled conditions. Int. J. Fish. Aquat. Species 2021, 9, 56–62. [Google Scholar]
- Carreño-León, D.; Racotta-Dimitrov, I.; Casillas-Hernández, R.; Monge-Quevedo, A.; Ocampo-Victoria, L.; Naranjo-Páramo, J.; Villarreal, H. Growth, metabolic and physiological response of juvenile Cherax quadricarinatus fed different available nutritional substrates. J. Aquac. Res. Dev. 2014, 5, 1. [Google Scholar]
- Xu, Y.; Li, Y.; Xue, M.; Yang, T.; Luo, X.; Fan, Y.; Meng, Y.; Liu, W.; Lin, G.; Li, B.; et al. Effects of dietary Saccharomyces cerevisiae YFI-SC2 on the growth performance, intestinal morphology, immune parameters, intestinal microbiota, and disease resistance of Crayfish (Procambarus clarkia). Animals 2021, 11, 1963. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, M.; Ji, T.; Zhang, Y.; Wei, W.; Liu, Q. Bacillus subtilis CK3 used as an aquatic additive probiotics enhanced the immune response of crayfish Procambarus clarkii against newly identified Aeromonas veronii pathogen. Aquac. Res. 2022, 53, 255–264. [Google Scholar] [CrossRef]
- Castro-Mejía, J.; Castro-Mejía, G.; Alcántara-Muñiz, N.F.; Pacheco-Goómez, V.; Rodríguez-Estrada, E. Comparison of weight increase in Cambarellus montezumae (Saussure, 1857) fed with a diet enriched with probiotic. E BIOS 2011, 1, 20–26. [Google Scholar]
- Özdoğan, H.B.E.; Koca, S.B.; Özmen, Ö.; Ekinci, K.; Ekinci, E.; Koca, H.U.; Yiğit, N.Ö. Effect of feed supplementation with effective microorganisms (em) bokashi on hepatopancreas and gut histology, growth performance, and survival rate of freshwater crayfish Pontastacus leptodactylus (Eschscholtz, 1823). Turk. J. Vet. Anim. Sci. 2022, 46, 396–402. [Google Scholar] [CrossRef]
- Dıdınen, B.I.; Bahadır Koca, S.; Metin, S.E.Ç.İ.L.; Dıler, O.; Erol, K.G.; Dulluc, A.; Koca, H.U.; Yigit, N.O.; Ozkok, R.; Kucukkara, R. Effect of lactic acid bacteria and the potential probiotic Hafnia alvei on growth and survival rates of narrow clawed crayfish (Astacus leptodactylus Esch., 1823) stage II juveniles. Iran. J. Fish. Sci. 2016, 15, 1307–1317. [Google Scholar]
- Foysal, M.J.; Nguyen, T.T.T.; Chaklader, M.R.; Siddik, M.A.; Tay, C.Y.; Fotedar, R.; Gupta, S.K. Marked variations in gut microbiota and some innate immune responses of freshwater crayfish, marron (Cherax cainii, Austin 2002) fed dietary supplementation of Clostridium butyricum. PeerJ 2019, 7, e7553. [Google Scholar] [CrossRef] [PubMed]
- Pronina, G.; Shishanova, E.; Isaev, D.; Tarazanova, T.V.; Prokhorov, A.A. Improving the aquatic organisms immune resistance with probiotics for the aquaculture sustainable development. IOP Conf. Ser Earth Environ. Sci. 2021, 937, 032031. [Google Scholar] [CrossRef]
- Ambas, I.; Fotedar, R.; Buller, N. Survival and immunity of marron Cherax cainii (Austin, 2002) fed Bacillus mycoides supplemented diet under simulated transport. J. Aquac. Res. Dev. 2015, 7, 1–6. [Google Scholar]
- Nedaei, S.; Noori, A.; Valipour, A.; Khanipour, A.A.; Hoseinifar, S.H. Effects of dietary galactooligosaccharide enriched commercial prebiotic on growth performance, innate immune response, stress resistance, intestinal microbiota and digestive enzyme activity in Narrow clawed crayfish (Astacus leptodactylus Eschscholtz, 1823). Aquaculture 2019, 499, 80–89. [Google Scholar]
- Safari, O.; Paolucci, M.; Motlagh, H.A. Dietary supplementation of Chlorella vulgaris improved growth performance, immunity, intestinal microbiota and stress resistance of juvenile narrow clawed crayfish, Pontastacus leptodactylus Eschscholtz, 1823. Aquaculture 2022, 554, 738138. [Google Scholar] [CrossRef]
- Safari, O.; Shahsavani, D.; Paolucci, M.; Atash, M.M.S. Single or combined effects of fructo-and mannan oligosaccharide supplements on the growth performance, nutrient digestibility, immune responses and stress resistance of juvenile narrow clawed crayfish, Astacus leptodactylus leptodactylus Eschscholtz, 1823. Aquaculture 2014, 432, 192–203. [Google Scholar] [CrossRef]
- Cai, M.; Hui, W.; Deng, X.; Wang, A.; Hu, Y.; Liu, B.; Chen, K.; Liu, F.; Tian, H.; Gu, X.; et al. Dietary Haematococcus pluvialis promotes growth of red swamp crayfish Procambarus clarkii (Girard, 1852) via positive regulation of the gut microbial co-occurrence network. Aquaculture 2022, 551, 737900. [Google Scholar] [CrossRef]
- Duan, H.; Zuo, J.; Pan, N.; Cui, X.; Guo, J.; Sui, L. 3-Hydroxybutyrate helps crayfish resistant to Vibrio parahaemolyticus infection in versatile ways. Fish Shellfish Immunol. 2023, 132, 108444. [Google Scholar] [CrossRef]
- Sang, H.M.; Fotedar, R. Dietary supplementation of mannan oligosaccharide improves the immune responses and survival of marron, Cherax tenuimanus (Smith, 1912) when challenged with different stressors. Fish Shellfish Immunol. 2009, 27, 341–348. [Google Scholar] [CrossRef]
- Huang, Q.; Zhu, Y.; Yu, J.; Fang, L.; Li, Y.; Wang, M.; Liu, J.; Yan, P.; Xia, J.; Liu, G.; et al. Effects of sulfated β-glucan from Saccharomyces cerevisiae on growth performance, antioxidant ability, nonspecific immunity, and intestinal flora of the red swamp crayfish (Procambarus clarkii). Fish Shellfish Immunol. 2022, 127, 891–900. [Google Scholar] [CrossRef]
- Sang, H.M.; Fotedar, R.; Filer, K. Effects of dietary mannan oligosaccharide on the survival, growth, immunity and digestive enzyme activity of freshwater crayfish, Cherax destructor Clark (1936). Aquac. Nutr. 2011, 17, e629–e635. [Google Scholar] [CrossRef]
- Safari, O.; Paolucci, M. Modulation of growth performance, immunity, and disease resistance in narrow-clawed crayfish, Astacus leptodactylus leptodactylus (Eschscholtz, 1823) upon synbiotic feeding. Aquaculture 2017, 479, 333–341. [Google Scholar] [CrossRef]
- Safari, O.; Paolucci, M.; Motlagh, H.A. Effects of synbiotics on immunity and disease resistance of narrow-clawed crayfish, Astacus leptodactylus leptodactylus (Eschscholtz, 1823). Fish Shellfish Immunol. 2017, 64, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Mona, M.H.; Rizk, E.S.T.; Salama, W.M.; Younis, M.L. Efficacy of probiotics, prebiotics, and immunostimulant on growth performance and immunological parameters of Procambarus clarkii juveniles. JOBAZ 2015, 69, 17–25. [Google Scholar] [CrossRef]
- Siddik, M.A.; Fotedar, R.; Chaklader, M.R.; Foysal, M.J.; Nahar, A.; Howieson, J. Fermented animal source protein as substitution of fishmeal on intestinal microbiota, immune-related cytokines and resistance to Vibrio mimicus in freshwater crayfish (Cherax cainii). Front.Physiol. 2020, 10, 1635. [Google Scholar] [CrossRef]
- Oktaviana, A.; Febriani, D. Additional Lactobacillus and Coconut Powder to Increase Growth and Survival Rate on Cherax sp. IOP Conf. Ser: Earth Environ. Sci. 2022, 1012, 012045. [Google Scholar] [CrossRef]
- Bœuf, G.; Payan, P. How should salinity influence fish growth? Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 130, 411–423. [Google Scholar] [CrossRef]
- Ansaldo, M.; Luquet, C.M.; Evelson, P.A.; Polo, J.M.; Llesuy, S. Antioxidant levels from different Antarctic fish caught around South Georgia Island and Shag Rocks. Polar Biol. 2000, 23, 160–165. [Google Scholar] [CrossRef]
- Holt, C.C.; Bass, D.; Stentiford, G.D.; van der Giezen, M. Understanding the role of the shrimp gut microbiome in health and disease. J. Invertebr. Pathol. 2021, 186, 107387. [Google Scholar] [CrossRef]
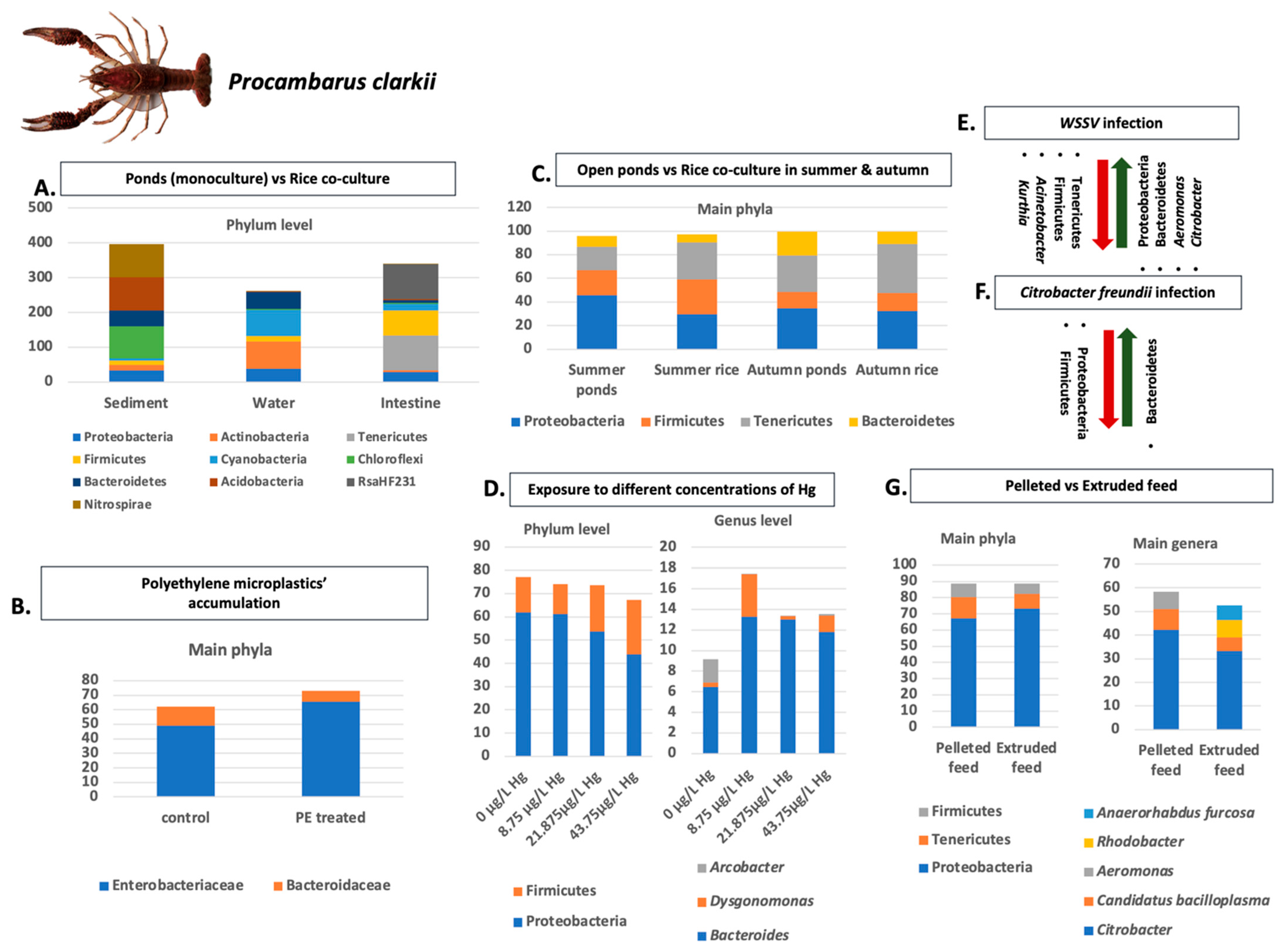
- Wang, Y.; Wang, C.; Chen, Y.; Zhang, D.; Zhao, M.; Li, H.; Guo, P. Microbiome analysis reveals microecological balance in the emerging rice–crayfish integrated breeding mode. Front. Microbiol. 2021, 12, 669570. [Google Scholar] [CrossRef]
- Liu, Q.; Long, Y.N.; Li, B.; Zhao, L.; Luo, J.; Xu, L.; Luo, W.; Du, Z.; Zhou, J.; Yang, S. Rice-shrimp culture: A better intestinal microbiota, immune enzymatic activities, and muscle relish of crayfish (Procambarus clarkii) in Sichuan Province. Appl. Microbiol. Biotechnol. 2020, 104, 9413–9420. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Jiang, N.; Fan, Y.; Yang, T.; Li, M.; Liu, W.; Li, Y.; Li, B.; Zeng, L.; Zhou, Y. White spot syndrome virus (WSSV) infection alters gut histopathology and microbiota composition in crayfish (Procambarus clarkii). Aquac. Rep. 2022, 22, 101006. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, Y.; Song, Z.; Liang, H.; Zhong, S.; Yu, Y.; Liu, T.; Sha, H.; He, L.; Gan, J. Mercury Induced Tissue Damage, Redox Metabolism, Ion Transport, Apoptosis, and Intestinal Microbiota Change in Red Swamp Crayfish (Procambarus clarkii): Application of Multi-Omics Analysis in Risk Assessment of Hg. Antioxidants 2022, 11, 1944. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jin, Z.; Shen, M.; Chang, Z.; Yu, G.; Wang, L.; Xia, X. Accumulation of polyethylene microplastics induces oxidative stress, microbiome dysbiosis and immunoregulation in crayfish. Fish Shellfish Immunol. 2022, 125, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Xi, Q.; Tang, J.; Liu, T.; Liu, C.; Li, H.; Gu, X.; Shen, M.; Zhang, M.; Fang, J.; et al. Effects of Pelleted and Extruded Feed on Growth Performance, Intestinal Histology and Microbiota of Juvenile Red Swamp Crayfish (Procambarus clarkii). Animals 2022, 12, 2252. [Google Scholar] [CrossRef]
- Li, M.; Wang, J.; Deng, H.; Li, L.; Huang, X.; Chen, D.; Ouyang, P.; Geng, Y.; Yang, S.; Yin, L.; et al. The Damage of the Crayfish (Procambarus Clarkii) Digestive Organs Caused by Citrobacter Freundii Is Associated with the Disturbance of Intestinal Microbiota and Disruption of Intestinal-Liver Axis Homeostasis. Front. Cell. Infect. Microbiol. 2022, 12, 940576. [Google Scholar] [CrossRef] [PubMed]
- Boets, P.; Lock, K.; Cammaerts, R.; Plu, D.; Goethals, P.L. Occurrence of the invasive crayfish Procambarus clarkii (Girard, 1852) in Belgium (Crustacea: Cambaridae). Belg. J. Zool. 2009, 139, 173–175. [Google Scholar]
- Mo, A.; Li, H.; Zhai, Y.; Yang, H.; Wang, J.; Wang, Y.; Yi, L.; Guo, W.; Yuan, Y. Assessment of Bacillus subtilis applied in rice-crayfish coculture system on physicochemical properties, microbial sulfur cycling, Cd accumulation and bioavailability. J. Clean. Prod. 2022, 381, 135158. [Google Scholar] [CrossRef]
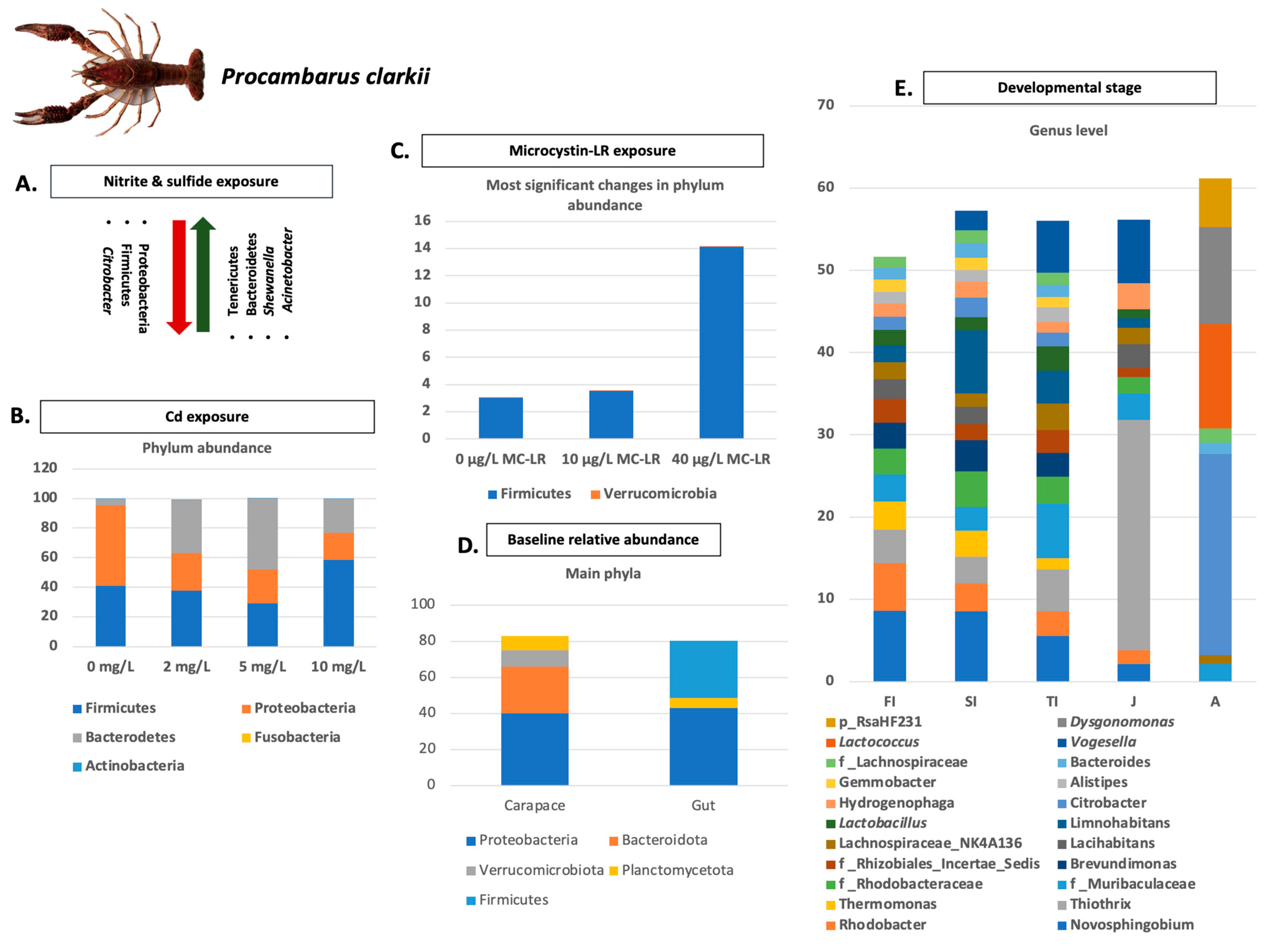
- Guo, K.; Ruan, G.; Fan, W.; Fang, L.; Wang, Q.; Luo, M.; Yi, T. The effect of nitrite and sulfide on the antioxidant capacity and microbial composition of the intestines of red swamp crayfish, Procambarus clarkii. Fish Shellfish Immunol. 2020, 96, 290–296. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Z.; Kholodkevich, S.; Sharov, A.; Feng, Y.; Ren, N.; Sun, K. Microcystin-LR-induced changes of hepatopancreatic transcriptome, intestinal microbiota, and histopathology of freshwater crayfish (Procambarus clarkii). Sci. Total Environ. 2020, 711, 134549. [Google Scholar] [CrossRef]
- Xie, M.; Zhang, S.; Xu, L.; Wu, Z.; Yuan, J.; Chen, X. Comparison of the intestinal microbiota during the different growth stages of red swamp crayfish (Procambarus clarkii). Front. Microbiol. 2021, 12, 696281. [Google Scholar] [CrossRef] [PubMed]
- Xavier, R.; Soares, M.C.; Silva, S.M.; Banha, F.; Gama, M.; Ribeiro, L.; Anastácio, P.; Cardoso, S.C. Environment and host-related factors modulate gut and carapace bacterial diversity of the invasive red swamp crayfish (Procambarus clarkii). Hydrobiology 2021, 848, 4045–4057. [Google Scholar] [CrossRef]
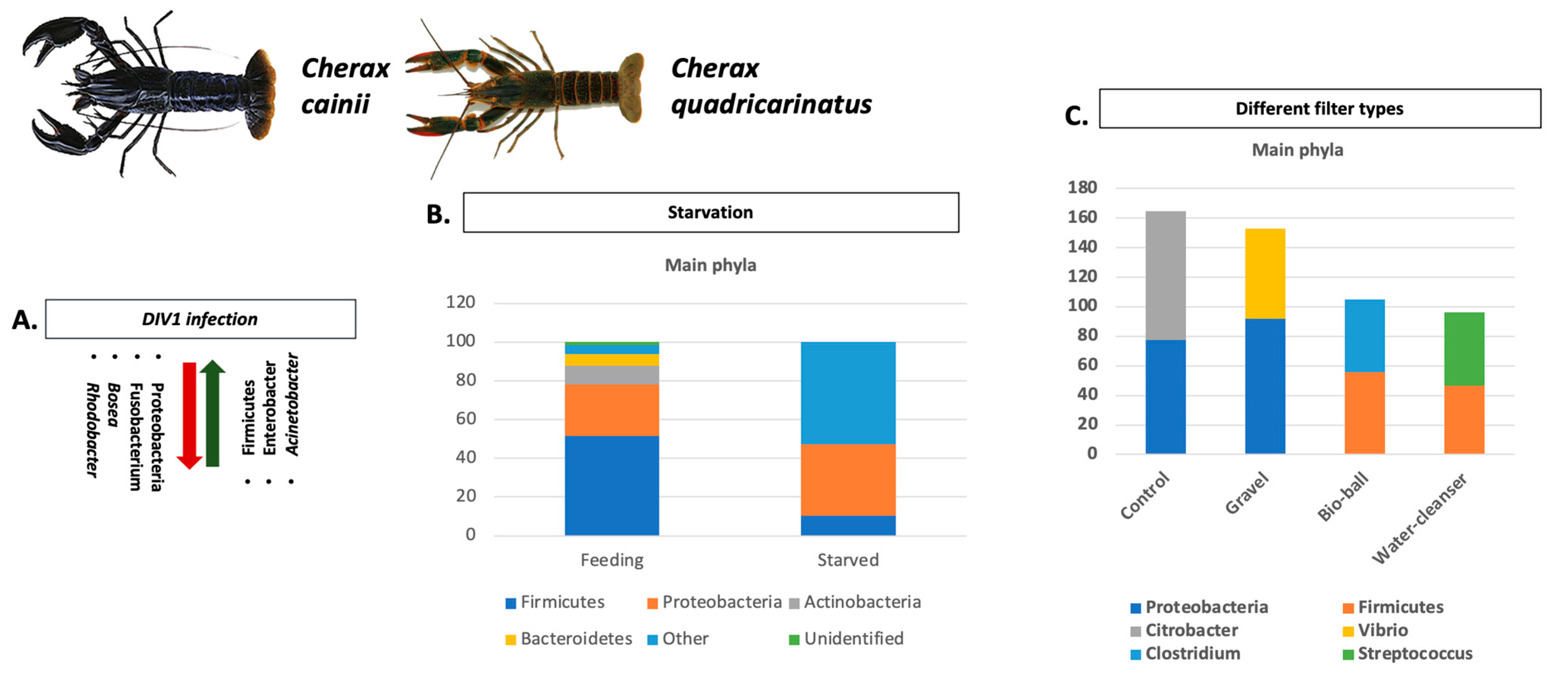
- Foysal, M.J.; Fotedar, R.; Tay, A.C.Y.; Gupta, S.K. Effects of long-term starvation on health indices, gut microbiota and innate immune response of freshwater crayfish, marron (Cherax cainii, Austin 2002). Aquaculture 2020, 514, 734444. [Google Scholar] [CrossRef]
- Zheng, J.; Jia, Y.; Li, F.; Chi, M.; Cheng, S.; Liu, S.; Jiang, W.; Liu, Y. Changes in the gene expression and gut microbiome to the infection of decapod iridescent virus 1 in Cherax quadricarinatus. Fish Shellfish Immunol. 2023, 132, 108451. [Google Scholar] [CrossRef] [PubMed]
- Foysal, M.J.; Fotedar, R.; Tay, C.Y.; Gupta, S.K. Biological filters regulate water quality, modulate health status, immune indices and gut microbiota of freshwater crayfish, marron (Cherax cainii, Austin, 2002). Chemosphere 2020, 247, 125821. [Google Scholar] [CrossRef]
- Cherax cainii. Available online: https://www.fish.wa.gov.au/species/marron/pages/default.aspx (accessed on 25 April 2023).
- Belle, C.C.; Yeo, D.C. New observations of the exotic Australian red-claw crayfish, Cherax quadricarinatus (von Martens, 1868) (Crustacea: Decapoda: Parastactidae) in Singapore. NiS 2010, 3, 99–102. [Google Scholar]
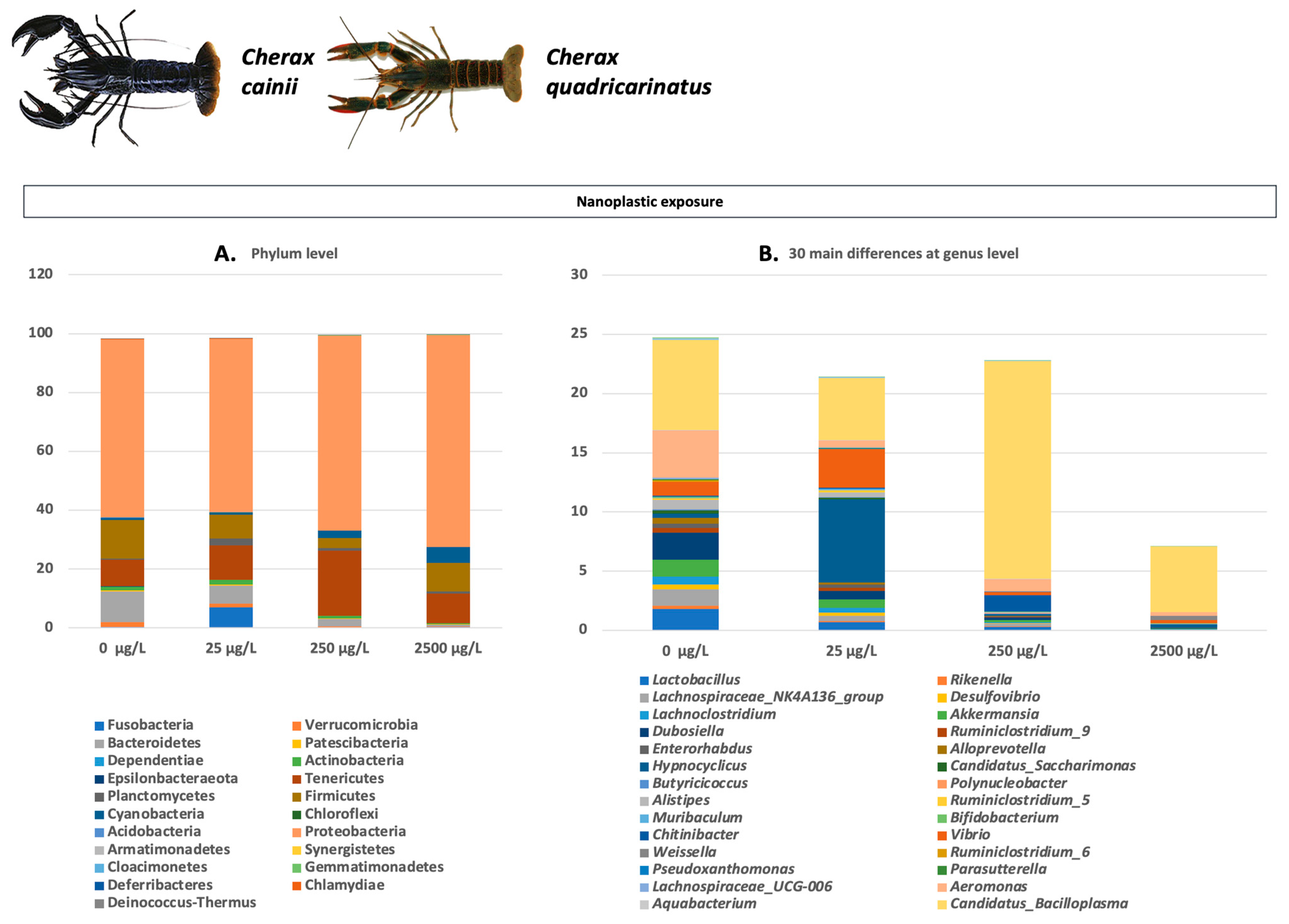
- Cheng, H.; Dai, Y.; Ruan, X.; Duan, X.; Zhang, C.; Li, L.; Huang, F.; Shan, J.; Liang, K.; Jia, X.; et al. Effects of nanoplastic exposure on the immunity and metabolism of red crayfish (Cherax quadricarinatus) based on high-throughput sequencing. Ecotoxicol. Environ. Saf. 2022, 245, 114114. [Google Scholar] [CrossRef]
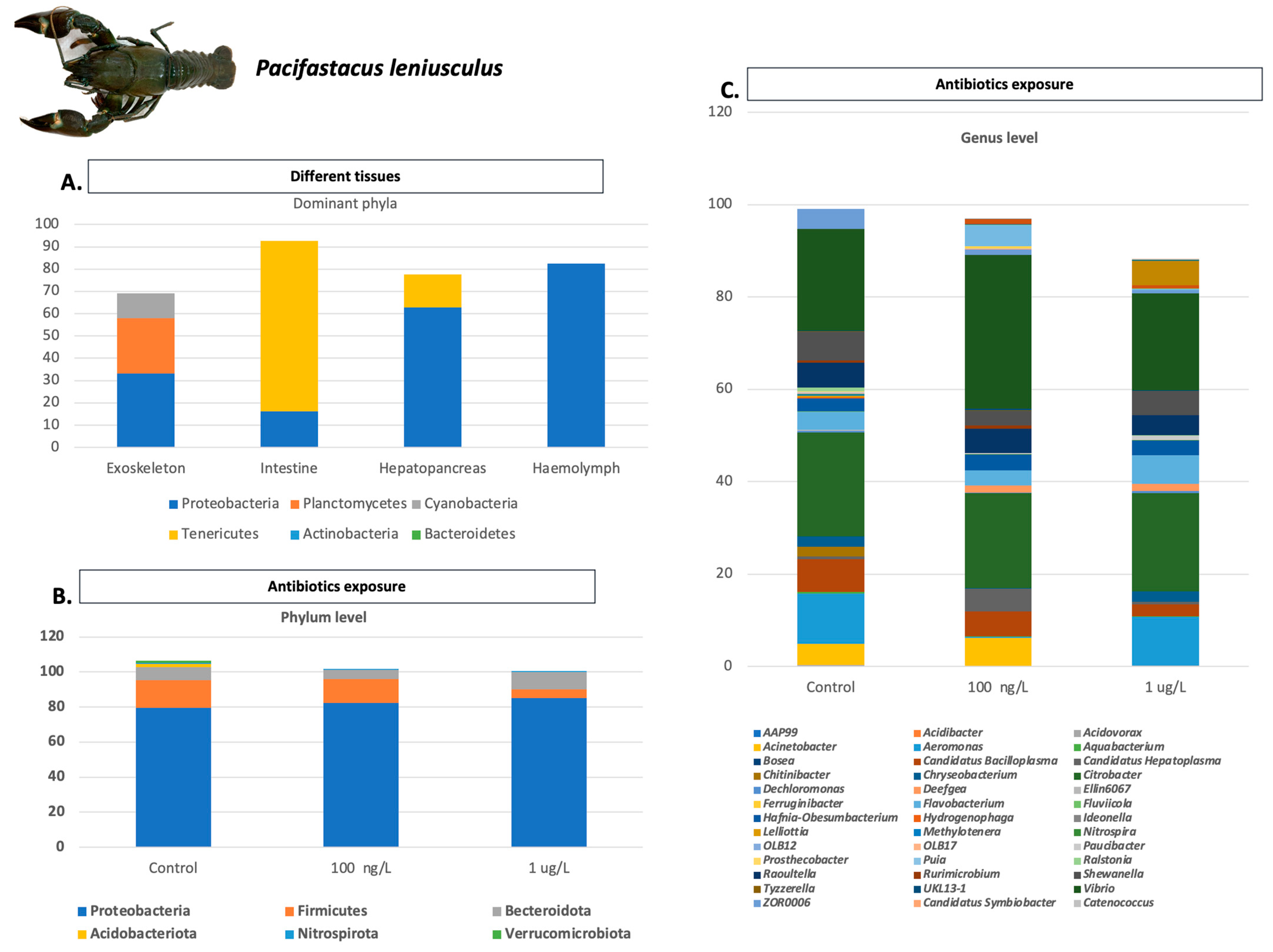
- Hernandez-Perez, A.; Zamora-Briseno, J.A.; Söderhäll, K.; Söderhäll, I. Gut microbiome alterations in the crustacean Pacifastacus leniusculus exposed to environmental concentrations of antibiotics and effects on susceptibility to bacteria challenges. DCI 2022, 126, 104181. [Google Scholar] [CrossRef]
- Dragičević, P.; Bielen, A.; Petrić, I.; Vuk, M.; Žučko, J.; Hudina, S. Microbiome of the successful freshwater invader, the signal crayfish, and its changes along the invasion range. Microbiol. Spectr. 2021, 9, e00389-21. [Google Scholar] [CrossRef]
- Jin, S.; Jacquin, L.; Xiong, M.; Li, R.; Lek, S.; Li, W.; Zhang, T. Reproductive pattern and population dynamics of commercial red swamp crayfish (Procambarus clarkii) from China: Implications for sustainable aquaculture management. PeerJ 2019, 7, e6214. [Google Scholar] [CrossRef]
- Chen, X.; Fan, L.; Qiu, L.; Dong, X.; Wang, Q.; Hu, G.; Meng, S.; Li, D.; Chen, J. Metagenomics Analysis Reveals Compositional and Functional Differences in the Gut Microbiota of Red Swamp Crayfish, Procambarus clarkii, Grown on Two Different Culture Environments. Front. Microbiol. 2021, 12, 735190. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Li, M.; Huang, Y.; Yang, H.; Geng, Y.; Ouyang, P.; Chen, D.; Yin, L.; Yang, S.; Jiang, J.; et al. Microbiome analysis reveals microecological advantages of emerging ditchless rice-crayfish co-culture mode. Front. Microbiol. 2022, 13, 892026. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Li, M.; Duan, H.; Li, L.; Ouyang, P.; Chen, D.; Geng, Y.; Huang, X.; Yang, S.; Yin, L.; et al. Microbial analysis reveals the potential colonization of pathogens in the intestine of crayfish (Procambarus clarkii) in traditional aquaculture environments. Ecotoxicol. Environ. Saf. 2021, 224, 112705. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhang, Q.; Zhang, T.; Chen, J.; Wang, S.; Hao, J.; Lin, Y.; Li, A. Association of the microbiota dysbiosis in the hepatopancreas of farmed crayfish (Procambarus clarkii) with disease outbreaks. Aquaculture 2021, 536, 736492. [Google Scholar] [CrossRef]
- Chen, H.; Wang, Y.; Zhang, J.; Bao, J. Intestinal microbiota in white spot syndrome virus infected red swamp crayfish (Procambarus clarkii) at different health statuses. Aquaculture 2021, 542, 736826. [Google Scholar] [CrossRef]
- Li, P.; Wu, G.; Li, Y.; Hu, C.; Ge, L.; Zheng, X.; Zhang, J.; Chen, J.; Zhang, H.; Bai, N.; et al. Long-term rice-crayfish-turtle co-culture maintains high crop yields by improving soil health and increasing soil microbial community stability. Geoderma 2022, 413, 115745. [Google Scholar] [CrossRef]
- Ruan, G.; Li, S.; He, N.; Fang, L.; Wang, Q. Short-term adaptability to non-hyperthermal stress: Antioxidant, immune and gut microbial responses in the red swamp crayfish, Procambarus clarkii. Aquaculture 2022, 560, 738497. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Z.; Kholodkevich, S.; Sharov, A.; Chen, C.; Feng, Y.; Ren, N.; Sun, K. Effects of cadmium on intestinal histology and microbiota in freshwater crayfish (Procambarus clarkii). Chemosphere 2020, 242, 125105. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, J.; Jin, X.; Liu, C.; Fan, C.; Guo, L.; Liang, Y.; Zheng, J.; Peng, N. Developmental, dietary, and geographical impacts on gut microbiota of red swamp crayfish (Procambarus clarkii). Microorganisms 2020, 8, 1376. [Google Scholar] [CrossRef]
- Foysal, M.J.; Nguyen, T.T.T.; Sialumano, M.; Phiri, S.; Chaklader, M.R.; Fotedar, R.; Gagnon, M.M.; Tay, A. Zeolite mediated processing of nitrogenous waste in the rearing environment influences gut and sediment microbial community in freshwater crayfish (Cherax cainii) culture. Chemosphere 2022, 298, 134276. [Google Scholar] [CrossRef]
- Tulsankar, S.S.; Foysal, M.J.; Cole, A.J.; Gagnon, M.M.; Fotedar, R. A mixture of manganese, silica and phosphorus supplementation alters the plankton density, species diversity, gut microbiota and improved the health status of cultured marron (Cherax cainii, Austin and Ryan, 2002). Biol. Trace Elem. Res. 2022, 200, 1383–1394. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Ji, L.; Cheng, M.; Wei, H.; Wang, Z.; Ning, K. Sustainability of the rice-crayfish co-culture aquaculture model: Microbiome profiles based on multi-kingdom analyses. Environ. Microbiome 2022, 17, 27. [Google Scholar] [CrossRef]
- Chen, L.; Xu, J.; Wan, W.; Xu, Z.; Hu, R.; Zhang, Y.; Zheng, J.; Gu, Z. The microbiome structure of a rice-crayfish integrated breeding model and its association with crayfish growth and water quality. Microbiol. Spectr. 2022, 10, e02204-21. [Google Scholar] [CrossRef] [PubMed]
- Parrillo, L.; Coccia, E.; Volpe, M.G.; Siano, F.; Pagliarulo, C.; Scioscia, E.; Varricchio, E.; Safari, O.; Eroldogan, T.; Paolucci, M. Olive mill wastewater-enriched diet positively affects growth, oxidative and immune status and intestinal microbiota in the crayfish, Astacus leptodactylus. Aquaculture 2017, 473, 161–168. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Ringø, E.; Esteban, M.A.; Dadar, M.; Dawood, M.A.O.; Faggio, C. Host- associated probiotics: A key factor in sustainable aquaculture. Rev. Fish. Sci. Aquacult. 2020, 28, 16–42. [Google Scholar] [CrossRef]
- Dong, X.; Li, Z.; Wang, X.; Zhou, M.; Lin, L.; Zhou, Y.; Li, J. Characteristics of Vibrio parahaemolyticus isolates obtained from crayfish (Procambarus clarkii) in freshwater. Int. J. Food Microbiol. 2016, 238, 132–138. [Google Scholar] [CrossRef]
- Jiravanichpaisal, P.; Roos, S.; Edsman, L.; Liu, H.; Söderhäll, K. A highly virulent pathogen, Aeromonas hydrophila, from the freshwater crayfish Pacifastacus leniusculus. J. Invertebr. Pathol. 2009, 101, 56–66. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, J.M.; Zhou, Y.L.; Almeida, A.; Finn, R.D.; Danchin, A.; He, L.S. Phylogenomics of expanding uncultured environmental Tenericutes provides insights into their pathogenicity and evolutionary relationship with Bacilli. BMC Genom. 2020, 21, 1–12. [Google Scholar] [CrossRef]
- Ringø, E.; Doan, H.V.; Lee, S.O.; Soltani, M.; Hoseinifar, S.H.; Harikrishnan, R.; Song, S.K. Probiotics, lactic acid bacteria and bacilli: Interesting supplementation for aquaculture. J. Appl. Microbiol. 2020, 129, 116–136. [Google Scholar] [CrossRef]
- Ringø, E. Probiotics in shellfish aquaculture. Aquac. Fish. 2020, 5, 1–27. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Sun, Y.Z.; Wang, A.; Zhou, Z. Probiotics as means of diseases control in aquaculture, a review of current knowledge and future perspectives. Front. Microbiol. 2018, 9, 2429. [Google Scholar] [CrossRef] [PubMed]
- Akhter, N.; Wu, B.; Memon, A.M.; Mohsin, M. Probiotics and prebiotics associated with aquaculture: A review. Fish Shellfish Immunol. 2015, 45, 733–741. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, M.T.; Alagawany, M.; Patra, A.K.; Kar, I.; Tiwari, R.; Dawood, M.A.; Dhama, K.; Abdel-Latif, H.M. The functionality of probiotics in aquaculture: An overview. Fish Shellfish Immunol. 2021, 117, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Sumon, T.A.; Hussain, M.A.; Sumon, M.A.A.; Jang, W.J.; Abellan, F.G.; Sharifuzzaman, S.M.; Brown, C.L.; Lee, E.-W.; Hasan, M.T. Functionality and prophylactic role of probiotics in shellfish aquaculture. Aquac. Rep. 2022, 25, 101220. [Google Scholar] [CrossRef]
- Puvanasundram, P.; Chong, C.M.; Sabri, S.; Yusoff, M.S.; Karim, M. Multi-strain probiotics: Functions, effectiveness and formulations for aquaculture applications. Aquac. Rep. 2021, 21, 100905. [Google Scholar] [CrossRef]
- Saoud, I.P.; Ghanawi, J.; Thompson, K.R.; Webster, C.D. A review of the culture and diseases of redclaw crayfish Cherax quadricarinatus (von Martens 1868). JWAS 2013, 44, 1–29. [Google Scholar] [CrossRef]
- Alvanou, M.V.; Apostolidis, A.P.; Lattos, A.; Michaelidis, B.; Giantsis, I.A. The Coding Mitogenome of the Freshwater Crayfish Pontastacus leptodactylus (Decapoda: Astacidea: Astacidae) from Lake Vegoritida, Greece and Its Taxonomic Classification. Genes 2023, 14, 494. [Google Scholar] [CrossRef]
- Chiu, C.H.; Guu, Y.K.; Liu, C.H.; Pan, T.M.; Cheng, W. Immune responses and gene expression in white shrimp, Litopenaeus vannamei, induced by Lactobacillus plantarum. Fish Shellfish Immunol. 2007, 23, 364–377. [Google Scholar] [CrossRef]
- Mohapatra, S.; Chakraborty, T.; Kumar, V.; Deboeck, G.; Mohanta, K.N. Aquaculture and stress management: A review of probiotic intervention. J. Anim. Physiol. Anim. Nutr. 2013, 97, 405–430. [Google Scholar] [CrossRef]
- Huynh, T.-G.; Shiu, Y.-L.; Nguyen, T.-P.; Truong, Q.-P.; Chen, J.-C.; Liu, C.-H. Current applications, selection, and possible mechanisms of actions of synbiotics in improving the growth and health status in aquaculture: A review. Fish Shellfish Immunol. 2017, 64, 367–382. [Google Scholar] [CrossRef]
- Sugita, H.; Hirose, Y.; Matsuo, N.; Deguchi, Y. Production of the antibacterial substance by Bacillus sp. strain NM 12, an intestinal bacterium of Japanese coastal fish. Aquaculture 1998, 165, 269–280. [Google Scholar] [CrossRef]
- Verschuere, L.; Rombaut, G.; Sorgeloos, P.; Verstraete, W. Probiotic bacteria as biological control agents in aquaculture. Microbiol. Mol. Biol. Rev. 2000, 64, 655–671. [Google Scholar] [CrossRef] [PubMed]
- Merrifield, D.L.; Dimitroglou, A.; Foey, A.; Davies, S.J.; Baker, R.T.M.; Bøgwald, J.; Castex, M.; Ringø, E. The current status and future focus of probiotic and prebiotic applications for salmonids. Aquaculture 2010, 302, 1–18. [Google Scholar] [CrossRef]
- Bikel, S.; Valdez-Lara, A.; Cornejo-Granados, F.; Rico, K.; Canizales-Quinteros, S.; Soberón, X.; Del Pozo-Yauner, L.; Ochoa-Leyva, A. Combining metagenomics, metatranscriptomics and viromics to explore novel microbial interactions: Towards a systems-level understanding of human microbiome. CSBJ 2015, 13, 390–401. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probiotic | Source | Species | Administration | Concentration and Duration | Responses | Refs. |
|---|---|---|---|---|---|---|
| Bacillus pumilus; B. licheniformis; B. subtilis; Acinetobacter genospecies 6, Acinetobacter grimontii and Chryseobacterium balustinum | Commercial Bacillus probiotics, and other three from red clawed larvae | Cherax quadricarinatus | Inhibition test | 104 CFU/mL | ∙ unable to outgrow and out-compete pathogenic bacteria A. hydrophila | [62] |
| L. plantarum | Intestine of rainbow trout | Astacus leptodactylus | Dietary | 107 (LB7), 108 (LB8), and 109 (LB9) CFUg−1 for 97 days | LB7 and LB8: ↑ TPP and PO LB8: ↑ LGC count ↑ LYZ activity LB7: ↑ SOD All LB groups: ↑ autochthonous LAB levels, lipase activity ↑ THC, SGC, and HC count LB8 and LB9: ↑digestive enzyme activity (protease, amylase, and ALP) LB7 and LB9: ↑ catalase (CAT) activity ∙ no significant growth ∙ no mortality after 24 h air exposure | [63] |
| A23 (B. amyloliquefaciens) | Intestine of healthy P. clarkii | Procambarus clarkii | Dietary | 1 × 107 CFU/g (A23–1) and 1 × 108 CFU/g (A23–2) for 28 days | ↑ intestinal digestive enzyme activities, innate immune enzyme activities ↑ white-spot syndrome virus (WSSV) resistance ↓ the abundance of Proteobacteria with 108 CFU/g supplement ↑ the abundance of Firmicutes and Bacteroides ↑ intestinal microbial diversity | [64] |
| B. coagulans (SCC-19) and L. lactis (Z-2) | SCC-19 from the gut of common carp and Z-2 from the gut of Cyprinus carpio L. | Procambarus clarkii | Dietary | 106, 107, and 108 CFU/g for 28 days | ↑ activities of immune-related enzymes in intestine ↑ mRNA expression of two AMP genes in intestinal ↑ intestinal integrity, thicker mucosal layer ↑ density granules in epithelial cells ↑ diversity of intestinal microbiota ↑ phagocytosis rate of hemocytes and pathogen A. hydrophila resistance | [65] |
| L. fermentum GR-3 | Chinese traditional fermented food (Jiangshui noodles) | Procambarus clarkii | Dietary | 1 × 108 CFU/g for 30 days | ↓As(III) concentration and residual level in hepatopancreas by 36% ↓ gut microbiota dysbiosis due to As(III) exposure ↓ As(III) accumulation in field ↑ aquaculture production | [66] |
| B. mycoides | Provided by Department of Agriculture and Food, Western Australia | Cherax cainii | Dietary | 108 CFU/g for 60 days | ↑ health and immune indices (protein and energy in tail muscle, THC in hemolymph) ↑ different microbial communities in hindgut ↑ cytokine genes expression associated with immunity and health status | [67] |
| AQ2 (Bacillus sp.); A10 (B. mycoides); A12 (Shewanella sp.); PM3 (B. subtilis); and PM4 (Bacillus sp.) | A10 and A12 from healthy farmed marron intestines; AQ2 from Aquasonic Pty. Ltd. New South Wales, Australia; PM3 and PM4 from Enviroplus Pty Ltd. Singapore | Cherax tenuinamus | Dietary | 108 CFU/g for 70 days | ∙ no significant impact on survival, growth, and intermoult period ∙ physiological condition in tail muscle indices, proportion of GC, and THC ∙ bacteria in hemolymph ∙ bacterial community in gut ∙ resistance towards V. mimicus | [68] |
| B. amyloliquefaciens | Zhejiang Science and Technology University, Zhejiang, China | Procambarus clarkii | Dietary | 4, 5, 6 g/kg for 7 days | ∙ immune-related genes expression ∙ immune parameters (THC, PO activity, and SOD activity) ∙ hemocyte apoptosis ∙ innate immunity regulation ∙ mortality rate towards challenge with WSSV | [69] |
| L. plantarum | Quest L. plantarum, Nutra Pharma, West Yorkshire, UK | Cherax cainii | Dietary | 1 × 109 CFU mL−1/kg for 56 days | ∙ hemolymph parameters and gut health ∙ expression of innate immune response genes ∙ diversity of gut microbiota | [70] |
| Spomune© (B. subtilis and C. butyricum) | Not mentioned | Cambarellus montezumae | Dietary | 1 × 107 CFU/g for 24 weeks | ∙ survival, growth, and weight gain | [71] |
| (Ecoterra®) composed of B. licheniformis, B. subtilis, Nitrobacter, Nitrosomonas, Rizobium, Saccharomyces cereviciae, and T. oxidans | Not mentioned | Cherax quadricarinatus | Water additive | 200,000 cells/liter for 60 days | ∙ no effect on growth, FCR, and survival ∙ mean value of total lipids in hemolymph, hemolymph glucose, and total lactate | [72] |
| S. cerevisiae | Intestinal tract of crayfish | Procambarus clarkii | Dietary | 107 CFU/g for 28 days | ∙ weight gain, SGR, expression of lysozyme and prophenolxidase ∙ abundance of Cetobacterium and Lactobacillus ∙ abundance of Citrobacter and Bacteroides ∙ resistance towards C. freundii | [73] |
| B. subtilis CK3 | Intestine of P. clarkii | Procambarus clarkii | Water additive | 1 × 105 CFU/mL for 4 weeks | ∙ antioxidant and immune-related enzymes and enzymes activities in hepatopancreas ∙ mortality ∙ immune response of P. clarkii towards A. veronii | [74] |
| Lactobacillus sp. | Digestive tract of angel fish Pterophyllum scalare | Cambarellus moctezumae | Dietary | 100 mL of Lactobacillus solution (La3) for 24 weeks | ∙ overall well-being ∙ final weight | [75] |
| Effective microorganisms’ serum with two major microorganisms as B. amyloliquefaciens spp. and L. plantarum | Rice-washed water | Astacus leptodactylus | Dietary powder and water additive | 1% and 5% of powder in diet and 0.01% serum in water for 60 days | ∙ no significant difference in growth performance ∙ severe pathological finding in both guts and hepatopancreas (inflammatory cell infiltrations in interstitial tissue, and lack of B, F, and R epithelial cells) ∙ survival rate | [76] |
| H. alvei | Hepatopancreas, gills, and intestine of adult crayfish and whole body of stage II crayfish juveniles and rearing water of adult and juvenile crayfish | Astacus leptodactylus | Diet and water additive | lactic acid bacteria (0.015 gL−1); H. alvei (106 CFU mL−1) and H. alvei added to water (106 CFU mL−1) | ∙ no significant impact on growth and survival | [77] |
| C. butyricum | Advanced Orthomolecular Research (AOR, Calgary, AB, Canada) | Cherax cainii | Dietary | 107 CFU/mL per kg for 42 days | ∙ moult number, growth rate, THC, LYZ activity in hemolymph and protein content of tail muscle ∙ diversity of bacterial community ∙ Clostridium abundance ∙ crayfish pathogen abundance (Vibrio and Aeromonas) ∙ expression level of immune-responsive gene towards challenge with V. mimicus | [78] |
| Subtilis-C (B. subtilis, B. licheniformis) | Not mentioned | Pontastacus leptodactylus | Dietary | 1.5 g per 1 kg of feed | ∙ immunity, survival rate ∙ ACC of lysosomal cationic protein in hemocytes | [79] |
| L. acidophilus and L. plantarum | Nature Way Probiotic (Warriewood, New South Wales, Australia) | Cherax cainii | Dietary | 109 CFU/mL per kg for 60 days | ∙ no significant differences in weight gain ↑ hemolymph parameters and biochemical composition of tail muscle, hepatopancreas health ↑ microvilli counts ↑ shift of beneficial microbial communities ↑ metabolic functions and genes associated with innate immune response | [60] |
| B. mycoides | Marron origin | Cherax cainii | Dietary | 108 CFU/g of feed for 10 weeks | ↑ survival at 48 h of transport ↑ intestinal bacterial population and THC ↑ hemolymph bacteria (bacteraemia) level | [80] |
| Prebiotic | Source | Species | Administration | Concentration and Duration | Responses | Refs. |
|---|---|---|---|---|---|---|
| Astaxanthin | H. pluvialis | Procambarus clarkii | Dietary | o.6% | ↑ WGR, SGR, and haemolymph immune-related enzyme activities ↑ MDA ↑ microbial dysbiosis and gut immune damage | [81] |
| Chlorella vulgaris | Chlorella vulgaris | Pontastacus leptodactylus | Dietary | 75% substitution for 63 days | ↑ final weight, SGR, PER, ADCOM and ADCCP ↑ alkaline protease, lipase, amylase, PO, SOD, LYZ, and NOS activity ↑ FCR dietary fishmeal substitution level (%) for maximum growth, SGR, and weight gain values | [82] |
| MOS and FOS | MOS, immunogen®, International Commerce Corporation Co., Waltham, MA, USA and FOS, Raftilose® P95, Orafti Co., Tienen, Belgium | Pontastacus leptodactylus | Dietary | 1.5, 3.0 and 4.5 g kg−1 in the single diets and 0.75, 1.5 and 2.25 g kg−1 in the combined diets for 126 days | ↑ SGR, VFI, survival rate and ↑ FCR values in 2.25 g kg−1 MOS and 1.5 g kg−1 FOS ↑ PER, LER, EER, PPV, LPV and EPV in 2.25 g kg−1 MOS and 1.5 g kg−1 FOS ↑ amylase, lipase, and alkaline protease activities and the mean of hemolymph indices in 2.25 g kg−1 MOS and 1.5 g kg−1 FOS ↑ activities of PO, SOD, LYZ, and NOS after 12-h air exposure challenge in combined diets | [83] |
| Prebiotic Vivinal-GOS® (rich in GOS) | Friesland Foods Domo Company (Zwolle, The Netherlands) | Pontastacus leptodactylus | Dietary | 0, 1, 2, and 3% GOS for 97 days | ↑ THC, SGC, and HC counts in 2% GOS diets ↑ CAT and CAT activity in 3% GOS diet ↑ LYZ, amylase and lipase activity, LAB levels, in 2% and 3% GOS-enriched diets ↑ THC, SGC, and HC count in 1% and 2% GOS diets ↑ total intestinal heterotrophic bacteria (TIHB) in the first 14 days in all GOS diets | [84] |
| PHB monomer (3-HB) | (166,898, Sigma Darmstadt, Germany) | Cherax quadricarinatus | Injection | 5 × 10 CFU/mL | ↑ phagocytosis, expression of microtubule-related genes ↑ growth of pathogenic bacteria | [85] |
| MOS (Bio-Mos®) | cell wall of S. cerevisiae | Cherax tenuinamus | Dietary | 0.2% and 0.4% Bio-Mos® for 30 days, 112 days for V. mimicus challenge, and 0.4% for 42 days for NH3 challenge | ↑ survival after bacterial infection and exposure to NH3 ↑ unaltered THC after bacterial infection ↑ THCs after exposure to NH3 ↑ unaltered Vibrio spp. in hemolymph after bacterial infection and exposure to NH3 ↑ Hemolymph clotting time in Bio-Mos® diet | [86] |
| β-Glucan | S. cerevisiae | Procambarus clarkii | Dietary | 0.025%, 0.05%, 0.1%, and 0.2% for 8 weeks | ↑ growth performance, antioxidant capacity, immunity, function and structure of the intestinal flora ↑ probiotics abundances of Hafnia, Acinetobacter ↑ probiotics abundance of Enterobacteriaceae ↑ Aeromonas abundance | [87] |
| MOS (Bio-Mos®) | Alltech | Cherax destructor | Dietary | 0.4% for 56 days | ↑ weight, SGR, and average weekly gain ↑ THC, GC, and SGC growth parameters ↑ protease activity in hepatopancreas ↑ amylase activity in the guts | [88] |
| Synbiotic | Source | Species | Administration | Concentration and Duration | Responses | Refs. |
|---|---|---|---|---|---|---|
| GOS+ Enterococcus faecalis | Enterococcus faecalis from gastrointestinal tract of aquatic species GOS from dairy products | Astacus leptodactylus | Dietary | 7.53 log CFU E. faecalis g−1 + 10 g kg−1 GOS for 126 days | ↑ SGR, VFI, survival rate ↑ FCR ↑ in vivo apparent digestibility coefficients ↑ ratios of presumptive autochthonous LAB to total viable aerobic heterotrophic bacteria ↑ PO, SOD, LYZ, and NOS activity ↑ mean survival rate towards A. hydrophila | [89] |
| prebiotics (MOS and XOS); probiotics (E. faecalis and P. acidilactici) and synbiotics | MOS from International Commerce Corporation Co., USA; XOS from Shandong Longlive Bio-Technology Co., China; E. faecalis from Nichi Nichi Pharmaceutical Co., Ltd., Japan; P. acidilactici (Bactocell®, Lallemand Inc., Montreal, QC, Canada) | Astacus leptodactylus | Dietary | 10 g kg−1 for prebiotics and 7.86 log CFU g−1 for probiotics for 126 days | XOS + E. faecalis: ↑ antibacterial activities in the shell mucus against Nocardia brasilience ↑ protein levels ↑ ALP and LYZ activities ↑ resistance after A. hydrophila injection ↑ growth rate and resistance to the A. hydrophila injection MOS + P. acidilactici: ↑ antibacterial activities in the shell mucus against Vibrio harveyi Both synbiotic diets: ↑ ratio of the Lactobacillus count to the total viable count | [90] |
| Biogen® (B. licheniformis and B. subtilis) + sodium alginate | cell walls of brown seaweed | Procambarus clarkii | Dietary | 1%, 2%, 3% Biogen® and (3 g/L) of sodium alginate | ↑ survival, wet weight, SGR, hemocyte count. and proPO activity | [91] |
| L. salivarius (LS) ATCC 11741 + PE | PE from Sigma-Aldrich Inc; L. salivarius from the Iranian Biological Resource Center | Postantacus leptodactylus | Dietary | LS1 (1 × 107 CFU/g), LS2 (1 × 109 CFU/g), PE1 (5 g/kg), PE2 (10 g/kg), LS1PE1 (1 × 107 CFU/g + 5 g/kg); LS2PE2 (1 × 109 CFU/g +10 g/kg) for 18 weeks | In all diets: ↑ final weight, weight gain, SGR and FCR ↑ TVC and LAB ↑ resistance towards A. hydrophila LS1PE1 and LS2PE2: ↑ amylase and protease enzymes activity ↑ GPx and SOD activity ↑ MDA content LS1PE1: ↑ THC, LGC, SGC, and HC count ↑ LYZ, PO, NOS, and AKP activity | [59] |
| Poultry by-product fermented by L. casei and S.cerevisiae | Poultry by-product from Specialty Feeds Pty. Ltd., Western Australia; L. casei and S. cerevisiae from Baker’s yeast | Cherax cainii | Dietary | 75% substitution for 70 days | ↑ no significant difference in final weight ↑ intestinal microvilli number ↑ Lactobacillus and Streptococcus in the intestine ↑ Aeromonas number in the intestine ↑ cytokines expression ↑ LYZ and phagocytic activity ↑ survival towards challenge with V. mimicus | [92] |
| Lactobacillus sp. and coconut powder | Coconut powder from coconut pulp from agricultural wastes; Lactobacillus sp. not mentioned | Cherax sp. | Dietary | Lactobacillus 2%/kg of feed + coconut powder 2%/kg feed | ↑ growth rate ↑ no significant differences in survival rate | [93] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvanou, M.V.; Feidantsis, K.; Staikou, A.; Apostolidis, A.P.; Michaelidis, B.; Giantsis, I.A. Probiotics, Prebiotics, and Synbiotics Utilization in Crayfish Aquaculture and Factors Affecting Gut Microbiota. Microorganisms 2023, 11, 1232. https://doi.org/10.3390/microorganisms11051232
Alvanou MV, Feidantsis K, Staikou A, Apostolidis AP, Michaelidis B, Giantsis IA. Probiotics, Prebiotics, and Synbiotics Utilization in Crayfish Aquaculture and Factors Affecting Gut Microbiota. Microorganisms. 2023; 11(5):1232. https://doi.org/10.3390/microorganisms11051232
Chicago/Turabian StyleAlvanou, Maria V., Konstantinos Feidantsis, Alexandra Staikou, Apostolos P. Apostolidis, Basile Michaelidis, and Ioannis A. Giantsis. 2023. "Probiotics, Prebiotics, and Synbiotics Utilization in Crayfish Aquaculture and Factors Affecting Gut Microbiota" Microorganisms 11, no. 5: 1232. https://doi.org/10.3390/microorganisms11051232