

Improving Rhamnolipids Biosynthesis in Pseudomonas sp. L01 through Atmospheric and Room-Temperature Plasma (ARTP) Mutagenesis
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Culture Conditions
2.2. Mutagenesis by ARTP
2.3. Preliminary Screening of Biosurfactants High-Yield Mutants
2.4. Crude Biosurfactant Extraction
2.5. Evaluation of Genetic Stability of the Mutants
2.6. Surface Tension Measurement
2.7. Genome Sequencing and Comparative Genomic Analysis of the Strain L01 and the Mutants
2.8. Statistical Analysis
3. Results
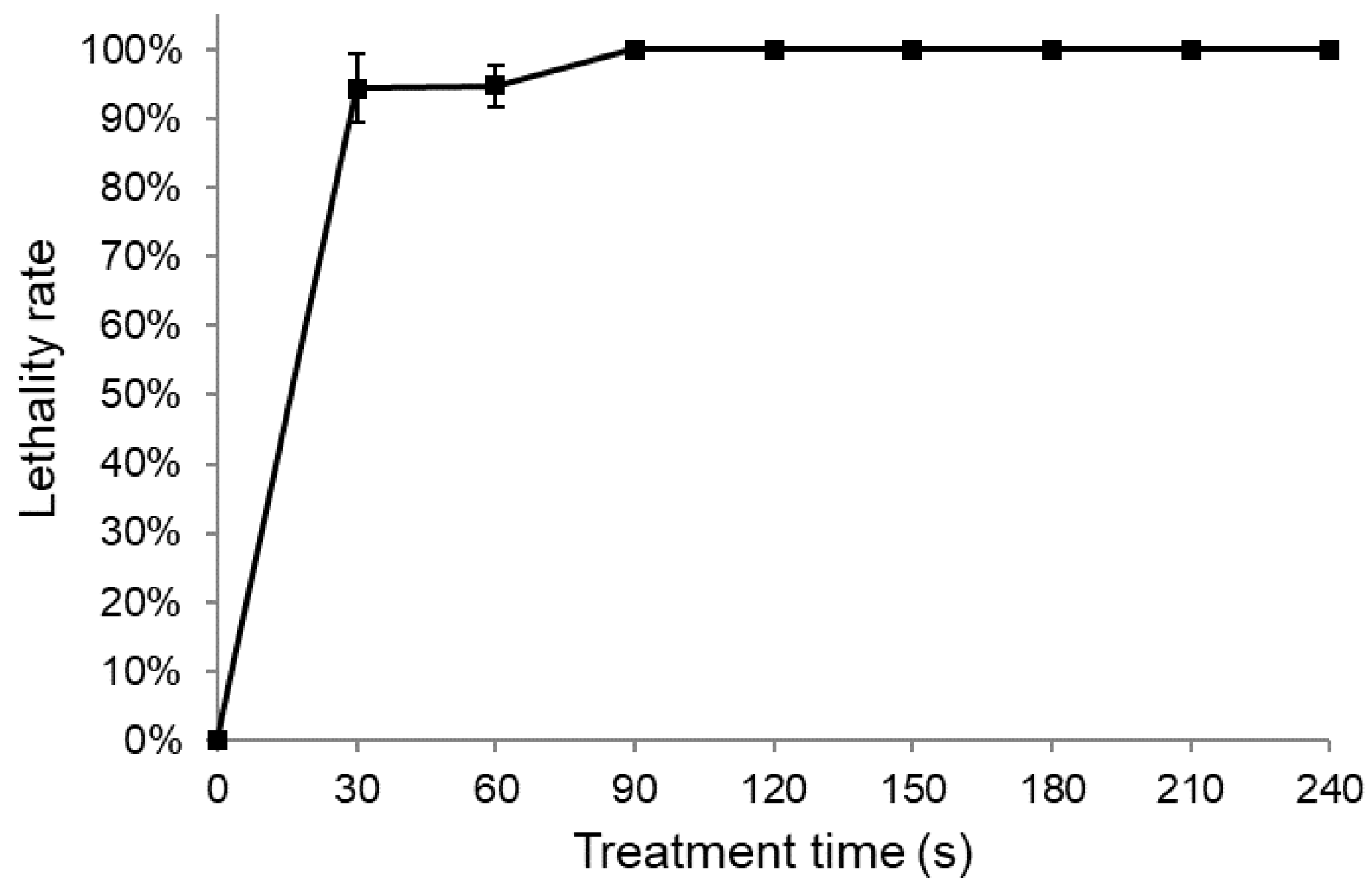
3.1. Lethality Rate of Pseudomonas sp. L01 with ARTP Treatment
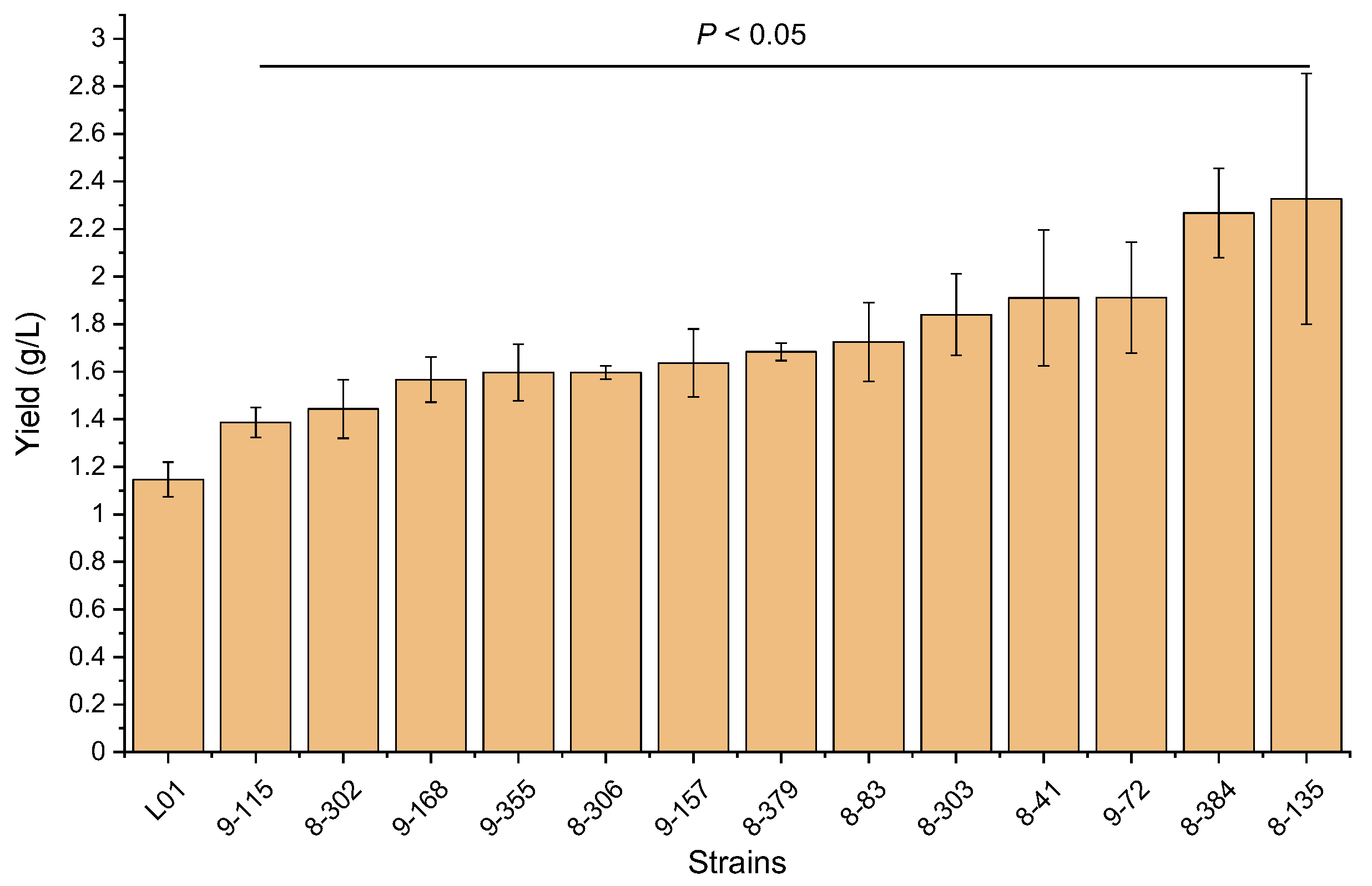
3.2. Screening and Verification of the Mutants with High-Yield Rhamnolipids Production
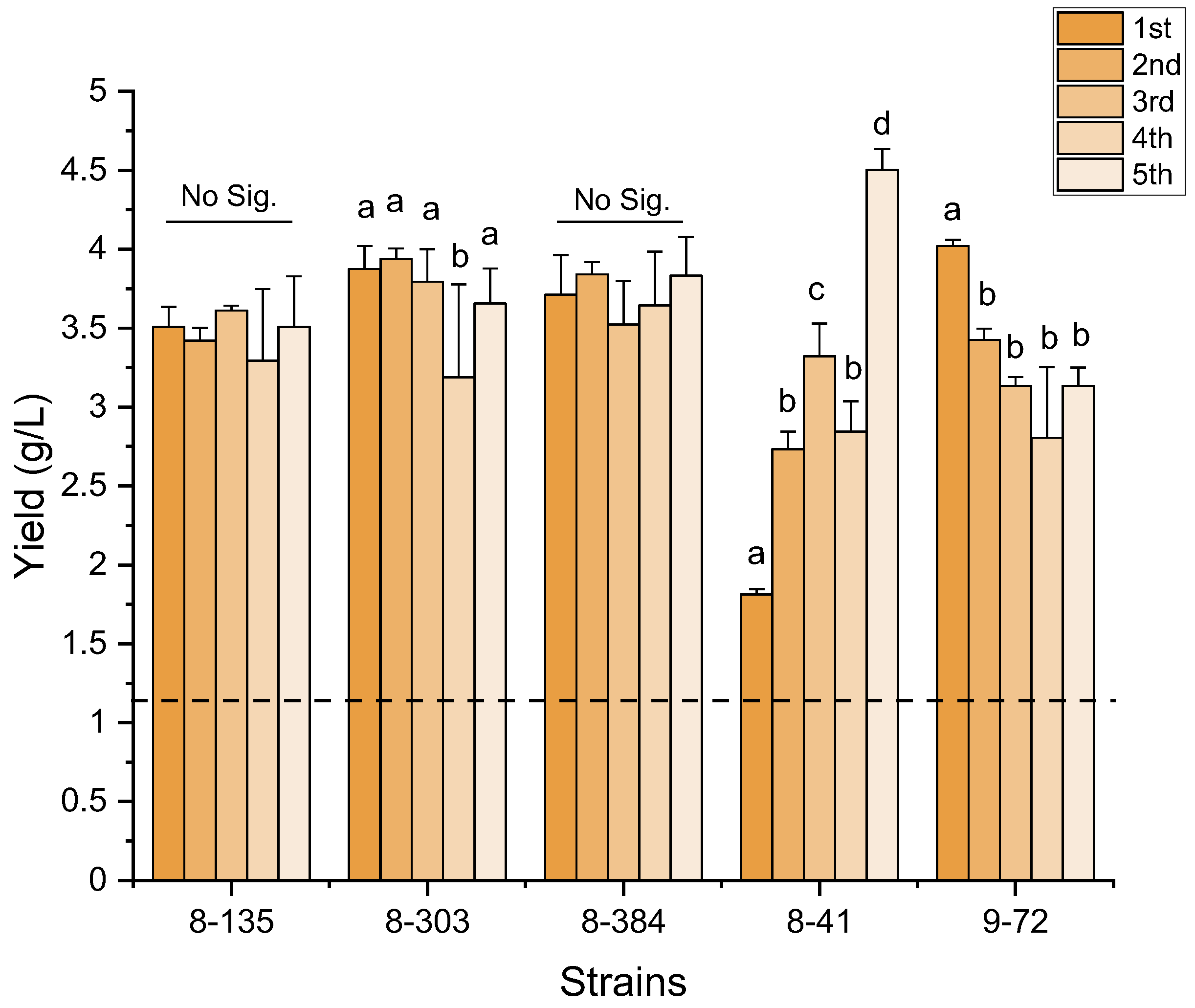
3.3. The Genetic Stability of the Mutants
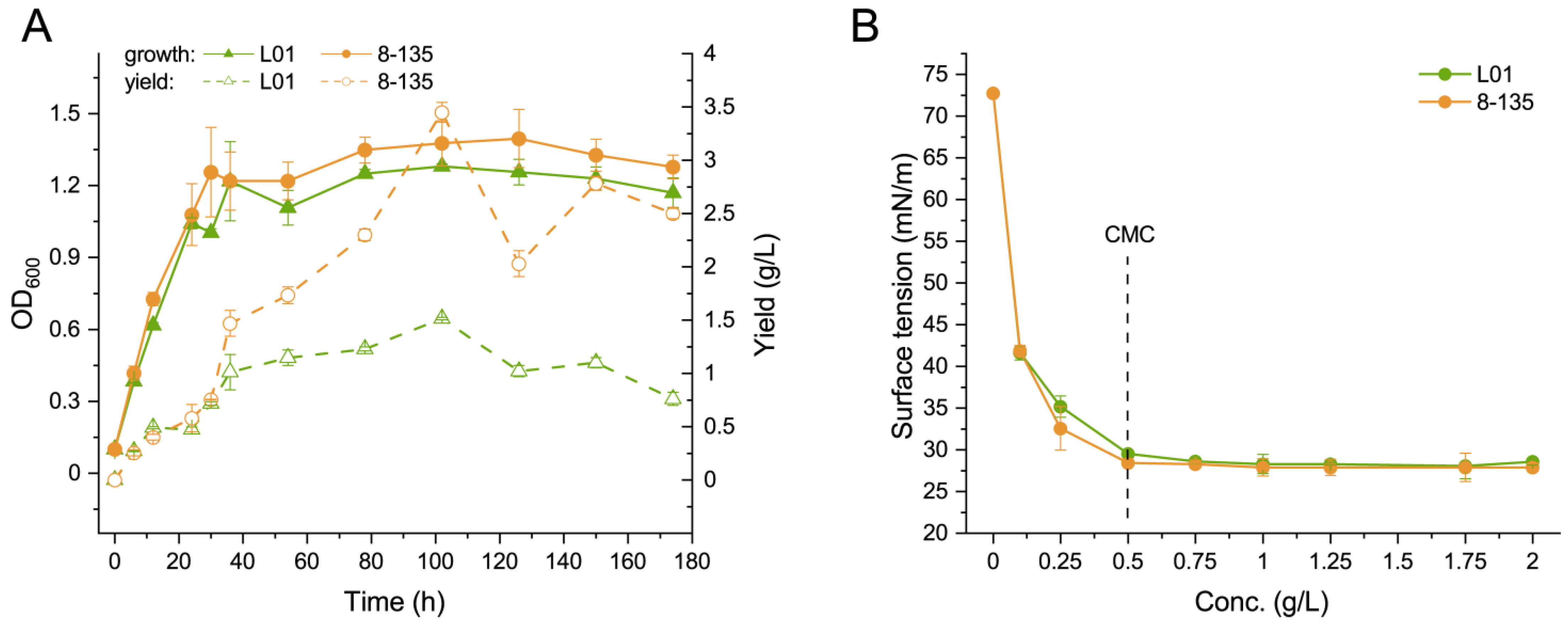
3.4. Comparison of the Growth and Biosurfactant Production between the Wild-Type Strain and 8-135 Strain
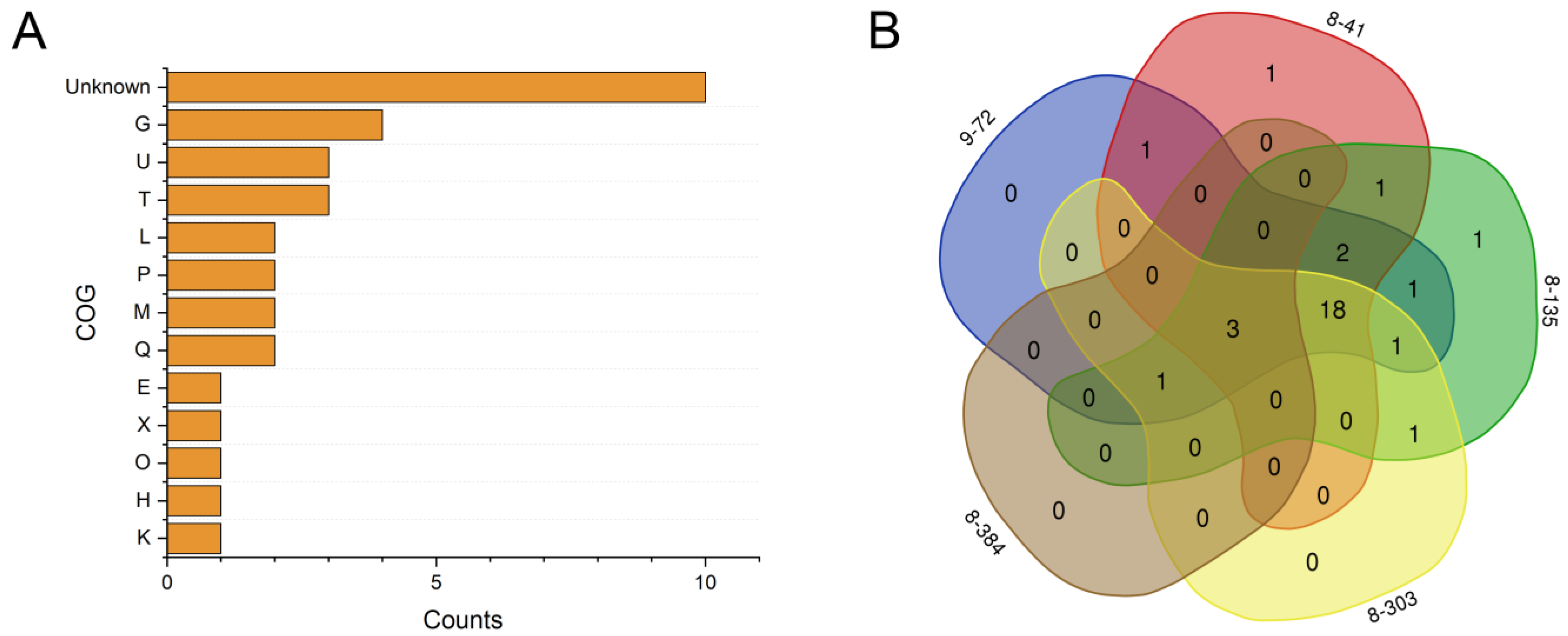
3.5. Comparative Genomic Analysis of the Wild-Type Strain and the Mutants
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geys, R.; Soetaert, W.; Van Bogaert, I. Biotechnological opportunities in biosurfactant production. Curr. Opin. Biotechnol. 2014, 30, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Nitschke, M.; Costa, S.G.V.A.O. Biosurfactants in food industry. Trends Food Sci. Technol. 2007, 18, 252–259. [Google Scholar] [CrossRef]
- Ron, E.Z.; Rosenberg, E. Biosurfactants and oil bioremediation. Curr. Opin. Biotech. 2002, 13, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Souza, E.C.; Vessoni-Penna, T.C.; Oliveira, R.P.D. Biosurfactant-enhanced hydrocarbon bioremediation: An overview. Int. Biodeterior. Biodegrad. 2014, 89, 88–94. [Google Scholar] [CrossRef]
- Desai, J.D.; Banat, I.M. Microbial production of surfactants and their commercial potential. Microbiol. Mol. Biol. Rev. 1997, 61, 47–64. [Google Scholar] [CrossRef]
- Chafale, A.; Kapley, A. Biosurfactants as microbial bioactive compounds in microbial enhanced oil recovery. J. Biotechnol. 2022, 352, 1–15. [Google Scholar] [CrossRef]
- Liu, G.S.; Zhong, H.; Yang, X.; Liu, Y.; Shao, B.B.; Liu, Z.F. Advances in applications of rhamnolipids biosurfactant in environmental remediation: A review. Biotechnol. Bioeng. 2018, 115, 796–814. [Google Scholar] [CrossRef]
- Mgbechidinma, C.L.; Akan, O.D.; Zhang, C.; Huang, M.; Linus, N.; Zhu, H.; Wakil, S.M. Integration of green economy concepts for sustainable biosurfactant production—A review. Bioresour. Technol. 2022, 364, 128021. [Google Scholar] [CrossRef]
- Sun, W.Y.; Zhu, B.K.; Yang, F.; Dai, M.; Sehar, S.; Peng, C.S.; Ali, I.; Naz, I. Optimization of biosurfactant production from Pseudomonas sp. CQ2 and its application for remediation of heavy metal contaminated soil. Chemosphere 2021, 265, 129090. [Google Scholar] [CrossRef]
- Xu, Y.; Jing, Y.L.; Zhang, Y.Q.; Liu, Q.J.; Xiu, J.L.; Zhang, K.; Jia, N.H.; Zhou, M.H.; Zhou, X.Y.; Huang, J.; et al. Single-cell-based high-throughput cultivation and functional characterization of biosurfactant-producing bacteria from soil and oilfield-produced water. Microorganisms 2022, 10, 2216. [Google Scholar] [CrossRef]
- Eslami, P.; Hajfarajollah, H.; Bazsefidpar, S. Recent advancements in the production of rhamnolipid biosurfactants byPseudomonas aeruginosa. RSC Adv. 2020, 10, 34014–34032. [Google Scholar] [CrossRef] [PubMed]
- Chong, H.Q.; Li, Q.X. Microbial production of rhamnolipids: Opportunities, challenges and strategies. Microb. Cell Fact. 2017, 16, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashif, A.; Rehman, R.; Fuwad, A.; Shahid, M.K.; Dayarathne, H.N.P.; Jamal, A.; Aftab, M.N.; Mainali, B.; Choi, Y. Current advances in the classification, production, properties and applications of microbial biosurfactants—A critical review. Adv. Colloid Interface Sci. 2022, 306, 102718. [Google Scholar] [CrossRef] [PubMed]
- Varjani, S.J.; Upasani, V.N. Critical review on biosurfactant analysis, purification and characterization using rhamnolipid as a model biosurfactant. Bioresour. Technol. 2017, 232, 389–397. [Google Scholar] [CrossRef]
- Randhawa, K.K.S.; Rahman, P.K.S.M. Rhamnolipid biosurfactants—Past, present, and future scenario of global market. Front. Microbiol. 2014, 5, 454. [Google Scholar] [CrossRef] [Green Version]
- Dobler, L.; Vilela, L.F.; Almeida, R.V.; Neves, B.C. Rhamnolipids in perspective: Gene regulatory pathways, metabolic engineering, production and technological forecasting. New Biotechnol. 2016, 33, 123–135. [Google Scholar] [CrossRef]
- Gutierrez-Gomez, U.; Soto-Aceves, M.P.; Servin-Gonzalez, L.; Soberon-Chavez, G. Overproduction of rhamnolipids in Pseudomonas aeruginosa PA14 by redirection of the carbon flux from polyhydroxyalkanoate synthesis and overexpression of the rhlAB-R operon. Biotechnol. Lett. 2018, 40, 1561–1566. [Google Scholar] [CrossRef]
- Beuker, J.; Barth, T.; Steier, A.; Wittgens, A.; Rosenau, F.; Henkel, M.; Hausmann, R. High titer heterologous rhamnolipid production. AMB Express 2016, 6, 124. [Google Scholar] [CrossRef] [Green Version]
- Kahraman, H.; Erenler, S.O. Rhamnolipid production by Pseudomonas aeruginosa engineered with the Vitreoscilla hemoglobin gene. Appl. Biochem. Microbiol. 2012, 48, 188–193. [Google Scholar] [CrossRef]
- Snow, A.A.; Andow, D.A.; Gepts, P.; Hallerman, E.M.; Power, A.; Tiedje, J.M.; Wolfenbarger, L.L. Genetically engineered organisms and the environment: Current status and recommendations. Ecol. Appl. 2005, 15, 377–404. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.M.; Zhang, J.; Ji, X.M.; Fang, Z.; Wu, Z.M.; Chen, J.; Du, G.C. Evolutionary engineering of industrial microorganisms-strategies and applications. Appl. Microbiol. Biotechnol. 2018, 102, 4615–4627. [Google Scholar] [CrossRef] [PubMed]
- Raza, Z.A.; Rehman, A.; Khan, M.S.; Khalid, Z.M. Improved production of biosurfactant by a Pseudomonas aeruginosa mutant using vegetable oil refinery wastes. Biodegradation 2007, 18, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Lotfabad, T.B.; Abassi, H.; Ahmadkhaniha, R.; Roostaazad, R.; Masoomi, F.; Zahiri, H.S.; Ahmadian, G.; Vali, H.; Noghabi, K.A. Structural characterization of a rhamnolipid-type biosurfactant produced by Pseudomonas aeruginosa MR01: Enhancement of di-rhamnolipid proportion using gamma irradiation. Colloid Surf. B-Biointerfaces 2010, 81, 397–405. [Google Scholar] [CrossRef]
- Raza, Z.A.; Khan, M.S.; Khalid, Z.M. Evaluation of distant carbon sources in biosurfactant production by a gamma ray-induced Pseudomonas putida mutant. Process Biochem. 2007, 42, 686–692. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, X.F.; Li, H.P.; Wang, L.Y.; Zhang, C.; Xing, X.H.; Bao, C.Y. Atmospheric and room temperature plasma (ARTP) as a new powerful mutagenesis tool. Appl. Microbiol. Biotechnol. 2014, 98, 5387–5396. [Google Scholar] [CrossRef] [PubMed]
- Ottenheim, C.; Nawrath, M.; Wu, J.C. Microbial mutagenesis by atmospheric and room-temperature plasma (ARTP): The latest development. Bioresour. Bioprocess. 2018, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.X.; Wu, S.H.; Wang, H.Y.; Dong, Y.Z.; Li, X.L.; Wang, S.J.; Fan, H.N.; Zhuang, X.L. Optimization of conditions for a surfactant-producing strain and application to petroleum hydrocarbon-contaminated soil bioremediation. Colloid Surf. B-Biointerfaces 2022, 213, 112428. [Google Scholar] [CrossRef]
- Ma, X.J.; Zhang, H.M.; Lu, X.F.; Han, J.; Zhu, H.X.; Wang, H.; Yao, R.S. Mutant breeding of Starmerella bombicola by atmospheric and room-temperature plasma (ARTP) for improved production of specific or total sophorolipids. Bioprocess. Biosyst. Eng. 2020, 43, 1869–1883. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Delcher, A.L.; Harmon, D.; Kasif, S.; White, O.; Salzberg, S.L. Improved microbial gene identification with GLIMMER. Nucleic Acids Res. 1999, 27, 4636–4641. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Proc, G.P.D. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Hua, X.F.; Wang, J.; Wu, Z.J.; Zhang, H.X.; Li, H.P.; Xing, X.H.; Liu, Z. A salt tolerant Enterobacter cloacae mutant for bioaugmentation of petroleum- and salt-contaminated soil. Biochem. Eng. J. 2010, 49, 201–206. [Google Scholar] [CrossRef]
- Dong, T.T.; Gong, J.S.; Gu, B.C.; Zhang, Q.; Li, H.; Lu, Z.M.; Lu, M.L.; Shi, J.S.; Xu, Z.H. Significantly enhanced substrate tolerance of Pseudomonas putida nitrilase via atmospheric and room temperature plasma and cell immobilization. Bioresour. Technol. 2017, 244, 1104–1110. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Xin, K.; Zhang, L.; Zhou, J.; Xu, A.; Dong, W.; Jiang, M. Integration of ARTP mutation and adaptive laboratory evolution to reveal 1,4-butanediol degradation in Pseudomonas putida KT2440. Microbiol. Spectr. 2023, e04988-22. [Google Scholar] [CrossRef] [PubMed]
- Morgan, W.F.; Day, J.P.; Kaplan, M.I.; McGhee, E.M.; Limoli, C.L. Genomic instability induced by ionizing radiation. Radiat. Res. 1996, 146, 247–258. [Google Scholar] [CrossRef] [PubMed]
- He, C.Q.; Dong, W.; Li, J.; Li, Y.P.; Huang, C.; Ma, Y.L. Characterization of rhamnolipid biosurfactants produced by recombinant Pseudomonas aeruginosa strain DAB with removal of crude oil. Biotechnol. Lett. 2017, 39, 1381–1388. [Google Scholar] [CrossRef]
- Zhao, F.; Yuan, M.L.; Lei, L.Y.; Li, C.Y.; Xu, X.M. Enhanced production of mono-rhamnolipid in Pseudomonas aeruginosa and application potential in agriculture and petroleum industry. Bioresour. Technol. 2021, 323, 124605. [Google Scholar] [CrossRef]
- Wang, Q.H.; Fang, X.D.; Bai, B.J.; Liang, X.L.; Shuler, P.J.; Goddard, W.A.; Tang, Y.C. Engineering bacteria for production of rhamnolipid as an agent for enhanced oil recovery. Biotechnol. Bioeng. 2007, 98, 842–853. [Google Scholar] [CrossRef]
- Collins, T.; Margesin, R. Psychrophilic lifestyles: Mechanisms of adaptation and biotechnological tools. Appl. Microbiol. Biotechnol. 2019, 103, 2857–2871. [Google Scholar] [CrossRef] [PubMed]
- Gesheva, V.; Stackebrandt, E.; Vasileva-Tonkova, E. Biosurfactant production by halotolerant Rhodococcus fascians from Casey Station, Wilkes Land, Antarctica. Curr. Microbiol. 2010, 61, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Perfumo, A.; Banat, I.M.; Marchant, R. Going green and cold: Biosurfactants from low-temperature environments to biotechnology applications. Trends Biotechnol. 2018, 36, 277–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowell, D.N.; Anderson, M.S.; Raetz, C.R.H. Molecular-cloning of the genes for lipid-a disaccharide synthase and udp-n-acetylglucosamine acyltransferase in Escherichia-coli. J. Bacteriol. 1986, 168, 152–159. [Google Scholar] [CrossRef] [Green Version]
- King, J.D.; Kocincova, D.; Westman, E.L.; Lam, J.S. Lipopolysaccharide biosynthesis in Pseudomonas aeruginosa. Innate Immun. 2009, 15, 261–312. [Google Scholar] [CrossRef]
- Soberon-Chavez, G.; Lepine, F.; Deziel, E. Production of rhamnolipids by Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2005, 68, 718–725. [Google Scholar] [CrossRef]
- Magri, M.; Abdel-Mawgoud, A.M. Identification of putative producers of rhamnolipids/glycolipids and their transporters using genome mining. Curr. Res. Biotechnol. 2022, 4, 152–166. [Google Scholar] [CrossRef]
- Rahim, R.; Ochsner, U.A.; Olvera, C.; Graninger, M.; Messner, P.; Lam, J.S.; Soberon-Chavez, G. Cloning and functional characterization of the Pseudomonas aeruginosa rhlC gene that encodes rhamnosyltransferase 2, an enzyme responsible for di-rhamnolipid biosynthesis. Mol. Microbiol. 2001, 40, 708–718. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Ti 1 | Tv 2 | Ti/Tv | nsSNPs 3 | sSNPs 4 | ncSNPs 5 |
|---|---|---|---|---|---|---|
| 8-135 | 81 | 70 | 1.16 | 35 | 30 | 86 |
| 8-303 | 79 | 60 | 1.32 | 29 | 31 | 79 |
| 8-384 | 32 | 10 | 3.2 | 6 | 21 | 25 |
| 8-41 | 85 | 64 | 1.33 | 29 | 35 | 85 |
| 9-72 | 86 | 71 | 1.21 | 33 | 31 | 93 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Jing, Y.; Zhang, Q.; Xiu, J.; Tian, M.; Cui, Q.; Ma, Y.; Yi, L.; Han, L.; Qian, Y.; et al. Improving Rhamnolipids Biosynthesis in Pseudomonas sp. L01 through Atmospheric and Room-Temperature Plasma (ARTP) Mutagenesis. Microorganisms 2023, 11, 1182. https://doi.org/10.3390/microorganisms11051182
Xu Y, Jing Y, Zhang Q, Xiu J, Tian M, Cui Q, Ma Y, Yi L, Han L, Qian Y, et al. Improving Rhamnolipids Biosynthesis in Pseudomonas sp. L01 through Atmospheric and Room-Temperature Plasma (ARTP) Mutagenesis. Microorganisms. 2023; 11(5):1182. https://doi.org/10.3390/microorganisms11051182
Chicago/Turabian StyleXu, Ying, Yali Jing, Qun Zhang, Jianlong Xiu, Maozhang Tian, Qingfeng Cui, Yuandong Ma, Lina Yi, Lu Han, Yuchen Qian, and et al. 2023. "Improving Rhamnolipids Biosynthesis in Pseudomonas sp. L01 through Atmospheric and Room-Temperature Plasma (ARTP) Mutagenesis" Microorganisms 11, no. 5: 1182. https://doi.org/10.3390/microorganisms11051182