

Plastoquinone Lipids: Their Synthesis via a Bifunctional Gene and Physiological Function in a Euryhaline Cyanobacterium, Synechococcus sp. PCC 7002


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cyanobacterial Strains, Media, and Growth Conditions
2.2. Preparation of Lipids and Analysis of Their Constituent Fatty Acids
2.3. MS Spectrometric Analysis
2.4. Gene Manipulation in Synechococcus 7002
3. Results
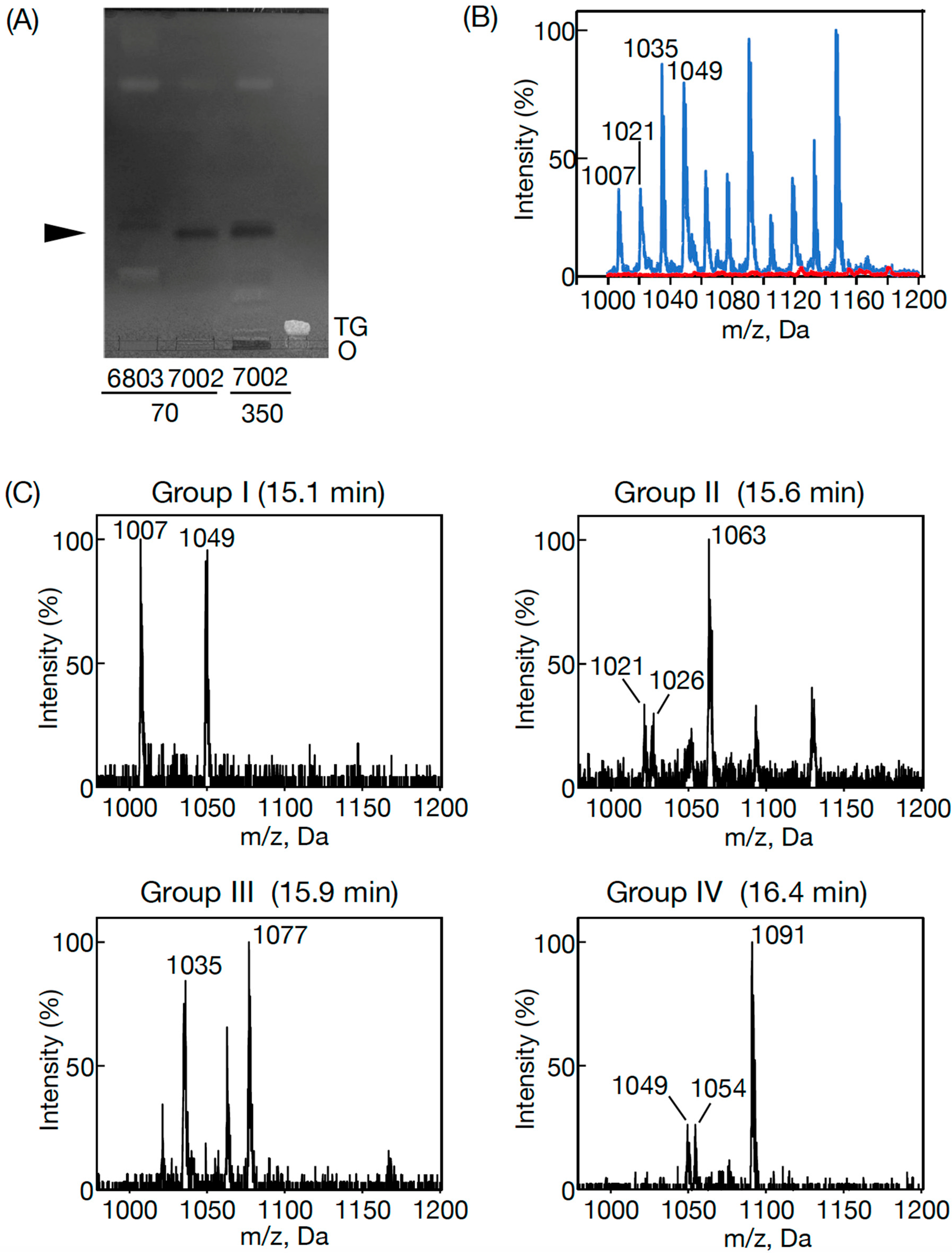
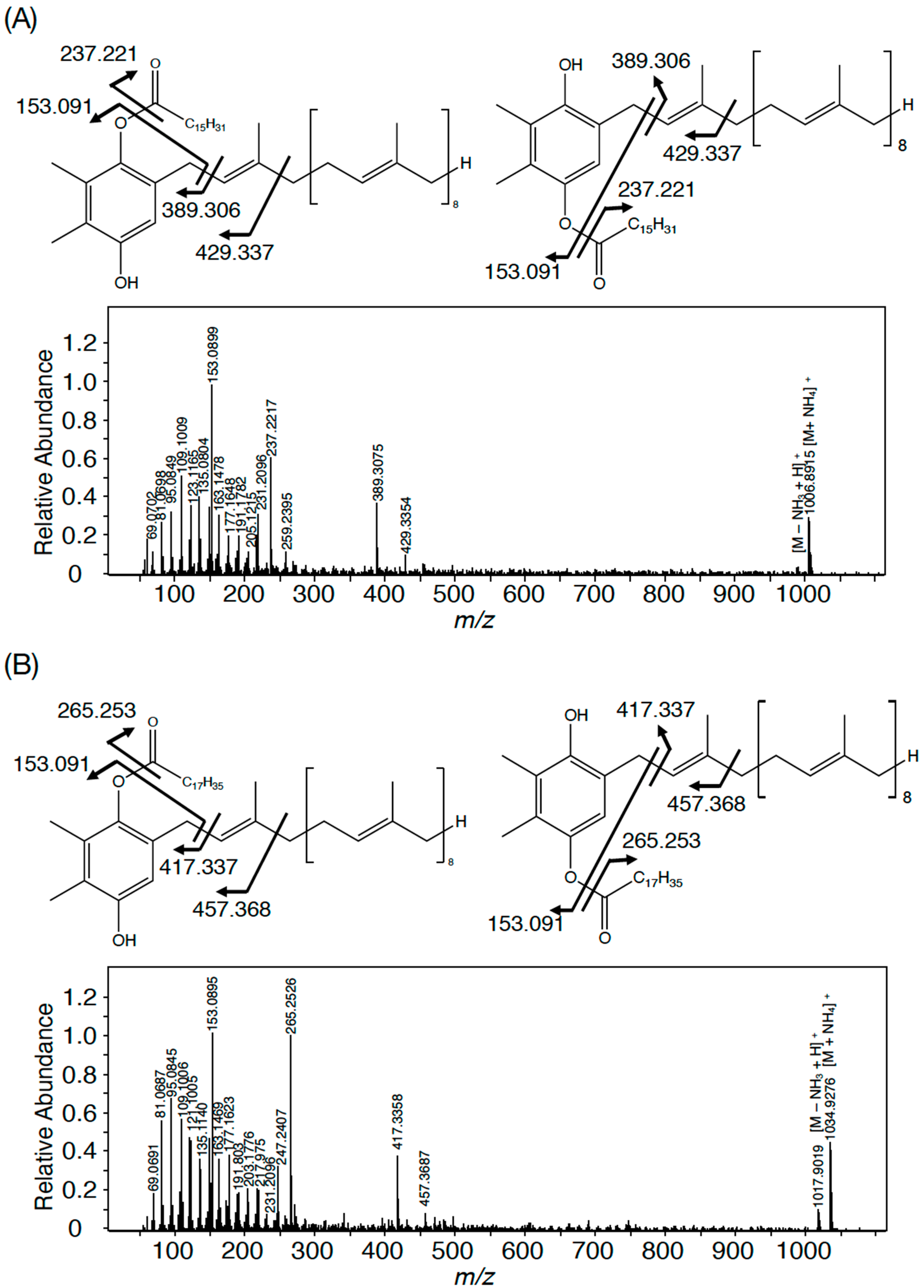
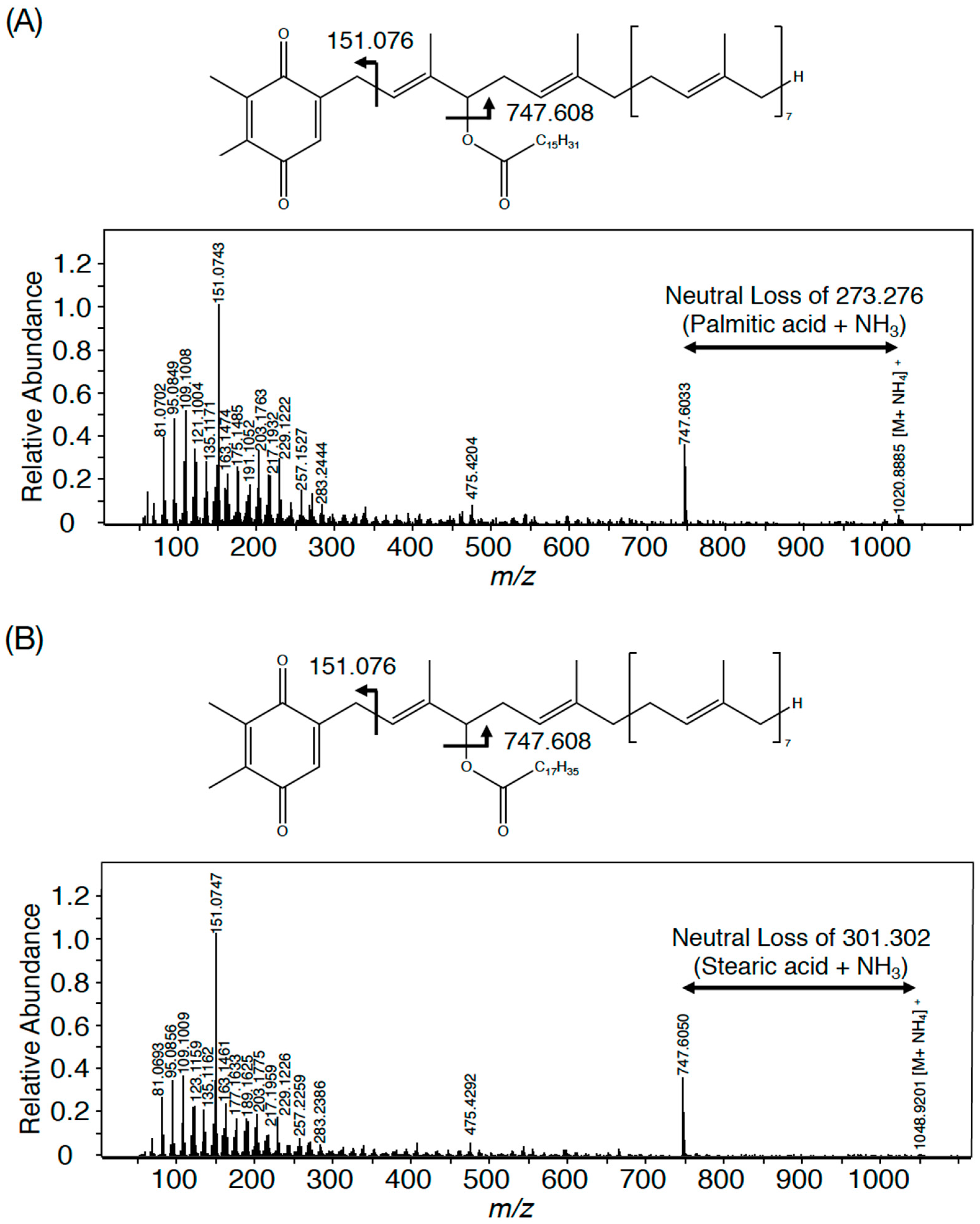
3.1. The Presence of PQ Lipids in Synechococcus 7002
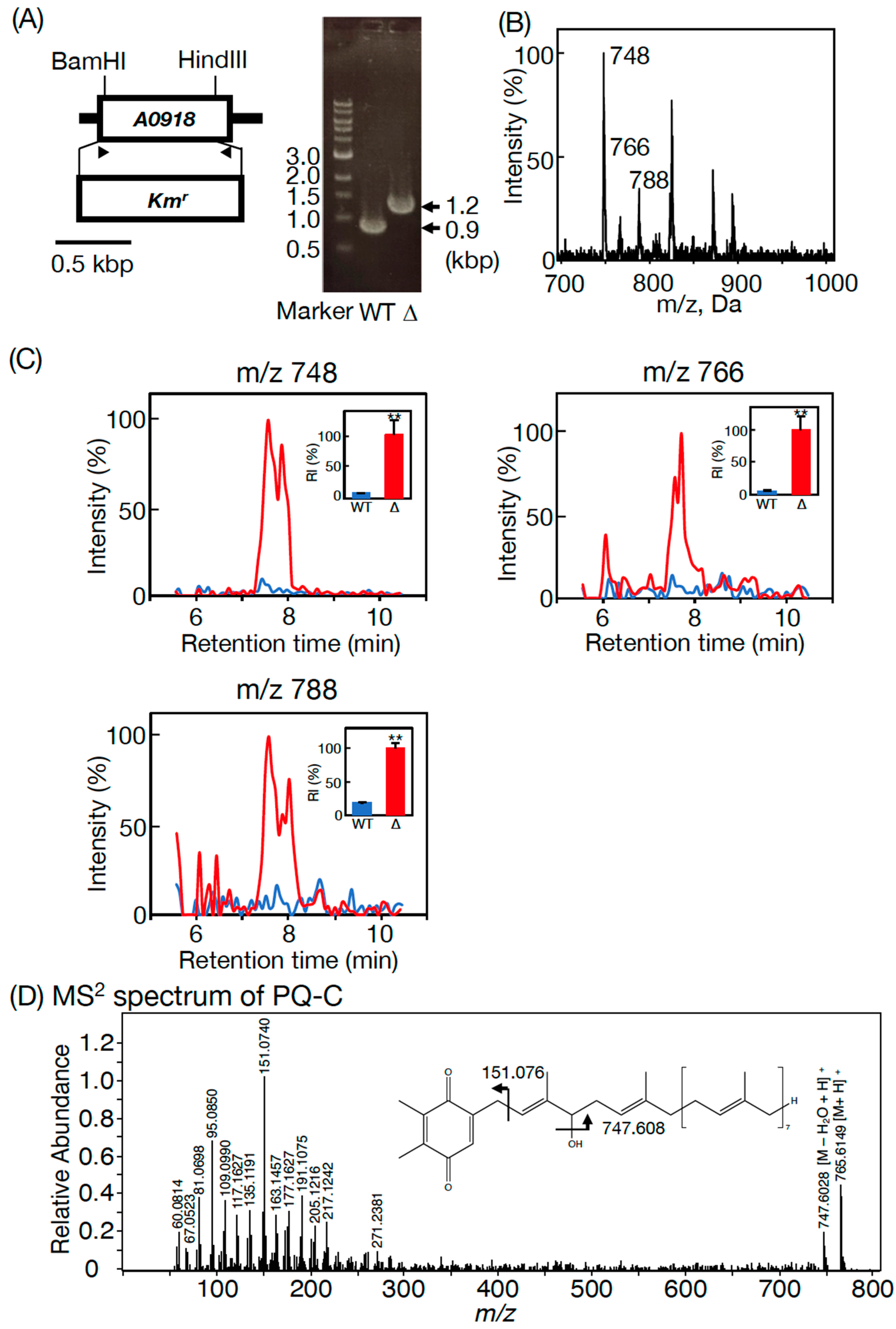
3.2. Essentiality of SYNPCC7002-A0918, an slr2103 Homolog, for PQ Lipid Synthesis in Synechococcus 7002
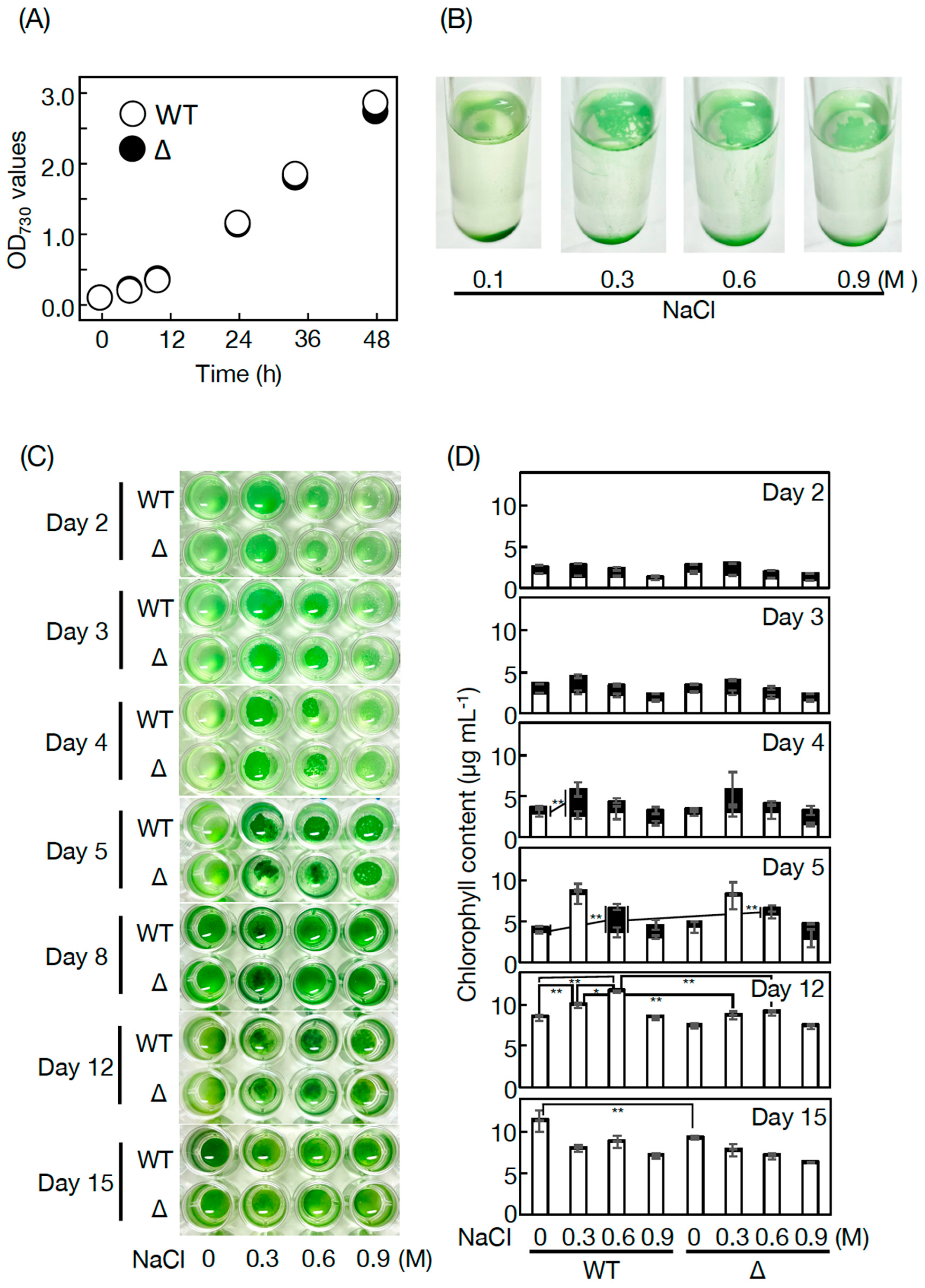
3.3. Physiological Roles of PQ Lipids in Synechococcus 7002
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kruk, J.; Szymańska, R. Singlet oxygen oxidation products of carotenoids, fatty acids and phenolic prenyllipids. J. Photochem. Photobiol. B 2021, 216, 112148. [Google Scholar] [CrossRef] [PubMed]
- Mori-Moriyama, N.; Yoshitomi, T.; Sato, N. Acyl PQH2 is a major cyanobacterial substance that co-migrates with triacylglycerol in thin-layer chromatography. Biochem. Biophys. Res. Commun. 2023, 641, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Aoki, M.; Hirai, K.; Sagam, T.; Ito, R.; Tsuzuki, M.; Sato, N. slr2103, a homolog of type-2 diacylglycerol acyltransferase genes, for plastoquinone-related neutral lipid synthesis and NaCl-stress acclimatization in a cyanobacterium Synechocystis sp. PCC 6803. Front. Plant Sci. 2023, 14, 1181180. [Google Scholar] [CrossRef]
- Dłużewska, J.; Zieliński, K.; Nowicka, B.; Szymańska, R.; Kruk, J. New prenyllipid metabolites identified in Arabidopsis during photo-oxidative stress. Plant Cell Environ. 2015, 38, 2698–2706. [Google Scholar] [CrossRef]
- Kruk, J.; Burda, K.; Schmid, G.H.; Radunz, A.; Strzałka, K. Function of plastoquinones B and C as electron acceptors in Photosystem II and fatty acid analysis of plastoquinone B. Photosynth. Res. 1998, 58, 203–209. [Google Scholar] [CrossRef]
- Goncalves, E.C.; Wilkie, A.C.; Kirst, M.; Rathinasabapathi, B. Metabolic regulation of triacylglycerol accumulation in the green algae: Identification of potential targets for engineering to improve oil yield. Plant Biotech. J. 2016, 14, 1649–1660. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef] [PubMed]
- Aizouq, M.; Peisker, H.; Gutbrod, K.; Melzer, M.; Hölzl, G.; Dörmann, P. Triacylglycerol and phytyl ester synthesis in Synechocystis sp. PCC6803. Proc. Natl. Acad. Sci. USA 2020, 117, 6216–6222. [Google Scholar] [CrossRef] [PubMed]
- Peramuna, A.; Summers, M.L. Composition and occurrence of lipid droplets in the cyanobacterium Nostoc Punctiforme. Arch. Microbiol. 2014, 196, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Piorreck, M.; Baasch, K.-H.; Pohl, P. Biomass production, total protein, chlorophyll, lipids and fatty acids of freshwater green algae and blue-green algae under different nitrogen regimes. Phytochemistry 1984, 23, 207–216. [Google Scholar] [CrossRef]
- Santana-Sánchez, A.; Lynch, F.; Sirin, S.; Allahverdiyeva, Y. Nordic cyanobacterial and algal lipids: Triacylglycerol accumulation, chemotaxonomy and bioindustrial potential. Physiol. Plant. 2021, 173, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Ishikawa, T.; Tamura, S.; Saito, Y.; Kawai-Yamada, M.; Hihara, Y. Quantitative and qualitative analyses of triacylglycerol production in the wild-type cyanobacterium Synechocystis sp. PCC 6803 and the strain expressing atfA from Acinetobacter baylyi ADP1. Plant Cell Physiol. 2020, 61, 1537–1547. [Google Scholar] [CrossRef] [PubMed]
- Taranto, P.A.; Keenan, T.W.; Potts, M. Rehydration induces rapid onset of lipid biosynthesis in desiccated Nostoc commune (Cyanobacteria). Biochem. Biophys. Acta 1993, 1168, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, M.F.; Asker, M.M.S.; Ibrahim, Z.K. Functional bioactive compounds and biological activities of Spirulina platensis lipids. Czech J. Food Sci. 2008, 26, 211–222. [Google Scholar] [CrossRef]
- Sato, N.; Kamimura, R.; Tsuzuki, M. Dispensability of a sulfolipid for photoautotrophic cell growth and photosynthesis in a marine cyanobacterium, Synechococcus sp. PCC 7002. Biochem. Biophys. Res. Commun. 2016, 477, 854–860. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, M.; Tabata, S. Synechocystis sp. PCC 6803—A useful tool in the study of the genetics of cyanobacteria. Photosynth. Res. 2001, 70, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.; Sato, N. Fatty acid content and composition of triacylglycerols of Chlorella kessleri. Bio Protoc. 2018, 8, e2676. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Oka, A.; Sugisaki, H.; Takanami, M. Nucleotide sequence of the kanamycin resistance transposon Tn903. J. Mol. Biol. 1981, 147, 217–226. [Google Scholar] [CrossRef]
- Xiong, Q.; Feng, J.; Li, S.T.; Zhang, G.Y.; Qiao, Z.X.; Chen, Z.; Wu, Y.; Lin, Y.; Li, T.; Ge, F.; et al. Integrated transcriptomic and proteomic analysis of the global response of Synechococcus to high light stress. Mol. Cell. Proteom. 2015, 14, 1038–1053. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 16:0 PQH2 | m/z 1007 | 18:0 PQH2 | m/z 1035 | 16:0 PQ-C | m/z 1021 | 18:0 PQ-C | m/z 1049 | PQ-C | m/z 766 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | m/z 153.1 | + | m/z 265.3 | + | m/z 151.1 | + | m/z 151.1 | + | m/z 151.1 | + |
| 2 | m/z 237.2 | + | m/z 153.1 | + | m/z 109.1 | + | m/z 109.1 | + | m/z 95.1 | + |
| 3 | m/z 109.1 | + | m/z 149.1 | + | m/z 95.1 | + | m/z 95.1 | + | m/z 191.1 | + |
| 4 | m/z 149.1 | + | m/z 95.1 | + | m/z 149.1 | + | m/z 81.1 | + | m/z 109.1 | + |
| 5 | m/z 95.1 | + | m/z 135.1 | + | m/z 81.1 | + | m/z 149.1 | + | m/z 149.1 | + |
| 6 | m/z 123.1 | + | m/z 109.1 | + | m/z 135.1 | + | m/z 135.1 | + | m/z 135.1 | + |
| 7 | m/z 81.1 | + | m/z 123.1 | + | m/z 123.1 | + | m/z 123.1 | + | m/z 123.1 | + |
| 8 | m/z 135.1 | + | m/z 121.1 | + | m/z 121.1 | + | m/z 121.1 | + | m/z 81.1 | + |
| 9 | m/z 163.1 | + | m/z 247.2 | + | m/z 163.2 | + | m/z 163.1 | + | m/z 121.1 | + |
| 10 | m/z 151.1 | − | m/z 163.1 | + | m/z 203.2 | + | m/z 229.1 | + | m/z 163.1 | + |
| 11 | m/z 137.1 | + | m/z 116.1 | − | m/z 189.2 | + | m/z 161.1 | + | m/z 177.2 | + |
| 12 | m/z 177.2 | + | m/z 151.1 | + | m/z 137.1 | + | m/z 177.2 | + | m/z 217.1 | + |
| 13 | m/z 121.1 | + | m/z 217.2 | + | m/z 191.1 | + | m/z 191.1 | + | m/z 189.2 | + |
| 14 | m/z 191.2 | + | m/z 191.2 | + | m/z 177.2 | + | m/z 189.2 | + | m/z 161.1 | − |
| 15 | m/z 219.2 | + | m/z 135.1 | + | m/z 161.1 | + | m/z 203.2 | + | m/z 175.1 | + |
| 16 | m/z 203.2 | + | m/z 177.2 | + | m/z 175.2 | + | m/z 137.1 | − | m/z 147.1 | − |
| 17 | m/z 189.2 | + | m/z 69.1 | + | m/z 147.1 | − | m/z 175.1 | + | m/z 203.2 | + |
| 18 | m/z 217.2 | + | m/z 205.2 | − | m/z 107.1 | + | m/z 107.1 | + | m/z 107.1 | + |
| 19 | m/z 175.1 | − | m/z 81.1 | + | m/z 229.1 | + | m/z 147.1 | + | m/z 116.1 | − |
| 20 | m/z 97.1 | + | m/z 231.2 | + | m/z 191.2 | − | m/z 217.2 | + | m/z 137.1 | + |
| Coverage (%) | 90 | 90 | 90 | 95 | 85 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondo, M.; Aoki, M.; Hirai, K.; Ito, R.; Tsuzuki, M.; Sato, N. Plastoquinone Lipids: Their Synthesis via a Bifunctional Gene and Physiological Function in a Euryhaline Cyanobacterium, Synechococcus sp. PCC 7002. Microorganisms 2023, 11, 1177. https://doi.org/10.3390/microorganisms11051177
Kondo M, Aoki M, Hirai K, Ito R, Tsuzuki M, Sato N. Plastoquinone Lipids: Their Synthesis via a Bifunctional Gene and Physiological Function in a Euryhaline Cyanobacterium, Synechococcus sp. PCC 7002. Microorganisms. 2023; 11(5):1177. https://doi.org/10.3390/microorganisms11051177
Chicago/Turabian StyleKondo, Mimari, Motohide Aoki, Kazuho Hirai, Ryo Ito, Mikio Tsuzuki, and Norihiro Sato. 2023. "Plastoquinone Lipids: Their Synthesis via a Bifunctional Gene and Physiological Function in a Euryhaline Cyanobacterium, Synechococcus sp. PCC 7002" Microorganisms 11, no. 5: 1177. https://doi.org/10.3390/microorganisms11051177