


Impact of Water Shortage on Soil and Plant Attributes in the Presence of Arbuscular Mycorrhizal Fungi from a Harsh Environment
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
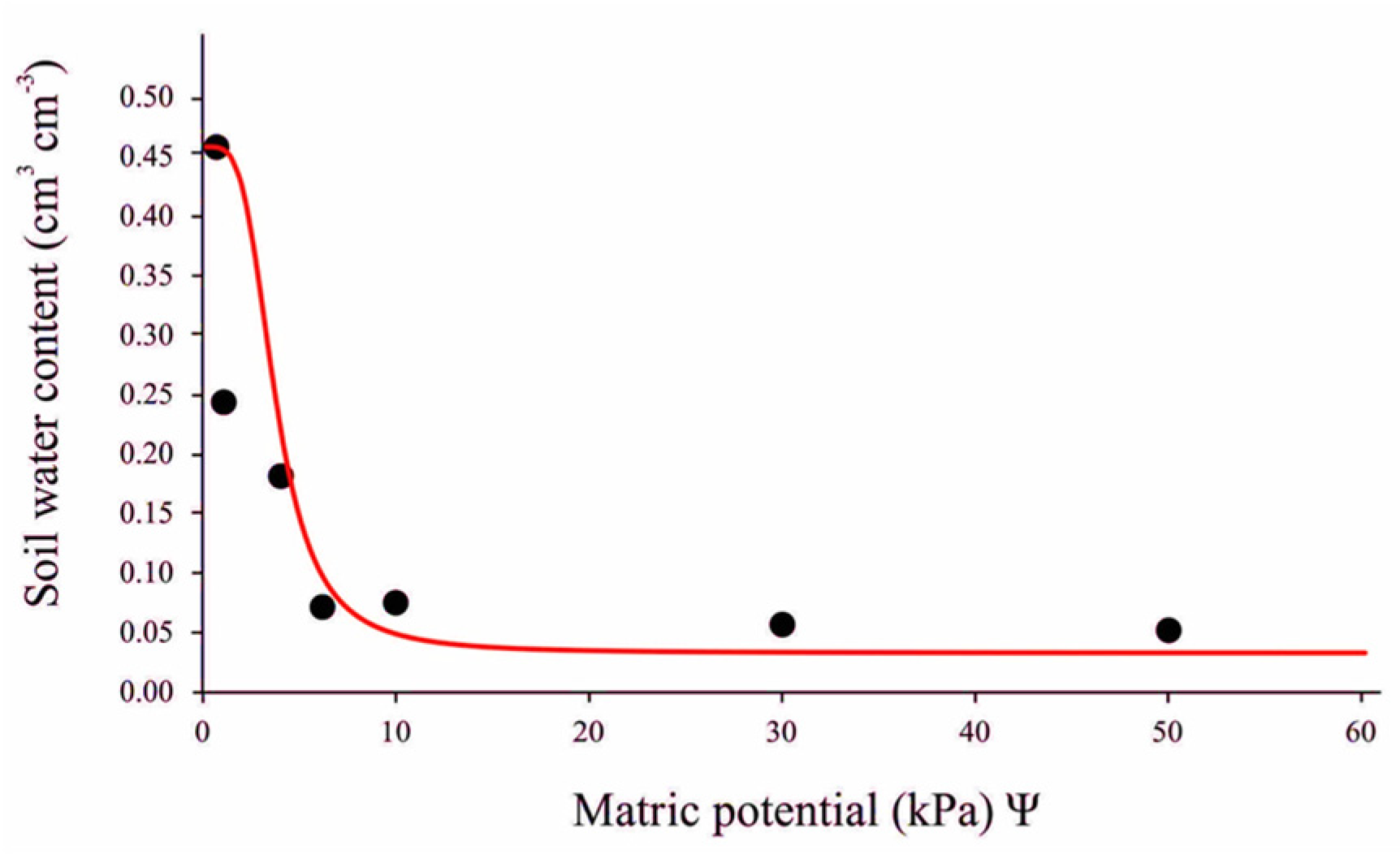
2.2. Water-Holding Capacity Determination and Experimental Management
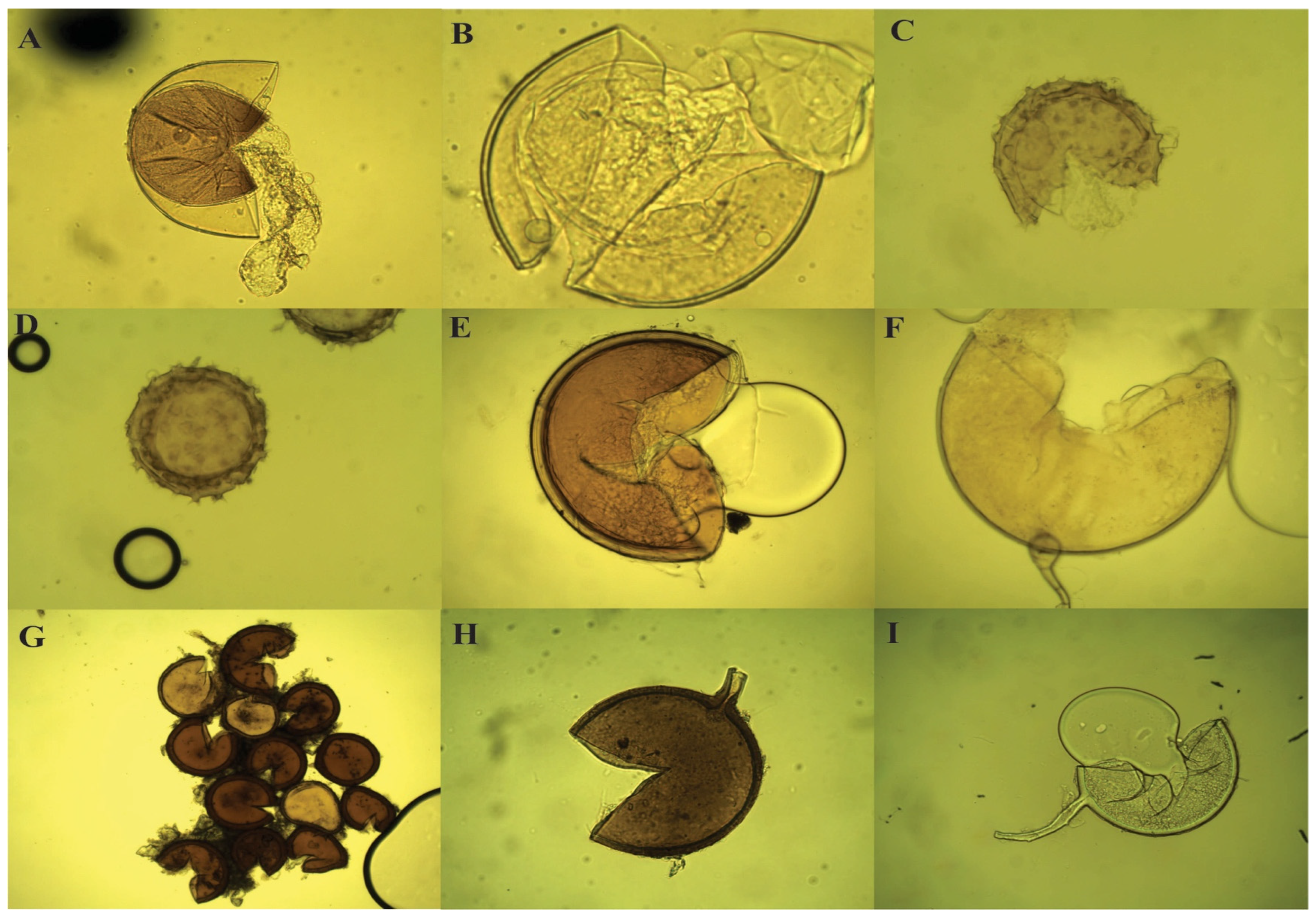
2.3. AMF Inoculum Characterization
2.4. Soil and Plant Sampling
2.5. Analytical Analyses
2.5.1. Plant Biomass and Nutrient Content
2.5.2. Soil and Plant Enzyme Activities
2.5.3. Soil Microbial Biomass (Carbon and Phosphorus)
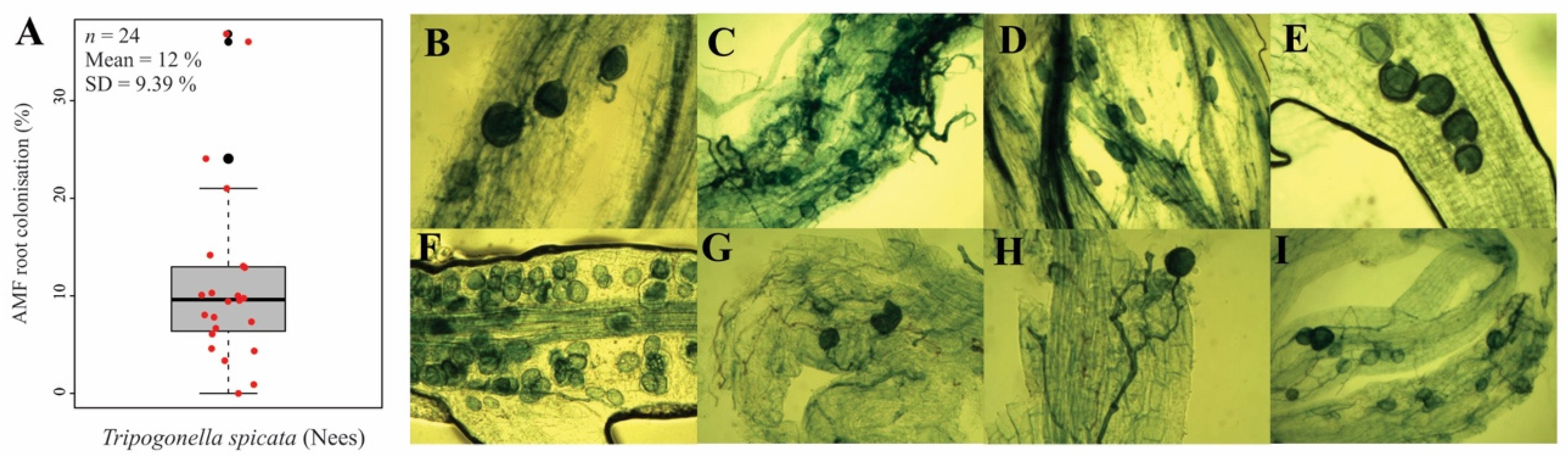
2.5.4. Soil Glomalin and AMF Root Colonisation
2.6. Data Analyses
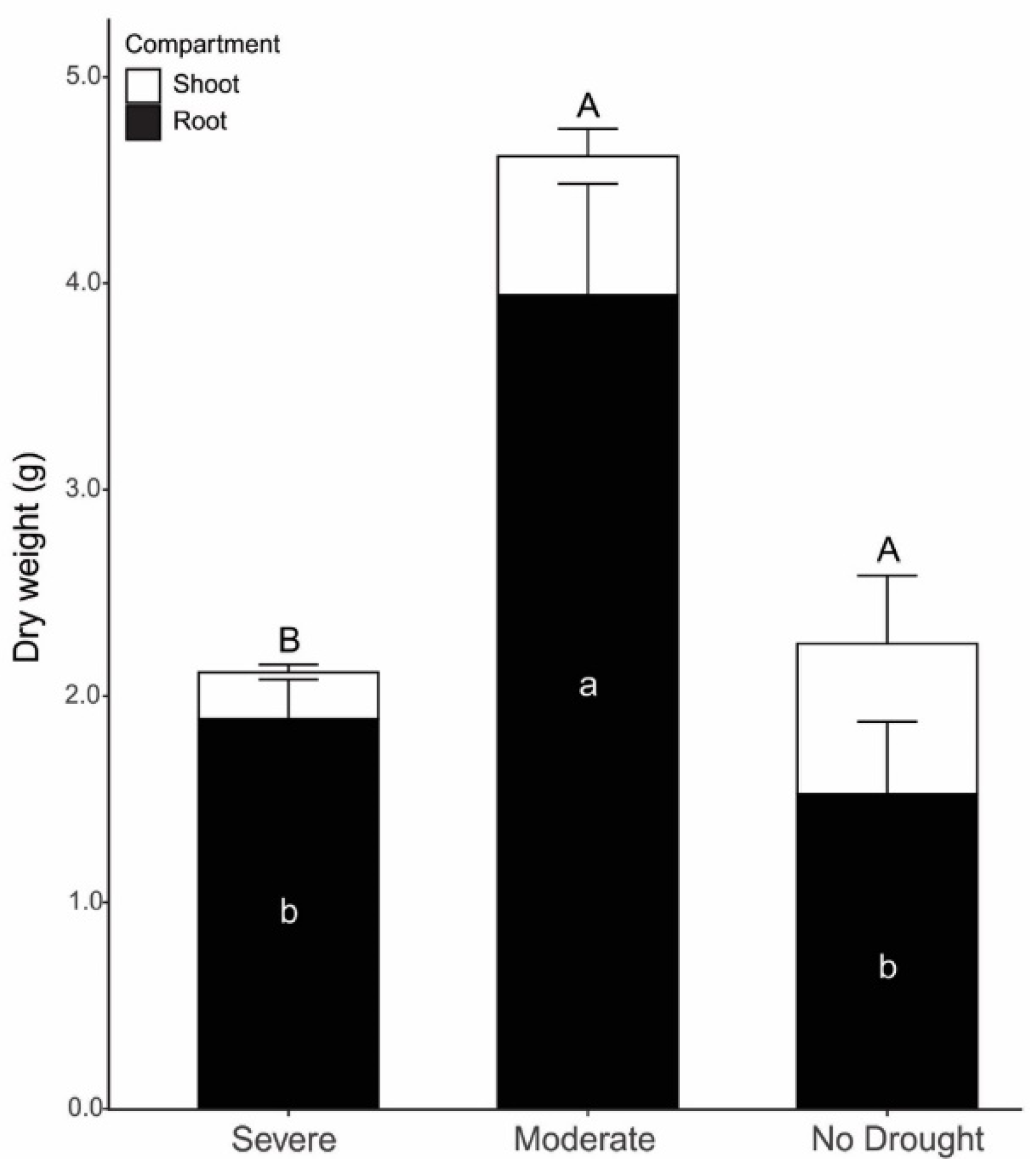
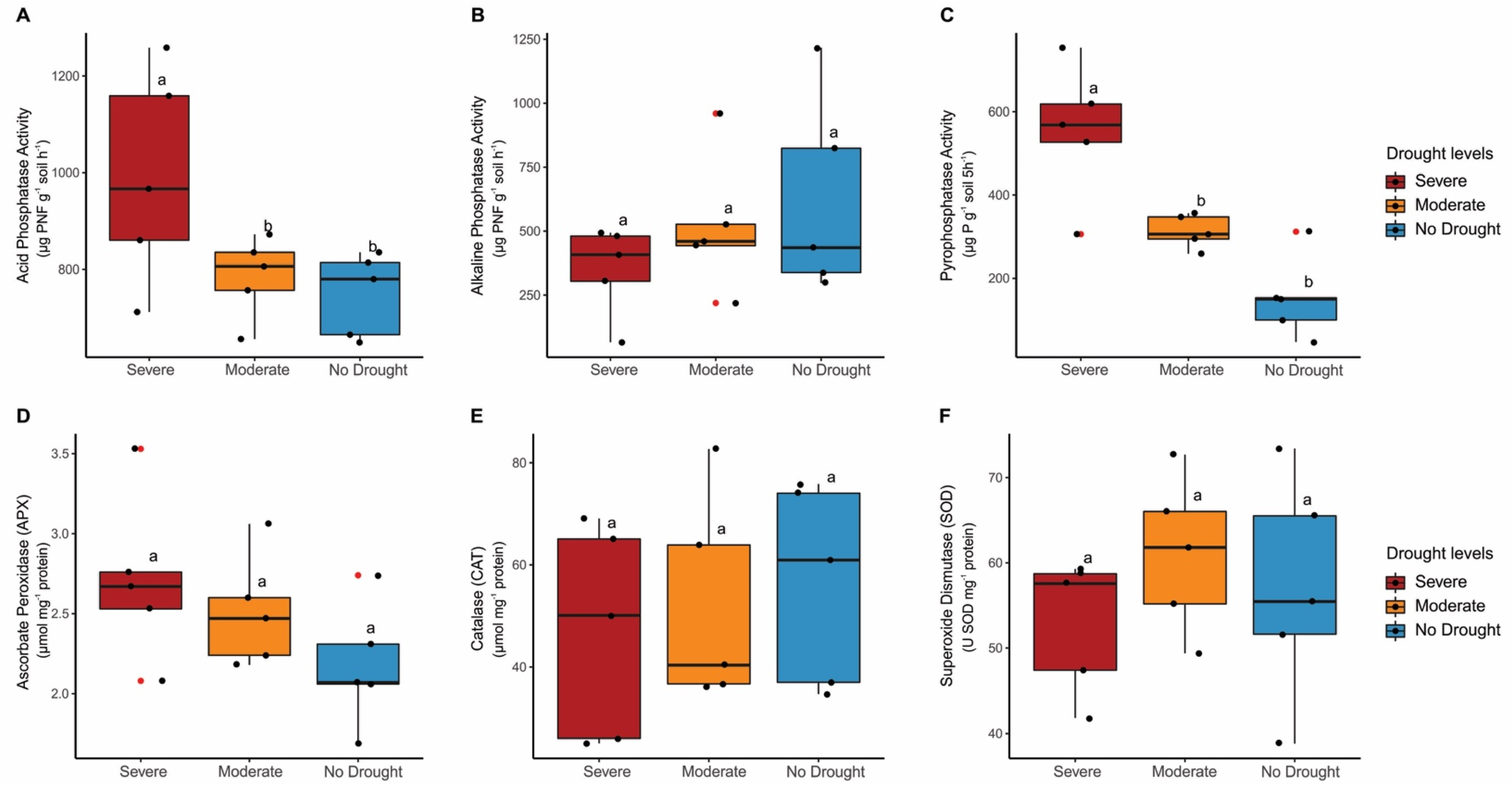
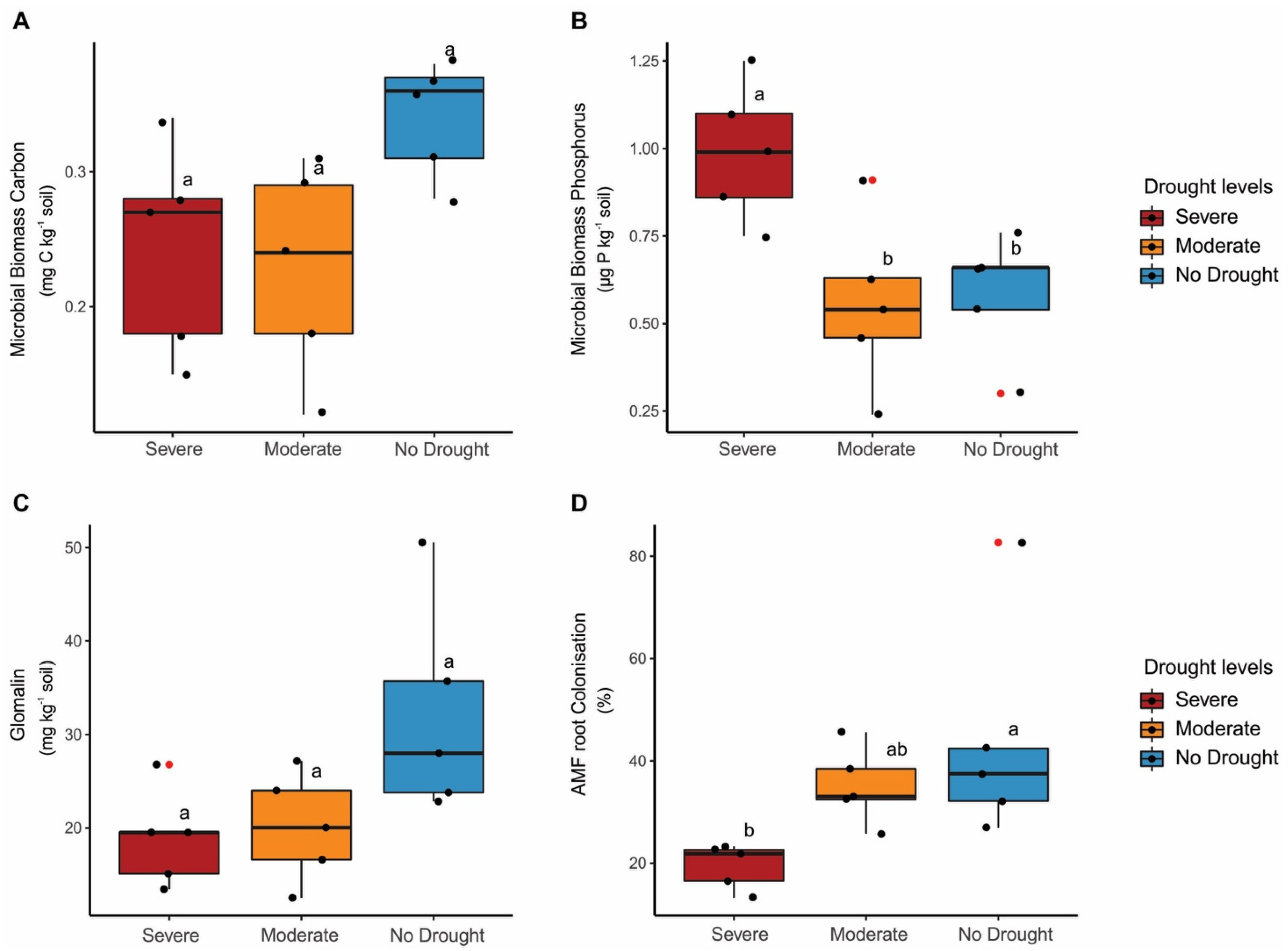
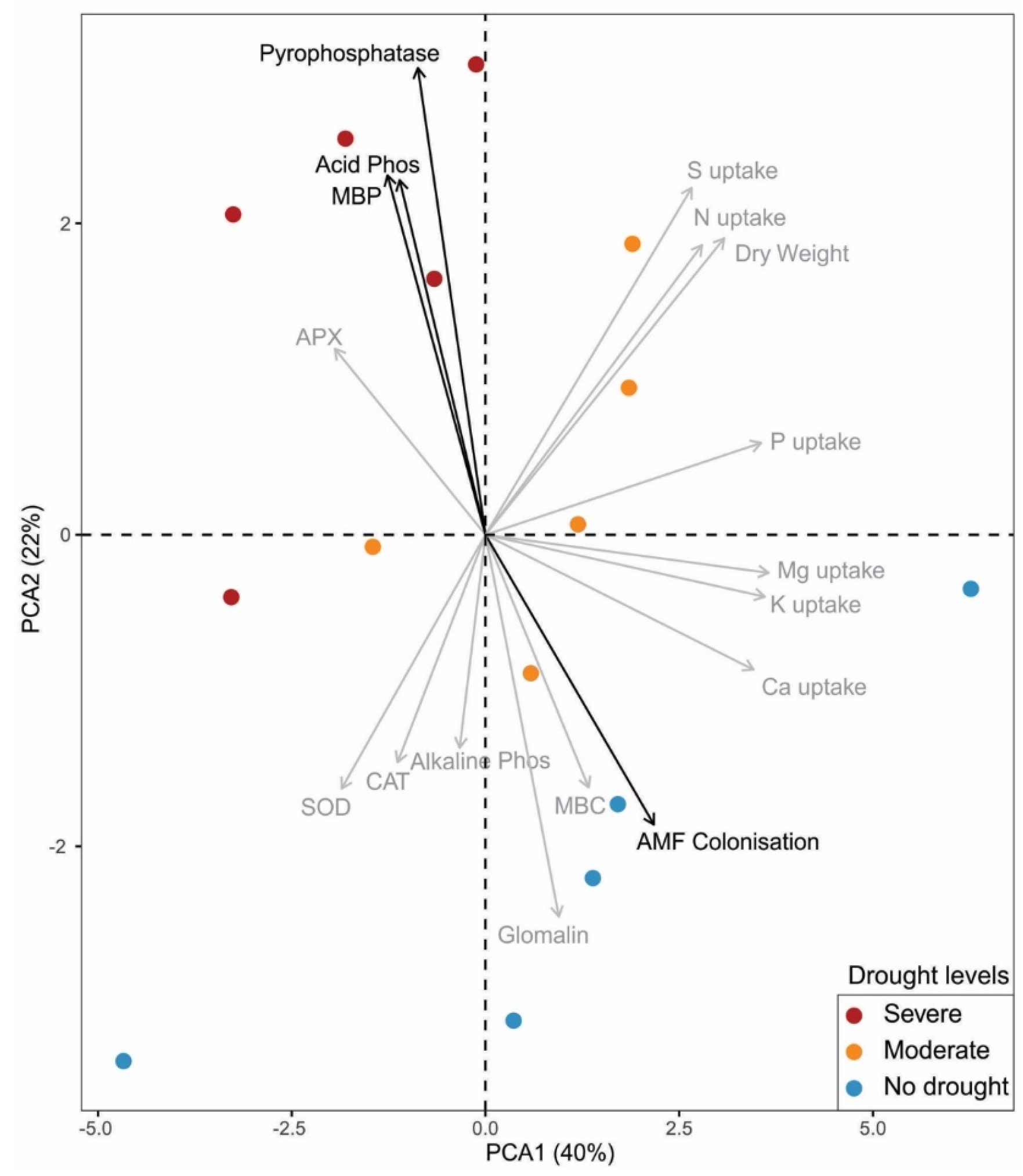
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

Appendix C

References
- Christian, J.I.; Basara, J.B.; Hunt, E.D.; Otkin, J.A.; Furtado, J.C.; Mishra, V.; Xiao, X.; Randall, R.M. Global Distribution, Trends, and Drivers of Flash Drought Occurrence. Nat. Commun. 2021, 12, 6330. [Google Scholar] [CrossRef] [PubMed]
- Neupane, J.; Guo, W. Agronomic basis and strategies for precision water management: A review. Agronomy 2019, 9, 87. [Google Scholar] [CrossRef]
- Muthuvel, D.; Sivakumar, B.; Mahesha, A. Future global concurrent droughts and their effects on maize yield. Sci. Total Environ. 2023, 855, 158860. [Google Scholar] [CrossRef] [PubMed]
- Reuters. Brazil Drought Threatening National Output Potential, Southern Farmers Say. Available online: https://www.reuters.com/world/americas/brazil-drought-threatening-national-output-potential-southern-farmers-say-2023-02-14/ (accessed on 15 April 2023).
- Rossi, J.B.; Ruhoff, A.; Fleischmann, A.S.; Laipelt, L. Drought Propagation in Brazilian Biomes Revealed by Remote Sensing. Remote Sens. 2023, 15, 454. [Google Scholar] [CrossRef]
- Shiferaw, B.; Prasanna, B.M.; Hellin, J.; Bänziger, M. Crops that feed the world 6. Past successes and future challenges to the role played by maize in global food security. Food Sec. 2011, 3, 307–327. [Google Scholar] [CrossRef]
- Erenstein, O.; Jaleta, M.; Sonder, K.; Mottaleb, K.; Prasanna, B.M. Global maize production, consumption and trade: Trends and R&D implications. Food Sec. 2022, 14, 1295–1319. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: San Diego, CA, USA, 2008; pp. 11–145. [Google Scholar]
- Tedersoo, L.; Bahram, M. Mycorrhizal Types Differ in Ecophysiology and Alter Plant Nutrition and Soil Processes. Biol. Rev. 2019, 94, 1857–1880. [Google Scholar] [CrossRef]
- Redecker, D.; Kodner, R.; Graham, L.E. Glomalean Fungi from the Ordovician. Science 2000, 289, 1920–1921. [Google Scholar] [CrossRef]
- Brundrett, M.C.; Tedersoo, L. Evolutionary History of Mycorrhizal Symbioses and Global Host Plant Diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef]
- Delavaux, C.S.; Smith-Ramesh, L.M.; Kuebbing, S.E. Beyond Nutrients: A Meta-Analysis of the Diverse Effects of Arbuscular Mycorrhizal Fungi on Plants and Soils. Ecology 2017, 98, 2111–2119. [Google Scholar] [CrossRef]
- Rasmann, S.; Bennett, A.; Biere, A.; Karley, A.; Guerrieri, E. Root Symbionts: Powerful Drivers of Plant above- and Belowground Indirect Defenses. Insect Sci. 2017, 24, 947–960. [Google Scholar] [CrossRef]
- Tedersoo, L. Biogeography of mycorrhizal symbiosis, 1st ed.; Springer: New York, NY, USA, 2017. [Google Scholar] [CrossRef]
- Deng, M.; Hu, S.; Guo, L.; Jiang, L.; Huang, Y.; Schmid, B.; Liu, C.; Chang, P.; Li, S.; Liu, X.; et al. Tree Mycorrhizal Association Types Control Biodiversity-Productivity Relationship in a Subtropical Forest. Sci. Adv. 2023, 9, eadd4468. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.M.M.; Jones, D.L.; Chadwick, D.R.; Qi, X.; Cotta, S.R.; Araújo, V.L.V.P.; Matteoli, F.P.; Lacerda-Júnior, G.V.; Pereira, A.P.A.; Fernandes-Júnior, P.I.; et al. Can Arbuscular Mycorrhizal Fungi and Rhizobacteria Facilitate 33P Uptake in Maize Plants under Water Stress? Microbiol. Res. 2023, 271, 127350. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Abd Elgawad, H.; Xiong, Y.C.; Macovei, A.; Brestic, M.; Skalicky, M.; Shaghaleh, H.; Alhaj Hamoud, Y.; El-Sawah, A.M. Inoculation with Bacillus amyloliquefaciens and mycorrhiza confers tolerance to drought stress and improve seed yield and quality of soybean plant. Physiol. Plant. 2021, 172, 2153–2169. [Google Scholar] [CrossRef] [PubMed]
- Sheteiwy, M.S.; Ali, D.F.; Xiong, Y.C.; Brestic, M.; Skalicky, M.; Hamoud, Y.A.; Ulhassan, Z.; Shaghaleh, H.; AbdElgawad, H.; Farooq, M.; et al. Physiological and biochemical responses of soybean plants inoculated with Arbuscular mycorrhizal fungi and Bradyrhizobium under drought stress. BMC Plant Biol. 2021, 21, 195. [Google Scholar] [CrossRef] [PubMed]
- El-Sawah, A.M.; Abdel-Fattah, G.G.; Holford, P.; Korany, S.M.; Alsherif, E.A.; AbdElgawad, H.; Ulhassan, Z.; Jośko, I.; Ali, B.; Sheteiwy, M.S. Funneliformis constrictum modulates polyamine metabolism to enhance tolerance of Zea mays L. to salinity. Microbiol. Res. 2023, 266, 127254. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Ahmed, M.; Korany, S.M.; Alsherif, E.A.; Mowafy, A.M.; Chen, J.; Jośko, I.; Selim, S.; AbdElgawad, H. Arbuscular Mycorrhizal Fungus “Rhizophagus irregularis” impacts on physiological and biochemical responses of ryegrass and chickpea plants under beryllium stress. Environ. Pollut. 2022, 315, 120356. [Google Scholar] [CrossRef]
- Augé, R.M. Water Relations, Drought and Vesicular-Arbuscular Mycorrhizal Symbiosis. Mycorrhiza 2001, 11, 3–42. [Google Scholar] [CrossRef]
- Cheng, S.; Zou, Y.N.; Kuča, K.; Hashem, A.; Abd-Allah, E.F.; Wu, Q.S. Elucidating the Mechanisms Underlying Enhanced Drought Tolerance in Plants Mediated by Arbuscular Mycorrhizal Fungi. Front. Microbiol. 2021, 12, 809473. [Google Scholar] [CrossRef]
- Agnihotri, R.; Sharma, M.P.; Prakash, A.; Ramesh, A.; Bhattacharjya, S.; Patra, A.K.; Manna, M.C.; Kurganova, I.; Kuzyakov, Y. Glycoproteins of Arbuscular Mycorrhiza for Soil Carbon Sequestration: Review of Mechanisms and Controls. Sci. Total Environ. 2022, 806, 150571. [Google Scholar] [CrossRef]
- WRB-FAO. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; Food and Agriculture Organization of the United Nations: Rome, Italy, 2015. [Google Scholar]
- van Raij, B.; Andrade, J.C.; Cantarella, H.; Quaggio, J.A. Análise química para avaliação da fertilidade de solos tropicais = Chemical analysis for fertility assessment of tropical soils. In Boletim Técnico Instituto Agronômico de Campinas; IAC: Campinas, Brazil, 2001. [Google Scholar]
- van Genuchten, M.T. A closed-form equation for predicting the hydraulic conductivity of unsaturated soils. SSSA J. 1980, 44, 892–898. [Google Scholar] [CrossRef]
- de Lima, R.P.; da Silva, A.R.; da Silva, A.P.; Leão, T.P.; Mosaddeghi, M.R. Soilphysics: An R package for calculating soil water availability to plants by different soil physical indices. Comput. Electron. Agric. 2016, 120, 63–71. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2. WIREs Comp. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 32. [Google Scholar]
- Xiao, L.; Zhao, M.; Liu, G.; Li, P.; Liu, F.; Xue, S. Soil biochemical index-based assessment of the effect of drought stress on the rhizosphere soil quality in three typical grass species in the Loess Plateau, China. J. Soils Sediments 2022, 22, 2982–2994. [Google Scholar] [CrossRef]
- Sendek, A.; Karakoç, C.; Wagg, C.; Domínguez-Begines, J.; do Couto, G.M.; van der Heijden, M.G.; Naz, A.A.; Lochner, A.; Chatzinotas, A.; Klotz, S.; et al. Drought modulates interactions between arbuscular mycorrhizal fungal diversity and barley genotype diversity. Sci. Rep. 2019, 9, 9650. [Google Scholar] [CrossRef]
- Torres-Arias, Y.; Fors, R.O.; Nobre, C.; Gómez, E.F.; Berbara, R.L.L. Production of Native Arbuscular Mycorrhizal Fungi Inoculum under Different Environmental Conditions. Braz. J. Microbiol. 2017, 48, 87–94. [Google Scholar] [CrossRef]
- Fernandes-Júnior, P.I.; de Tarso Aidar, S.; Morgante, C.V.; Gava, C.A.T.; Zilli, J.E.; Souza, L.S.B.; Marinho, R.C.N.; Nóbrega, R.S.A.; Brasil, M.S.; Seido, S.L.; et al. The resurrection plant Tripogon spicatus (Poaceae) harbors a diversity of plant growth promoting bacteria in northeastern Brazilian caatinga. Rev. Bras. Cienc. Solo 2015, 39, 993–1002. [Google Scholar] [CrossRef]
- Aidar, S.D.T.; Chaves, A.R.D.M.; Fernandes-Júnior, P.I.; Oliveira, M.S.; Costa Neto, B.P.; Junior, T.C.; Morgante, C.V. Vegetative desiccation tolerance of Tripogon spicatus (Poaceae) from the tropical semiarid region of northeastern Brazil. Funct. Plant Biol. 2017, 44, 1124–1133. [Google Scholar] [CrossRef]
- Kavamura, V.N.; Santos, S.N.; da Silva, J.L.; Parma, M.M.; Ávila, L.A.; Visconti, A.; Zucchi, T.D.; Taketani, R.G.; Andreote, F.D.; de Melo, I.S. Screening of Brazilian cacti rhizobacteria for plant growth promotion under drought. Microbiol. Res. 2013, 168, 183–191. [Google Scholar] [CrossRef]
- Silva, A.M.M.; Feiler, H.P.; Lacerda-Júnior, G.V.; Fernandes-Júnior, P.I.; Aidar, S.T.; Araújo, V.L.V.P.; Matteoli, F.P.; Pereira, A.P.A.; de Melo, I.S.; Cardoso, E.J.B.N. Arbuscular mycorrhizal fungi associated with the rhizosphere of an endemic terrestrial bromeliad and a grass in the Brazilian neotropical dry forest. Braz. J. Microbiol. 2022; submitted. [Google Scholar]
- Malavolta, E.; Vitti, G.C.; Oliveira, S.B. Assessment of the Nutritional Status of Plants-Principles and Applications, 2nd ed.; Potafos: Piracicaba, Brazil, 1997. [Google Scholar]
- Dick, W.A.; Tabatabai, M.A. Inorganic pyrophosphatase activity of soils. Soil Biol. Biochem. 1978, 10, 58–65. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil enzymes. In Methods of Soil Analysis: Part 2 Microbiological and Biochemical Properties; Weaver, R.W., Angle, S., Bottomley, P., Bezdiecek, D., Eds.; Soil Science Society of American: Madison, WI, USA, 1994; pp. 775–833. [Google Scholar]
- Azevedo, R.A.; Alas, R.M.; Smith, R.J.; Lea, P.J. Response of antioxidant enzymes to transfer from elevated carbon dioxide to air and ozone fumigation, in the leaves and root of wild-type and a catalase-deficient mutant of barley. Physiol. Plant. 1998, 104, 280–292. [Google Scholar] [CrossRef]
- Moldes, C.A.; Medici, L.O.; Abrahão, O.S.; Tsai, S.M.; Azevedo, R.A. Biochemical responses of glyphosate resistant and susceptible soybean plants exposed to glyphosate. Acta Physiol. Plant 2008, 30, 469–479. [Google Scholar] [CrossRef]
- Kraus, T.E.; McKersie, B.D.; Fletcher, R.A. Paclobutrazol-induced tolerance of wheat leaves to paraquat may involve increased antioxidant enzyme activity. J. Plant Physiol. 1995, 145, 570–576. [Google Scholar] [CrossRef]
- Ciannopolitis, C.N.; Ries, S.K. Superoxide dismutase I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Cembrowska-Lech, D.; Koprowski, M.; Kępczyński, J. Germination induction of dormant Avena fatua caryopses by KAR1 and GA3 involving the control of reactive oxygen species (H2O2 and O2•−) and enzymatic antioxidants (superoxide dismutase and catalase) both in the embryo and the aleurone layers. J. Plant Physiol. 2015, 176, 169–179. [Google Scholar] [CrossRef]
- Brookes, P.C.; Powlson, D.S.; Jenkinson, D.S. Measurement of microbial biomass phosphorus in soil. Soil Biol. Biochem. 1982, 14, 319–329. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.F.; Upadhyaya, A. Extraction of an abundant and unusual protein from soil and comparison with hyphal protein from arbuscular mycorrhizal fungi. Soil Sci. 1996, 161, 575–586. [Google Scholar] [CrossRef]
- Koske, R.E.; Gemma, J.N. A modified procedure for staining roots to detect VA mycorrhizas. Mycol. Res. 1989, 92, 486–488. [Google Scholar] [CrossRef]
- Vierheilig, H.; Coughlan, A.P.; Wyss, U.; Piché, Y. Ink and vinegar, a simple staining technique for arbuscular-mycorrhizal fungi. Appl. Environ. Microbiol. 1998, 64, 5004–5007. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular—Arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Ferreira, E.B.; Cavalcanti, P.P.; Nogueira, D.A. ExpDes: An R package for ANOVA and experimental designs. Appl. Math. 2014, 5, 51204. [Google Scholar] [CrossRef]
- Ramette, A. Multivariate analyses in microbial ecology. FEMS Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Factoextra: Extract and visualize the results of multivariate data analyses. R Package Version 2017, 1, 337–354. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. 2020. Available online: https://www.r-project.org/ (accessed on 17 January 2022).
- Rubin, J.A.; Görres, J.H. Effects of mycorrhizae, plants, and soils on phosphorus leaching and plant uptake: Lessons learned from a mesocosm study. Plants People Planet 2022, 4, 403–415. [Google Scholar] [CrossRef]
- Poorter, H.; Karl, J.N.; Peter, B.R.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Metaanalyses of interspecific variation and environmental control. New Phytol. 2011, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Peng, L.; Xiao, L.; Wang, W.; Yu, K.X.; Shi, P. Heterogeneity in short-term allocation of carbon to roots of Pinus tabuliformis seedlings and root respiration under drought stress. Plant Soil. 2020, 452, 359–378. [Google Scholar] [CrossRef]
- Maltz, M.R.; Treseder, K.K. Sources of inocula influence mycorrhizal colonization of plants in restoration projects: A meta-analysis. Restor. Ecol. 2015, 23, 625–634. [Google Scholar] [CrossRef]
- Medeiros, A.S.; Scaloppi, J.C.; Damasceno, E.S.; Goto, B.T.; Vieira, D.C.; Socolowski, F.; Rodrigues, R.G.; Yano-Melo, A.M. Arbuscular mycorrhizal fungi communities shaped by host-plant affect the outcome of plant–soil feedback in dryland restoration. J. Appl. Ecol. 2022, 60, 507–518. [Google Scholar] [CrossRef]
- Rúa, M.A.; Antoninka, A.; Antunes, P.M.; Chaudhary, V.B.; Gehring, C.; Lamit, L.J.; Piculell, B.J.; Bever, J.D.; Zabinski, C.; Meadow, J.F.; et al. Home-field advantage? evidence of local adaptation among plants, soil, and arbuscular mycorrhizal fungi through meta- analysis. BMC Evol. Biol. 2016, 16, 122. [Google Scholar] [CrossRef]
- Liu, X.; Mei, S.; Salles, J.F. Do inoculated microbial consortia perform better than single strains in living soil? A meta-analysis. BioRxiv 2023. [Google Scholar] [CrossRef]
- Yadav, R.; Ror, P.; Beniwal, R.; Kumar, S.; Ramakrishna, W. Bacillus sp. and arbuscular mycorrhizal fungi consortia enhance wheat nutrient and yield in the second-year field trial: Superior performance in comparison with chemical fertilizers. J. Appl. Microbiol. 2022, 132, 2203–2219. [Google Scholar] [CrossRef]
- Leite, R.D.C.; Pereira, Y.C.; Oliveira-Paiva, C.A.D.; Moraes, A.J.G.D.; Silva, G.B.D. Increase in yield, leaf nutrient, and profitability of soybean co-inoculated with Bacillus strains and Arbuscular mycorrhizal fungi. Rev. Bras. Cienc. Solo 2022, 46, e0220007. [Google Scholar] [CrossRef]
- Prasanna Kumar, B.; Trimurtulu, N.; Vijaya Gopal, A.; Nagaraju, Y. Impact of Culturable Endophytic Bacteria on Soil Aggregate Formation and Peanut (Arachis hypogaea L.) Growth and Yield Under Drought Conditions. Curr. Microbiol. 2022, 79, 308. [Google Scholar] [CrossRef]
- Bogati, K.; Walczak, M. The impact of drought stress on soil microbial community, enzyme activities and plants. Agronomy 2022, 12, 189. [Google Scholar] [CrossRef]
- Hosseini, S.S.; Lakzian, A.; Razavi, B.S. Reduction in root active zones under drought stress controls spatial distribution and catalytic efficiency of enzyme activities in rhizosphere of wheat. Rhizosphere 2022, 23, 100561. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Increasing drought decreases phosphorus availability in an evergreen Mediterranean forest. Plant Soil 2004, 267, 367–377. [Google Scholar] [CrossRef]
- He, Z.L.; Wu, J.; O’Donnell, A.G.; Syers, J.K. Seasonal responses in microbial biomass carbon, phosphorus and sulphur in soils under pasture. Biol. Fertil. Soils 1997, 24, 421–428. [Google Scholar] [CrossRef]
- Arenberg, M.R.; Arai, Y. Uncertainties in soil physicochemical factors controlling phosphorus mineralization and immobilization processes. Adv. Agron. 2019, 154, 153–200. [Google Scholar] [CrossRef]
- Richardson, A.E.; Simpson, R.J. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef]
- de Vries, F.T.; Griffiths, R.I.; Knight, C.G.; Nicolitch, O.; Williams, A. Harnessing rhizosphere microbiomes for drought-resilient crop production. Science 2020, 368, 270–274. [Google Scholar] [CrossRef]
- Chareesri, A.; De Deyn, G.B.; Sergeeva, L.; Polthanee, A.; Kuyper, T.W. Increased arbuscular mycorrhizal fungal colonization reduces yield loss of rice (Oryza sativa L.) under drought. Mycorrhiza 2020, 30, 315–328. [Google Scholar] [CrossRef]
- Li, J.; Meng, B.; Chai, H.; Yang, X.; Song, W.; Li, S.; Lu, A.; Zhang, T.; Sun, W. Arbuscular mycorrhizal fungi alleviate drought stress in C3 (Leymus chinensis) and C4 (Hemarthria altissima) grasses via altering antioxidant enzyme activities and photosynthesis. Front. Plant Sci. 2019, 10, 499. [Google Scholar] [CrossRef]
- Amer, S.O.; Aliat, T.; Kucher, D.E.; Bensaci, O.A.; Rebouh, N.Y. Investigating the Potential of Arbuscular Mycorrhizal Fungi in Mitigating Water Deficit Effects on Durum Wheat (Triticum durum Desf.). Agriculture 2023, 13, 552. [Google Scholar] [CrossRef]
- Chandrasekaran, M. Arbuscular Mycorrhizal Fungi Mediated Alleviation of Drought Stress via Non-Enzymatic Antioxidants: A Meta-Analysis. Plants 2022, 11, 2448. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AMF Species | Number of Spores/50 g of Inoculum |
|---|---|
| Acaulospora morrowiae Spain and N.C. Schenck | 94 |
| Ambispora sp. | 417 |
| Gigaspora decipiens I.R. Hall and L.K. Abbott | 63 |
| Gigaspora gigantea (T.H. Nicholson and Gerd) Gerd. and Trappe | 9 |
| Glomus glomerulatum Sieverding | 7 |
| Glomus sp. | 9 |
| Rhizophagus clarus (T.H. Nicholson and N.C. Schenck) C. Walker and A. Schüßler) | 21 |
| Total | 620 |
| Compartment/Drought Levels | N | P | K | Ca | Mg | S |
|---|---|---|---|---|---|---|
| Shoot | mg SDW−1 | |||||
| Severe | 3.80 ± 1.90 a * | 0.22 ± 0.07 a | 7.16 ± 1.97 a | 1.08 ± 0.35 a | 1.13 ± 0.42 a | 0.43 ± 0.13 a |
| Moderate | 8.58 ± 4.00 a | 0.61 ± 0.40 a | 23.13 ± 11.65 a | 3.24 ± 1.63 a | 3.21 ± 1.75 a | 1.09 ± 0.54 a |
| No Drought | 7.45 ± 5.31 a | 0.74 ± 0.75 a | 21.43 ± 17.00 a | 3.12 ± 2.86 a | 3.33 ± 3.61 a | 1.23 ± 1.26 a |
| Root | mg RDW−1 | |||||
| Severe | 12.10 ± 6.02 a | 1.43 ± 0.74 a | 6.70 ± 3.26 a | 1.68 ± 0.90 a | 0.89 ± 0.48 a | 5.22 ± 2.66 a |
| Moderate | 15.47 ± 9.78 a | 2.61 ± 1.61 a | 7.54 ± 6.28 a | 1.96 ± 1.30 a | 0.93 ± 0.71 a | 8.25 ± 5.48 a |
| No Drought | 10.01 ± 10.22 a | 2.97 ± 2.69 a | 21.03 ± 17.04 a | 12.34 ± 11.51 a | 3.03 ± 2.74 a | 3.79 ± 3.05 a |
| Total dry weight | mg plant−1 | |||||
| Severe | 15.90 ± 6.66 a | 1.65 ± 0.75 a | 13.86 ± 3.22 a | 2.76 ± 0.99 a | 2.02 ± 0.71 a | 5.65 ± 2.65 a |
| Moderate | 24.05 ± 9.45 a | 3.22 ± 1.57 a | 30.67 ± 9.45 a | 5.20 ± 1.67 a | 4.14 ± 1.54 a | 9.34 ± 5.36 a |
| No Drought | 17.46 ± 13.86 a | 3.71 ± 3.33 a | 42.46 ± 30.63 a | 15.46 ± 13.78 a | 6.36 ± 6.13 a | 5.02 ± 4.08 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, A.M.M.; Feiler, H.P.; Qi, X.; de Araújo, V.L.V.P.; Lacerda-Júnior, G.V.; Fernandes-Júnior, P.I.; Cardoso, E.J.B.N. Impact of Water Shortage on Soil and Plant Attributes in the Presence of Arbuscular Mycorrhizal Fungi from a Harsh Environment. Microorganisms 2023, 11, 1144. https://doi.org/10.3390/microorganisms11051144
Silva AMM, Feiler HP, Qi X, de Araújo VLVP, Lacerda-Júnior GV, Fernandes-Júnior PI, Cardoso EJBN. Impact of Water Shortage on Soil and Plant Attributes in the Presence of Arbuscular Mycorrhizal Fungi from a Harsh Environment. Microorganisms. 2023; 11(5):1144. https://doi.org/10.3390/microorganisms11051144
Chicago/Turabian StyleSilva, Antonio Marcos Miranda, Henrique Petry Feiler, Xue Qi, Victor Lucas Vieira Prudêncio de Araújo, Gileno Vieira Lacerda-Júnior, Paulo Ivan Fernandes-Júnior, and Elke Jurandy Bran Nogueira Cardoso. 2023. "Impact of Water Shortage on Soil and Plant Attributes in the Presence of Arbuscular Mycorrhizal Fungi from a Harsh Environment" Microorganisms 11, no. 5: 1144. https://doi.org/10.3390/microorganisms11051144