
Grass–Legume Mixture with Rhizobium Inoculation Enhanced the Restoration Effects of Organic Fertilizer


Abstract
:1. Introduction
2. Materials and Methods
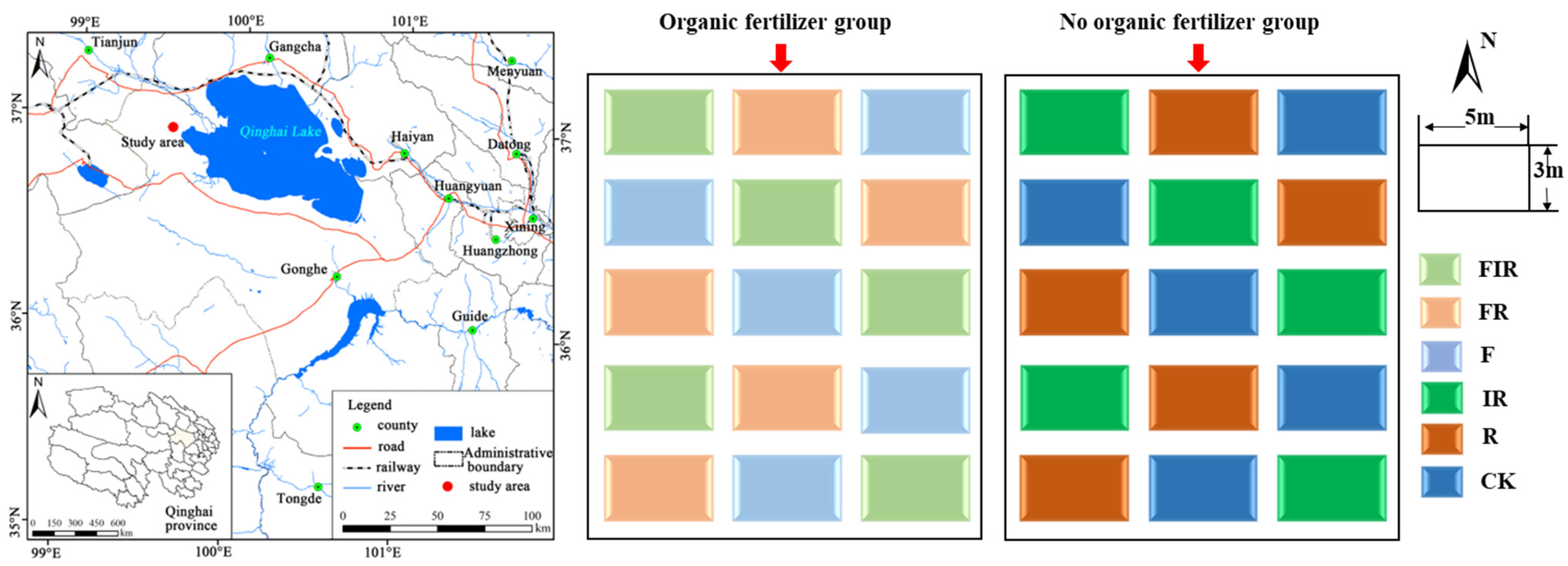
2.1. Site Overview and Study Design
2.2. Vegetation and Soil Sampling
2.3. Determination of Soil Nutrient Contents
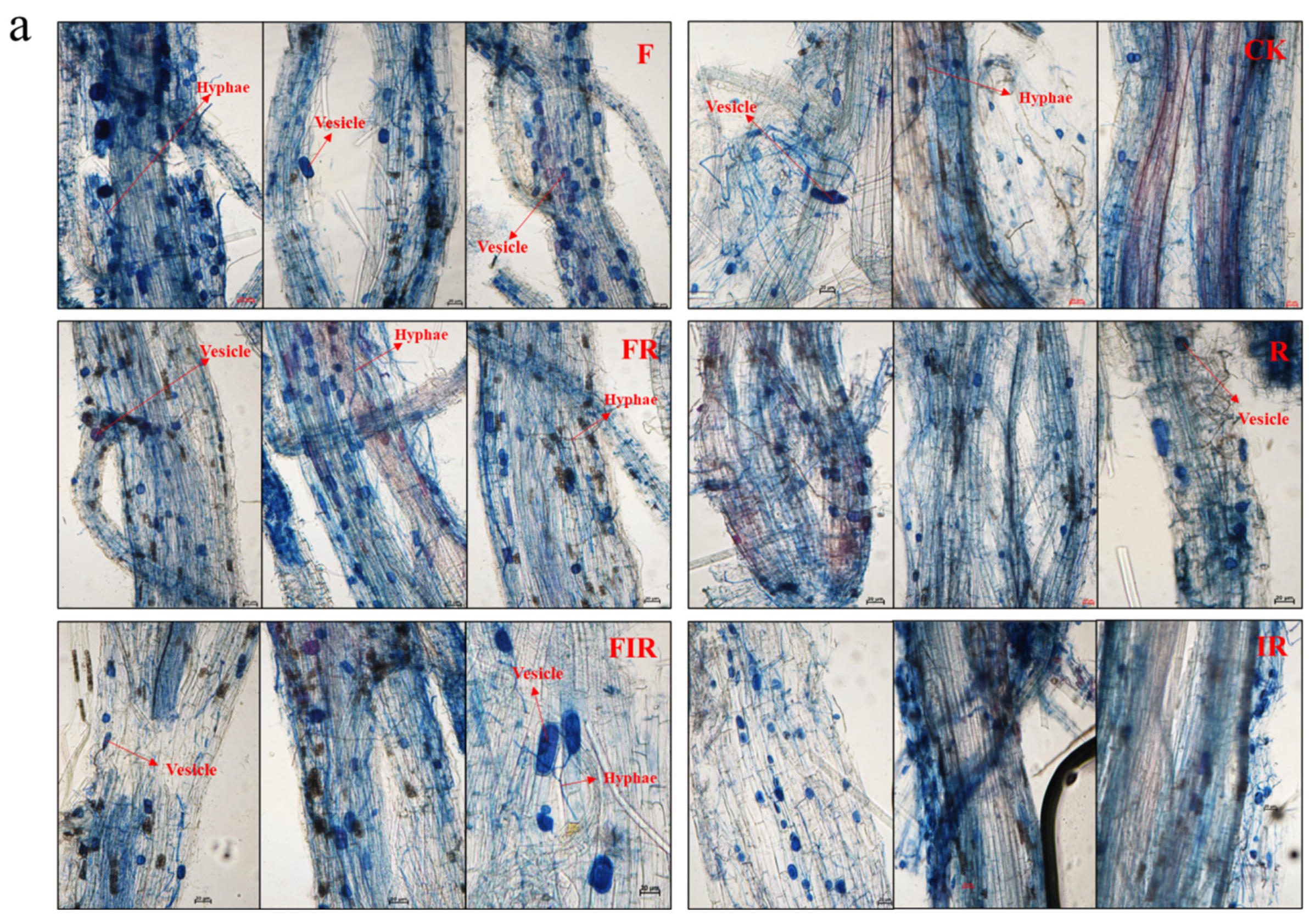
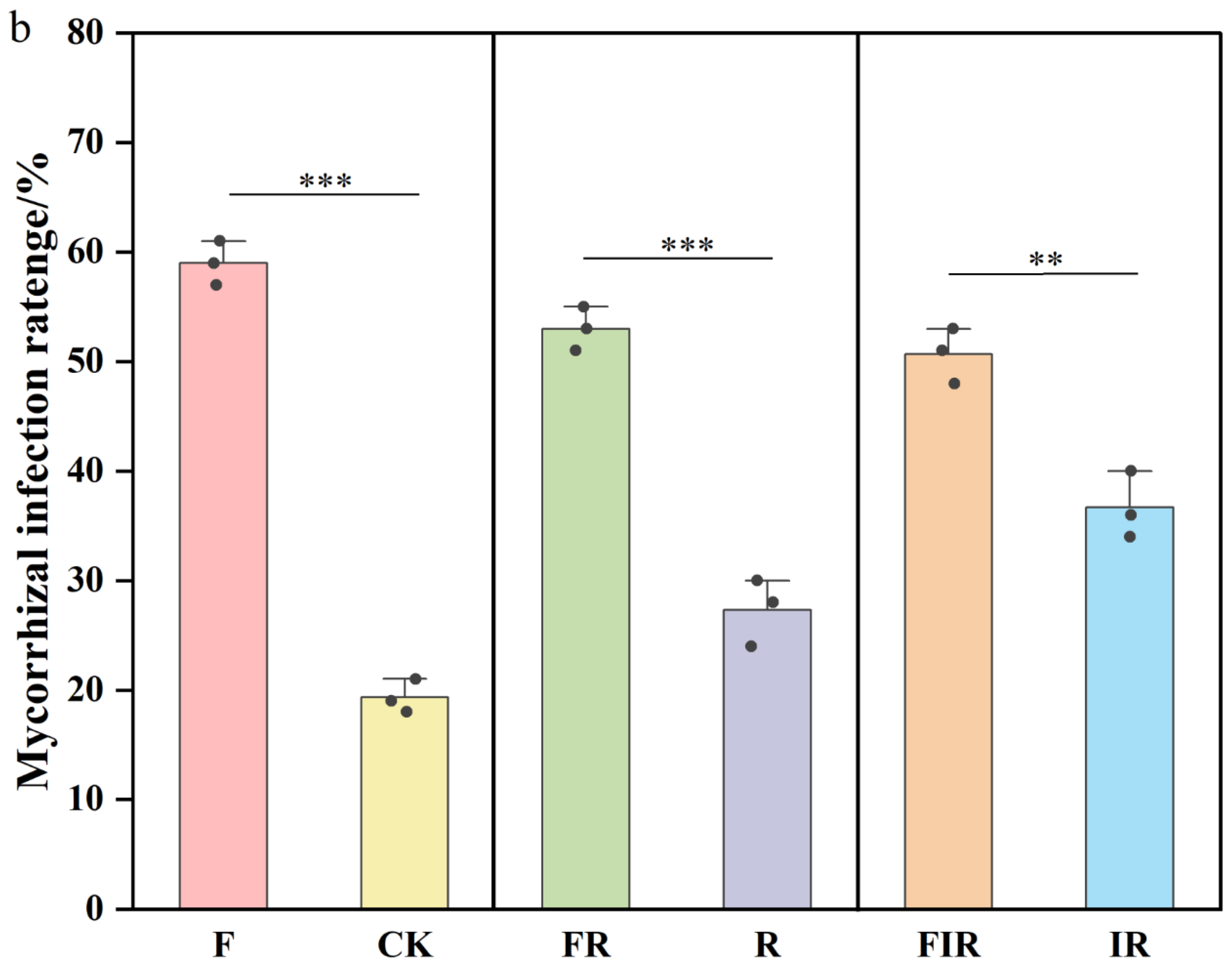
2.4. Determination of Native Mycorrhizal Colonization Rate
2.5. DNA Extraction and PCR Amplification
2.6. Sequencing Data Processing
2.7. Data Analysis
3. Results
3.1. Increased Yield and Soil Nutrient Contents Due to Organic Fertilizer
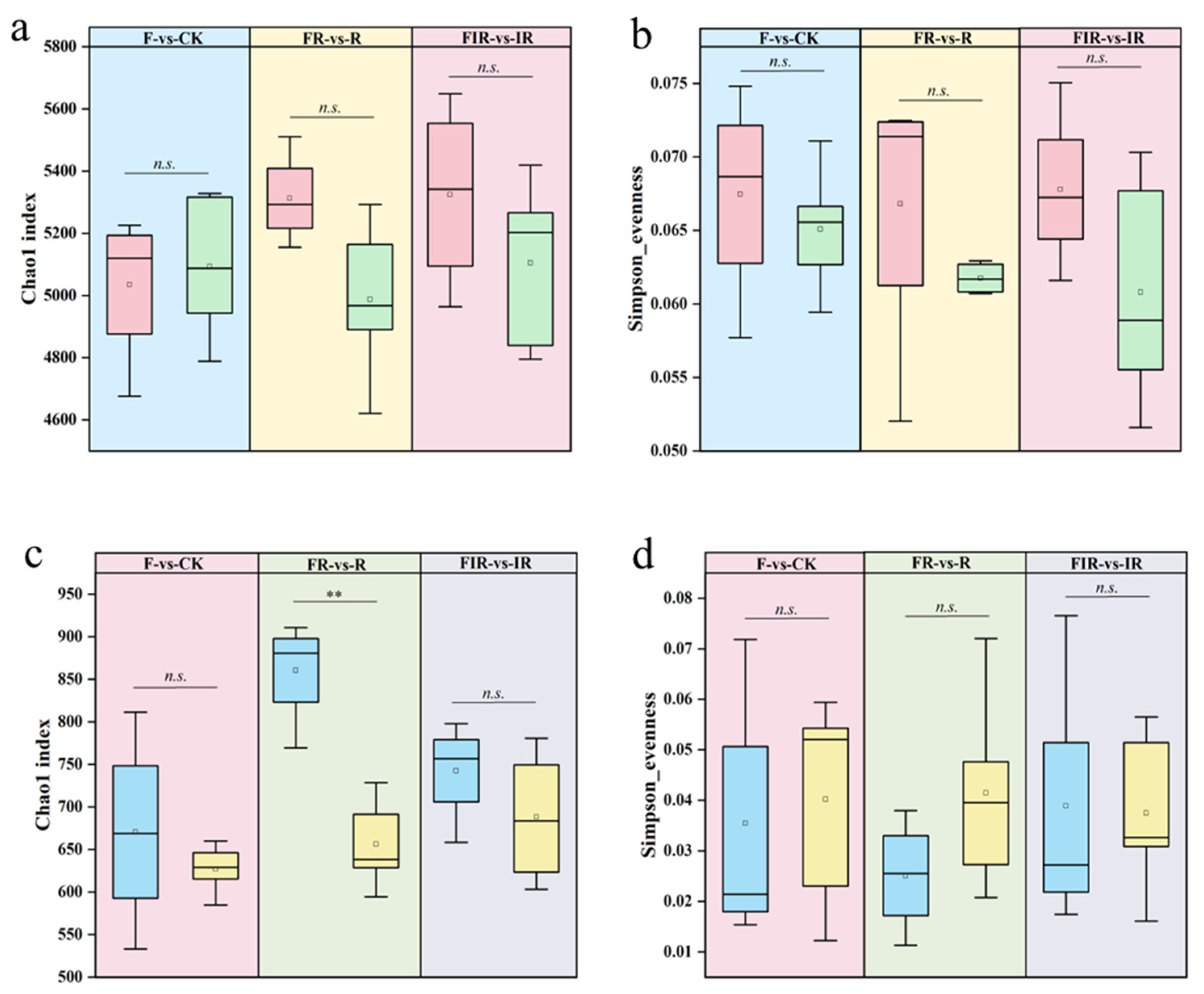
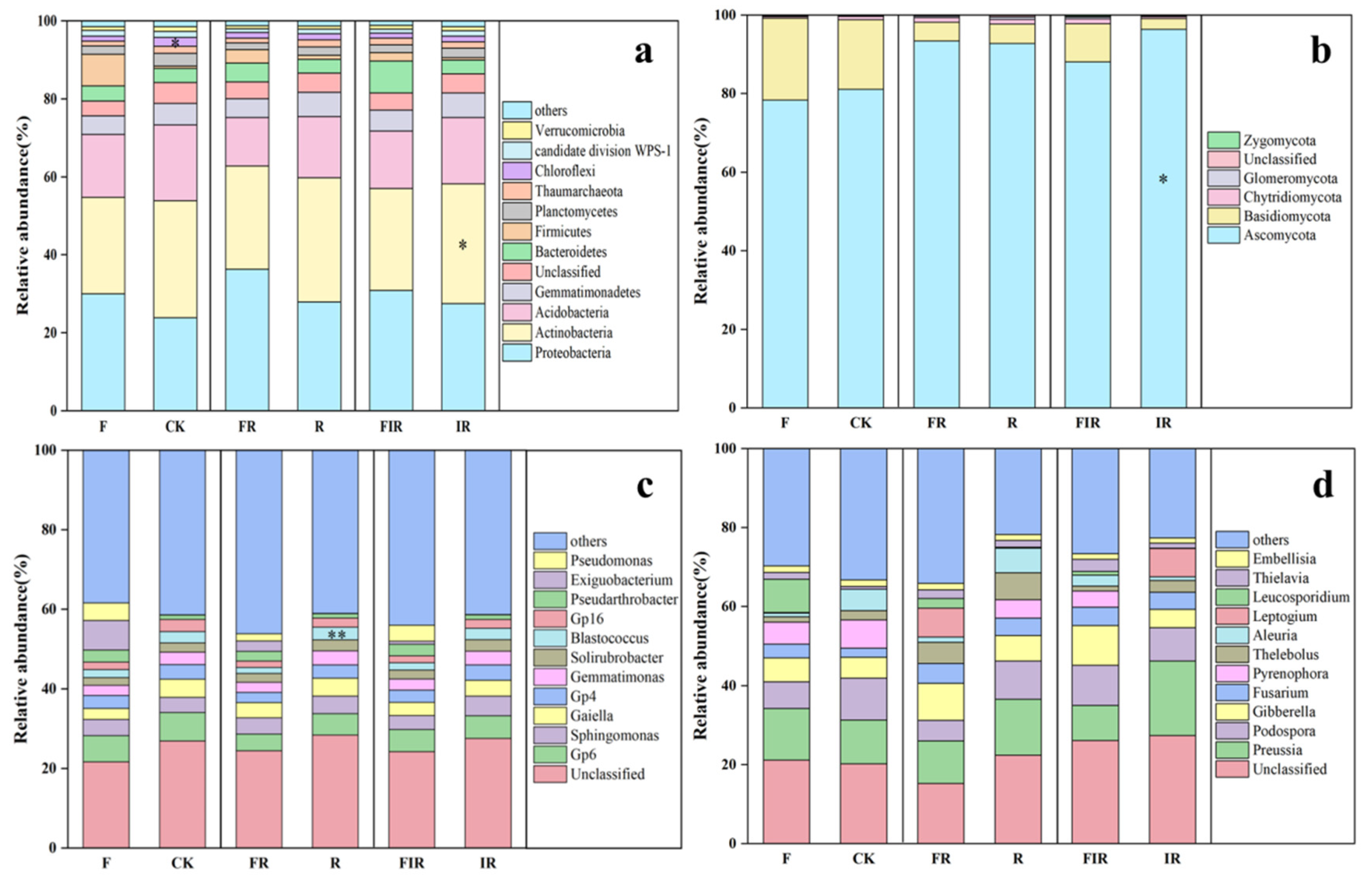
3.2. Organic Fertilizer Application Influenced Soil Microbial Diversity and Species Composition
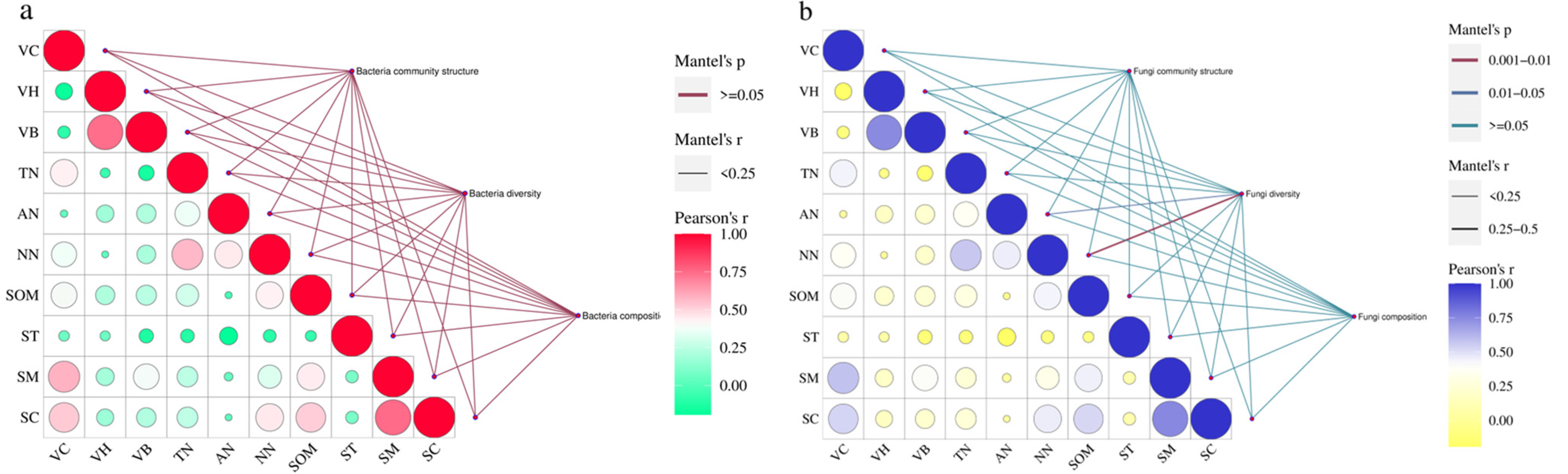
3.3. Correlation between Environmental Factors and Soil Microbial Community Structure
3.4. Increased Native Mycorrhizal Colonization Rate Due to Organic Fertilizer
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Su, X.; Han, W.; Liu, G.; Zhang, Y.; Lu, H. Substantial gaps between the protection of biodiversity hotspots in alpine grasslands and the effectiveness of protected areas on the Qinghai-Tibetan Plateau, China. Agric. Ecosyst. Environ. 2019, 278, 15–23. [Google Scholar] [CrossRef]
- Yang, T.; Adams, J.M.; Shi, Y.; He, J.; Jing, X.; Chen, L.; Tedersoo, L.; Chu, H. Soil fungal diversity in natural grasslands of the Tibetan Plateau: Associations with plant diversity and productivity. New Phytol. 2017, 215, 756–765. [Google Scholar] [CrossRef]
- Zhao, X.; Zhao, L.; Li, Q.; Chen, H.; Zhou, H.; Xu, S.; Dong, Q.; Wu, G.; He, Y. Using balance of seasonal herbage supply and demand to inform sustainable grassland management on the Qinghai-Tibetan Plateau. Front. Agric. Sci. Eng. 2018, 5, 1–8. [Google Scholar] [CrossRef]
- Liu, X.; Huang, X.; Qin, W.; Li, X.; Ma, Z.; Shi, H.; Li, L.; Li, C. Effects of establishing cultivated grassland on soil organic carbon fractions in a degraded alpine meadow on the Tibetan Plateau. PeerJ 2022, 10, e14012. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Yu, F. Reduced compensatory effects explain the nitrogen-mediated reduction in stability of an alpine meadow on the Tibetan Plateau. New Phytol. 2015, 207, 70–77. [Google Scholar] [CrossRef]
- Song, X.; Zhao, C.; Wang, X.; Li, J. Study of nitrate leaching and nitrogen fate under intensive vegetable production pattern in northern China. Comptes Rendus Biol. 2008, 332, 385–392. [Google Scholar] [CrossRef]
- Song, W.; Shu, A.; Liu, J.; Shi, W.; Li, M.; Zhang, W.; Li, Z.; Liu, G.; Yuan, F.; Zhang, S.; et al. Effects of long-term fertilization with different substitution ratios of organic fertilizer on paddy soil. Pedosphere 2022, 32, 637–648. [Google Scholar] [CrossRef]
- Vedpathak, M.; Chavan, B. Impact of organic and chemical fertilizers on grass species composition and biomass. Int. J. Res. Eng. Appl. Sci. 2016, 6, 101–109. [Google Scholar]
- Anna, D.; Josef, H.; Zuzana, H.; Pavel, N. Relationships between long-term fertilization management and forage nutritive value in grasslands. Agric. Ecosyst. Environ. 2019, 279, 139–148. [Google Scholar]
- Chen, Y.; Jiang, Z.; Wu, D.; Wang, H.; Li, J.; Bi, M.; Zhang, Y. Development of a novel bio-organic fertilizer for the removal of atrazine in soil. J. Environ. Manag. 2019, 233, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Li, J.S.; Shao, X.Q.; Huang, D.; Shang, J.Y.; Liu, K.S.; Zhang, Q.; Yang, X.M.; Li, H.; He, Y.X. The addition of organic carbon and nitrogen accelerates the restoration of soil system of degraded alpine grassland in Qinghai-Tibet Plateau. Ecol. Eng. 2020, 158, 106084. [Google Scholar] [CrossRef]
- Wieslaw, G.; Grzegorz, Z.; Jan, R. Permanent Grassland Restoration Techniques-An Overview. Agric. Eng. 2016, 20, 51–58. [Google Scholar]
- Finn, J.A.; Kirwan, L.; Connolly, J.; Sebastià, M.T.; Helgadottir, A.; Baadshaug, O.H.; Bélanger, G.; Black, A.; Brophy, C.; Collins, R.P.; et al. Ecosystem function enhanced by combining four functional types of plant species in intensively managed grassland mixtures: A 3-year continental-scale field experiment. J. Appl. Ecol. 2013, 50, 365–375. [Google Scholar] [CrossRef]
- Tian, Y.; Sun, W.; Song, M.; Zhao, Y.; Wen, S.; Cui, Y.; Li, X.; Xu, X. Effects of grasslegume mixture on plant production and inorganic nitrogen acquisition. Rhizosphere 2021, 20, 100447. [Google Scholar] [CrossRef]
- Perret, X.; Steahelin, C.; Broughton, W. Molecular basis of symbiotic promiscuity. Microbiol. Mol. Biol. Rev. 2000, 64, 180–201. [Google Scholar] [CrossRef]
- Mclennon, E.; Solomon, J.K.Q.; Davison, J. Grass-Legume Forage Systems Effect on Phosphorus Removal from a Grassland Historically Irrigated with Reclaimed Wastewater. Sustainability 2020, 12, 2256. [Google Scholar] [CrossRef]
- Suter, M.; Connolly, J.; Finn, J.A.; Loges, R.; Kirwan, L.; Sebasti`a, M.-T.; Lüscher, A. Nitrogen yield advantage from grass-legume mixtures is robust over a wide range of legume proportions and environmental conditions. Glob. Change Biol. 2015, 21, 2424–2438. [Google Scholar] [CrossRef] [PubMed]
- Sierra, J.; Nygren, P. Transfer of N fixed by a legume tree to the associated grass in a tropical silvopastoral system. Soil Biol. Biochem. 2006, 38, 1893–1903. [Google Scholar] [CrossRef]
- Sierra, J.; Desfontaines, L. Role of root exudates and root turnover in the below-ground N transfer from Canavalia ensiformis (jackbean) to the associated Musa acuminata (banana). Crop Pasture Sci. 2009, 60, 6–10. [Google Scholar] [CrossRef]
- Gasser, H.; Guy, M.; Sikora, L. Efficiency of Rhizobium meliloti strains and their effects on alfalfa cultivars. Can. J. Plant Sci. 1972, 52, 441–448. [Google Scholar] [CrossRef]
- Allison, S.; Lu, Y.; Weihe, C.; Goulden, M.; Martiny, A.; Treseder, K.; Martiny, J. Microbial abundance and composition influence litter decomposition response to environmental change. Ecology 2013, 94, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Comerford, N.; Franzluebbers, A.; Stromberger, M.; Morris, L.; Markewitz, D.; Moore, R. Assessment and evaluation of soil ecosystem services. Soil Horiz. 2013, 54, 1–14. [Google Scholar] [CrossRef]
- Lau, J. Aboveground-belowground linkages: Biotic interactions, ecosystem processes, and global change. Q. Rev. Biol. 2011, 86, 340. [Google Scholar] [CrossRef]
- Ji, L.; Si, H.; He, J.; Fan, L.; Li, L. The shifts of maize soil microbial community and networks are related to soil properties under different organic fertilizers. Rhizosphere 2021, 19, 100388. [Google Scholar] [CrossRef]
- Bei, S.; Zhang, Y.; Li, T.; Christie, P.; Li, X.; Zhang, J. Response of the soil microbial community to different fertilizer inputs in a wheat-maize rotation on a calcareous soil. Agric. Ecosyst. Environ. 2018, 260, 58–69. [Google Scholar] [CrossRef]
- Kim, S.J.; Kim, J.H.; Kim, J.K.; Chang, K.J.; Choi, J.N. Comparative Study of Infection Effects with AMF (Arbuscular-Mycorrhizal Fungi) Isolated from Upland Plants. Korean J. Plant Resour. 2014, 27, 194–201. [Google Scholar] [CrossRef]
- Yan, H.; Gu, S.; Li, S.; Shen, W.; Zhou, X.; Yu, H.; Ma, K.; Zhao, Y.; Wang, Y.; Zheng, H.; et al. Grass-legume mixtures enhance forage production via the bacterial community. Agric. Ecosyst. Environ. 2022, 338, 108087. [Google Scholar] [CrossRef]
- Walkley, A.; Black, C.A. An examination of the digestion method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Strickland, T.; Sollins, P. Improved method for separating light-and heavy-fraction organic material from soil. Soil Sci. Soc. Am. J. 1987, 51, 1390. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, G.; Yu, H.; Du, X.; He, Q.; Yao, S.; Zhao, L.; Huang, C.; Wen, X.; Deng, Y. Meadow degradation increases spatial turnover rates of the fungal community through both niche selection and dispersal limitation. Sci. Total Environ. 2021, 798, 149362. [Google Scholar] [CrossRef] [PubMed]
- Blažková, A.; Jansa, J.; Püschel, D.; Vosátka, M.; Janoušková, M. Is mycorrhiza functioning influenced by the quantitative composition of the mycorrhizal fungal community? Soil Biol. Biochem. 2021, 157, 108249. [Google Scholar] [CrossRef]
- Kowal, J.; Arrigoni, E.; Serra, J.; Bidartondo, M. Prevalence and phenology of fine root endophyte colonization across populations of Lycopodiella inundata. Mycorrhiza 2020, 30, 577–587. [Google Scholar] [CrossRef]
- Feng, K.; Zhang, Z.; Cai, W.; Liu, W.; Xu, M.; Yin, H.; Wang, A.; He, Z.; Deng, Y. Biodiversity and species competition regulate the resilience of microbial biofilm community. Mol. Ecol. 2017, 26, 6170–6182. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, R.; Gao, J.; Wang, X.; Fan, F.; Ma, X.; Yin, H.; Zhang, C.; Feng, K.; Deng, Y. Thirty-one years of rice-rice-green manure rotations shape the rhizosphere microbial community and enrich beneficial bacteria. Soil Biol. Biochem. 2017, 104, 208–217. [Google Scholar] [CrossRef]
- Magoc, T.; Salzberg, S. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y. Btrim: A fast, lightweight adapter and quality trimming program for nextgeneration sequencing technologies. Genomics 2011, 98, 152–153. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Han, J.Q.; Dong, Y.Y.; Zhang, M. Chemical fertilizer reduction with organic fertilizer effectively improve soil fertility and microbial community from newly cultivated land in the Loess Plateau of China. Appl. Soil Ecol. 2021, 165, 103966. [Google Scholar] [CrossRef]
- Pan, H.; Chen, M.M.; Feng, H.J.; Wei, M.; Song, F.P.; Lou, Y.H.; Cui, X.M.; Wang, H.; Zhuge, Y.P. Organic and inorganic fertilizers respectively drive bacterial and fungal community compositions in a fluvo-aquic soil in northern China. Soil Tillage Res. 2020, 198, 104540. [Google Scholar] [CrossRef]
- Cai, A.D.; Xu, M.G.; Wang, B.R.; Zhang, W.J.; Liang, G.P.; Hou, E.Q.; Luo, Y.Q. Manure acts as a better fertilizer for increasing crop yields than synthetic fertilizer does by improving soil fertility. Soil Tillage Res. 2019, 189, 168–175. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, N.; Wu, L.; Xu, M.; Bingham, I.J.; Li, Z. Effects of enhancing soil organic carbon sequestration in the topsoil by fertilization on crop productivity and stability: Evidence from long-term experiments with wheat-maize cropping systems in China. Sci. Total Environ. 2016, 562, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Sabir, M.; Shahzadi, F.; Ali, F.; Shakeela, Q.; Niaz, Z.; Ahmed, S. Comparative Effect of Fertilization Practices on Soil Microbial Diversity and Activity: An Overview. Curr. Microbiol. 2021, 78, 3644–3655. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Tian, Y.; He, M.; Zheng, Y.; Lyu, Q.; Xie, R.; Ma, Y.; Deng, L.; Yi, S. Effects of Chemical Fertilizer Combined with Organic Fertilizer Application on Soil Properties, Citrus Growth Physiology, and Yield. Agriculture 2021, 11, 1207. [Google Scholar] [CrossRef]
- Yates, R.; Steel, E.; Poole, C.; Harrison, R.; Edwards, T.; Hackney, B.; Stagg, G.; Howieson, J. Optimizing the growth of forage and grain legumes on low pH soils through the application of superior Rhizobium leguminosarum biovar viciae strains. Grass Forage Sci. 2021, 76, 44–56. [Google Scholar] [CrossRef]
- KhHM, I.; Fadni, O. Effect of organic fertilizers application on growth, yield and quality of tomatoes in north kordofan (sandy soil) western Sudan. Greener J. Agric. Sci. 2013, 3, 299–304. [Google Scholar]
- Bittman, S.; Forge, T.A.; Kowalenko, C.G. Responses of the bacterial and fungal biomass in a grassland soil to multi-year applications of dairy manure slurry and fertilizer. Soil Biol. Biochem. 2005, 37, 613–623. [Google Scholar] [CrossRef]
- Guo, J.; Li, J.; Liu, K.; Tang, S.; Zhai, X.; Wang, K. Analysis of soil microbial dynamics at a cropland-grassland interface in an agro-pastoral zone in a temperate steppe in northern China. Catena 2018, 170, 257–265. [Google Scholar] [CrossRef]
- Fan, D.; Kong, W.; Wang, F.; Yue, L.; Li, X. Fencing decreases microbial diversity but increases abundance in grassland soils on the tibetan plateau. Land Degrad. Dev. 2020, 31, 2577–2590. [Google Scholar] [CrossRef]
- Silva, L.S.; Laroca, J.V.D.; Coelho, A.P.; Gonsalves, E.C.; Gomes, R.P.; Pacheco, L.P.; Carvalho, P.C.D.; Pires, G.C.; Oliveira, R.L.; de Souza, J.M.A.; et al. Does grass-legume intercropping change soil quality and grain yield in integrated crop-livestock systems? Appl. Soil Ecol. 2021, 170, 104257. [Google Scholar] [CrossRef]
- Chaudhry, V.; Rehman, A.; Mishra, A.; Chauhan, P.S.; Nautiyal, C.S. Changes in bacterial community structure of agricultural land due to long-term organic and chemical amendments. Microb. Ecol. 2012, 64, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Doane, T.A.; Morris, J.; Horwath, W.R. The effect of rice straw on the priming of soil organic matter and methane production in peat soils. Soil Biol. Biochem. 2015, 81, 98–107. [Google Scholar] [CrossRef]
- Guo, J.; Fan, J.; Zhang, F.; Yan, S.; Zheng, J.; Wu, Y.; Li, J.; Wang, Y.; Sun, X.; Liu, X. Blending urea and slow-release nitrogen fertilizer increases dryland maize yield and nitrogen use efficiency while mitigating ammonia volatilization. Sci. Total Environ. 2021, 790, 148058. [Google Scholar] [CrossRef]
- Du, Y.; Cui, B.; Zhang, Q.; Wang, Z.; Niu, W. Effects of manure fertilizer on crop yield and soil properties in China: A meta-analysis. Catena 2020, 193, 104617. [Google Scholar] [CrossRef]
- Li, H.; Hu, Z.; Wan, Q.; Mu, B.; Li, G.; Yang, Y. Integrated Application of Inorganic and Organic Fertilizer Enhances Soil Organo-Mineral Associations and Nutrients in Tea Garden Soil. Agronomy 2022, 12, 1330. [Google Scholar] [CrossRef]
- Xu, H.; Chen, C.; Pang, Z.; Zhang, G.; Wu, J.; Kan, H. Short-Term Vegetation Restoration Enhances the Complexity of Soil Fungal Network and Decreased the Complexity of Bacterial Network. J. Fungi 2022, 8, 1122. [Google Scholar] [CrossRef]
- Yelle, D.; Ralph, J.; Lu, F.; Hammel, K. Evidence for cleavage of lignin by a brown rot basidiomycete. Environ. Microbiol. 2008, 10, 1844–1894. [Google Scholar] [CrossRef]
- Deng, X.H.; Zhang, N.; Li, Y.C.; Zhu, C.Z.; Qu, B.Y.; Liu, H.J.; Li, R.; Bai, Y.; Shen, Q.R.; Salles, J.F. Bio-organic soil amendment promotes the suppression of Ralstonia solanacearum by inducing changes in the functionality and composition of rhizosphere bacterial communities. New Phytol. 2022, 235, 1558–1574. [Google Scholar] [CrossRef]
- Zhong, Y.Q.W.; Yan, W.M.; Shangguan, Z.P. Impact of long-term N additions upon coupling between soil microbial community structure and activity, and nutrient-use efficiencies. Soil Biol. Biochem. 2015, 91, 151–159. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, W.J.; Han, Y.Q.; Huang, X.D.; Wang, C.; Ma, K.X.; Kong, M.Y.; Jiang, N.; Pan, J. Microbial diversity of archaeological ruins of Liangzhu City and its correlation with environmental factors. Int. Biodeterior. Biodegrad. 2022, 175, 105501. [Google Scholar] [CrossRef]
- Wan, P.; Peng, H.; Ji, X.L.; Chen, X.L.; Zhou, H.M. Effect of stand age on soil microbial communities of a plantation Ormosia hosiei forest in southern China. Ecol. Inform. 2021, 62, 101282. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Luo, P.Y.; Yang, J.F.; Irfan, M.; Dai, J.; An, N.; Li, N.; Han, X.Y. Responses of Arbuscular Mycorrhizal Fungi Diversity and Community to 41-Year Rotation Fertilization in Brown Soil Region of Northeast China. Front. Microbiol. 2021, 12, 742651. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; An, X.; Shao, Y.; Kang, Y.; Chen, T.; Mei, X.; Dong, C.; Xu, Y.; Shen, Q. Responses of Arbuscular Mycorrhizal Fungi Occurrence to Organic Fertilizer: A meta-analysis of field studies. Plant Soil 2021, 469, 89–105. [Google Scholar] [CrossRef]
- Buil, P.A.; Jansa, J.; Blazkova, A.; Holubik, O.; Duffkova, R.; Rozmos, M.; Puschel, D.; Kotianova, M.; Janouskova, M. Infectivity and symbiotic efficiency of native arbuscular mycorrhizal fungi from high-input arable soils. Plant Soil 2023, 482, 627–645. [Google Scholar] [CrossRef]
- Fornara, D.A.; Flynn, D.; Caruso, T. Improving phosphorus sustainability in intensively managed grasslands: The potential role of arbuscular mycorrhizal fungi. Sci. Total Environ. 2020, 706, 135744. [Google Scholar] [CrossRef]
- Garo, G.; Van Geel, M.; Eshetu, F.; Swennen, R.; Honnay, O.; Vancampenhout, K. Arbuscular mycorrhizal fungi community composition, richness and diversity on enset (Ensete ventricosum (Welw.) Cheesman) in Ethiopia is influenced by manure application intensity in low-input farming systems. Plant Soil 2022, 478, 409–425. [Google Scholar] [CrossRef]
- Konvalinkova, T.; Puschel, D.; Rezacova, V.; Gryndlerova, H.; Jansa, J. Carbon fow from plant to arbuscular mycorrhizal fungi is reduced under phosphorus fertilization. Plant Soil 2017, 419, 319–333. [Google Scholar] [CrossRef]
- Lekberg, Y.; Koide, R.T.; Twomlow, S.J. Efect of agricultural management practices on arbuscular mycorrhizal fungal abundance in low-input cropping systems of southern Africa: A case study from Zimbabwe. Biol. Fertil. Soils 2008, 44, 917–923. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Vegetation Coverage/ % | Total Nitrogen/ mg/kg | Ammonia Nitrogen/ mg/kg | Nitrate Nitrogen/ mg/kg | Organic Matter/ % |
|---|---|---|---|---|---|
| F | 85.0 ± 5.00 a | 1647.42 ± 139.48 ab | 4.82 ± 0.29 a | 42.58 ± 3.40 bc | 4.53 ± 0.44 ab |
| CK | 57.0 ± 13.04 c | 1404.72 ± 233.60 b | 4.57 ± 0.61 a | 42.17 ± 2.52 bc | 3.52 ± 0.42 b |
| FR | 77.0 ± 9.75 ab | 1811.76 ± 287.35 a | 4.36 ± 1.50 a | 53.17 ± 7.22 ab | 4.89 ± 0.42 a |
| R | 68.0 ± 12.55 bc | 1916.68 ± 260.97 a | 6.43 ± 0.77 a | 51.16 ± 1.12 abc | 3.86 ± 1.04 ab |
| FIR | 87.0 ± 4.47 a | 1857.22 ± 196.34 a | 5.59 ± 2.49 a | 59.62 ± 12.41 a | 4.65 ± 0.52 ab |
| IR | 64.0 ± 13.99 bc | 1641.72 ± 129.47 ab | 4.72 ± 0.55 a | 41.03 ± 1.42 c | 3.82 ± 0.93 ab |
| GroupFactors | Bacteria | Fungi | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Df | SumOfSqs | F. Model | R2 | Pr (>F) | Df | SumOfSqs | F. Model | R2 | Pr (>F) | |
| Application of Organic fertilizer | 1 | 0.23561 | 2.5381 | 0.08311 | 0.018 * | 1 | 0.3494 | 1.6509 | 0.05568 | 0.012 * |
| Complementary seeding | 1 | 0.03831 | 0.3835 | 0.01351 | 0.858 | 1 | 0.1330 | 0.6248 | 0.02119 | 0.978 |
| Inoculated with Rhizobium | 1 | 0.05867 | 0.6096 | 0.02069 | 0.718 | 1 | 0.2588 | 1.2158 | 0.04124 | 0.174 |
| Residuals | 26 | 2.50226 | 0.88268 | 26 | 5.5344 | 0.88189 | ||||
| Total | 29 | 2.83484 | 1.00000 | 29 | 6.2756 | 1.00000 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Zheng, K.; Gu, S.; Wang, Y.; Zhou, X.; Yan, H.; Ma, K.; Zhao, Y.; Jin, X.; Lu, G.; et al. Grass–Legume Mixture with Rhizobium Inoculation Enhanced the Restoration Effects of Organic Fertilizer. Microorganisms 2023, 11, 1114. https://doi.org/10.3390/microorganisms11051114
Zhang H, Zheng K, Gu S, Wang Y, Zhou X, Yan H, Ma K, Zhao Y, Jin X, Lu G, et al. Grass–Legume Mixture with Rhizobium Inoculation Enhanced the Restoration Effects of Organic Fertilizer. Microorganisms. 2023; 11(5):1114. https://doi.org/10.3390/microorganisms11051114
Chicago/Turabian StyleZhang, Haijuan, Kaifu Zheng, Songsong Gu, Yingcheng Wang, Xueli Zhou, Huilin Yan, Kun Ma, Yangan Zhao, Xin Jin, Guangxin Lu, and et al. 2023. "Grass–Legume Mixture with Rhizobium Inoculation Enhanced the Restoration Effects of Organic Fertilizer" Microorganisms 11, no. 5: 1114. https://doi.org/10.3390/microorganisms11051114