
Pilot Study on the Prevalence of Entamoeba gingivalis in Austria—Detection of a New Genetic Variant


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling/DNA Extraction
2.2. Primer Design
2.3. PCR/Sequencing
2.4. Sequence Assembly
2.5. Phylogeny
2.6. Statistical Analysis
3. Results
3.1. Participants
3.2. Detection of Entamoeba gingivalis DNA and Association with Self-RGI
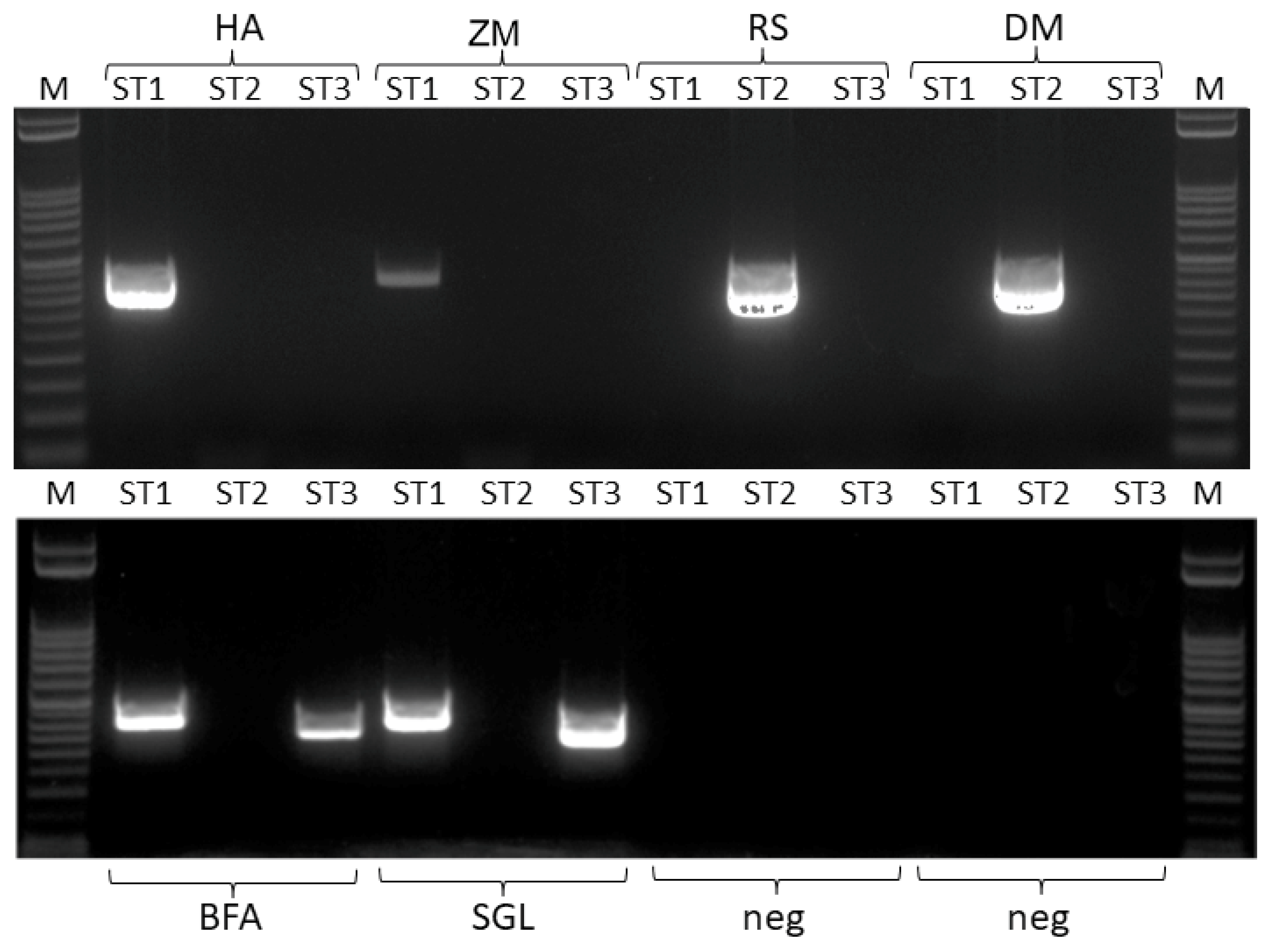
3.3. Sequence Analysis
3.4. Detection of Double Infections
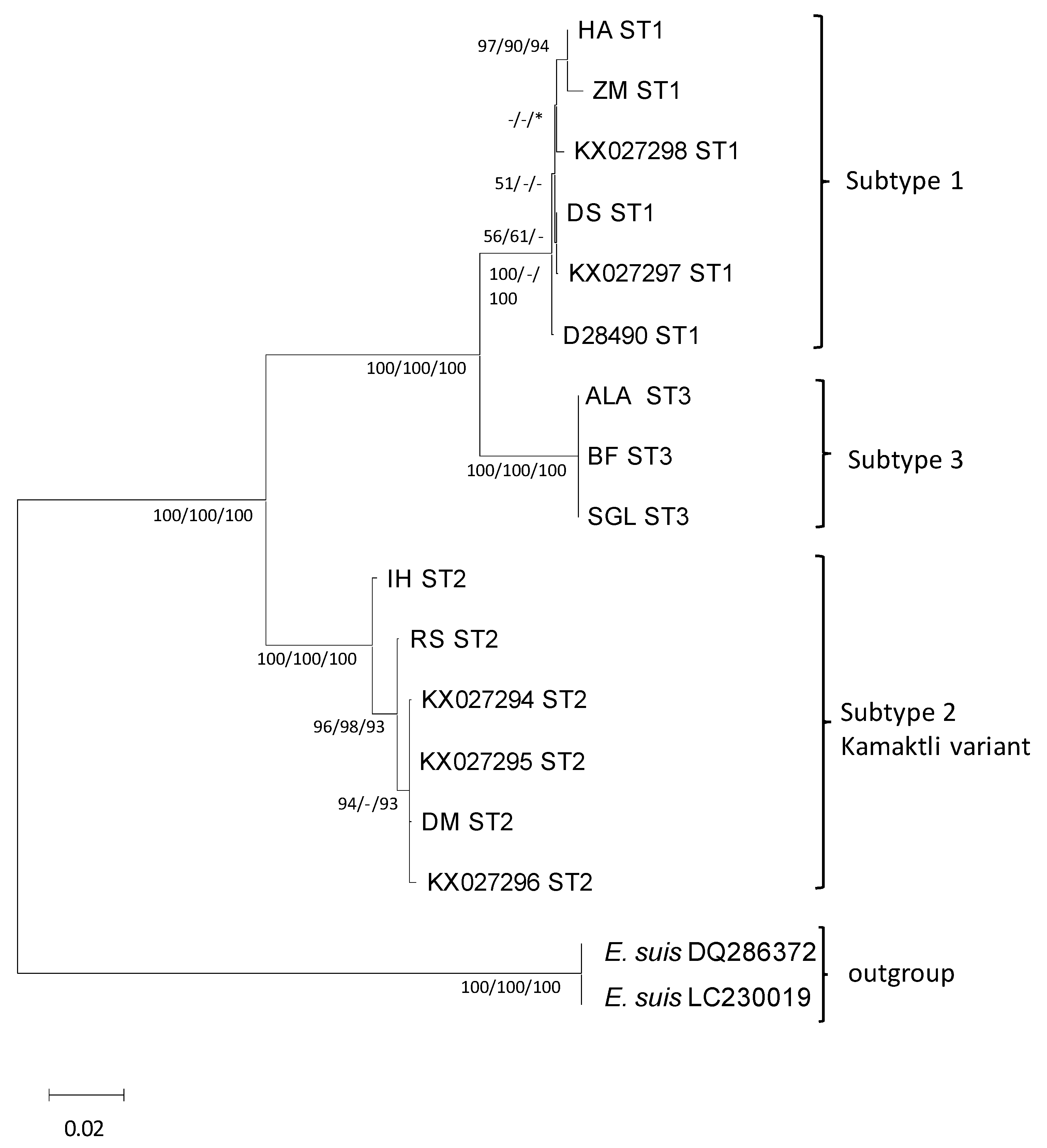
3.5. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Badri, M.; Olfatifar, M.; Abdoli, A.; Houshmand, E.; Zarabadipour, M.; Abadi, P.A.; Johkool, M.G.; Ghorbani, A.; Eslahi, A.V. Current Global Status and the Epidemiology of Entamoeba gingivalis in Humans: A Systematic Review and Meta-analysis. Acta Parasitol. 2021, 66, 1102–1113. [Google Scholar] [CrossRef]
- Randolph, N.K.; McAloney, C.A.; Ossiboff, R.; Hernandez, B.; Cook, M.; Hokamp, J.A. Nasal colonization by Entamoeba gingivalis in a 13-year-old Italian Greyhound. Vet. Clin. Pathol. 2022, 51, 269–272. [Google Scholar] [CrossRef]
- Trim, R.D.; Skinner, M.A.; Farone, M.B.; Dubois, J.D.; Newsome, A.L. Use of PCR to detect Entamoeba gingivalis in diseased gingival pockets and demonstrate its absence in healthy gingival sites. Parasitol. Res. 2011, 109, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Bonner, M.; Amard, V.; Bar-Pinatel, C.; Charpentier, F.; Chatard, J.M.; Desmuyck, Y.; Ihler, S.; Rochet, J.P.; Roux de La Tribouille, V.; Saladin, L.; et al. Detection of the amoeba Entamoeba gingivalis in periodontal pockets. Parasite 2014, 21, 30. [Google Scholar] [CrossRef]
- Bao, X.; Wiehe, R.; Dommisch, H.; Schaefer, A.S. Entamoeba gingivalis Causes Oral Inflammation and Tissue Destruction. J. Dent. Res. 2020, 99, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Bonner, M.; Fresno, M.; Girones, N.; Guillen, N.; Santi-Rocca, J. Reassessing the Role of Entamoeba gingivalis in Periodontitis. Front. Cell Infect. Microbiol. 2018, 8, 379. [Google Scholar] [CrossRef] [PubMed]
- Ralston, K.S.; Solga, M.D.; Mackey-Lawrence, N.M.; Somlata; Bhattacharya, A.; Petri, W.A., Jr. Trogocytosis by Entamoeba histolytica contributes to cell killing and tissue invasion. Nature 2014, 508, 526–530. [Google Scholar] [CrossRef]
- Miller, H.W.; Tam, T.S.Y.; Ralston, K.S. Entamoeba histolytica Develops Resistance to Complement Deposition and Lysis after Acquisition of Human Complement-Regulatory Proteins through Trogocytosis. mBio 2022, 13, e0316321. [Google Scholar] [CrossRef]
- Miller, H.W.; Suleiman, R.L.; Ralston, K.S. Trogocytosis by Entamoeba histolytica Mediates Acquisition and Display of Human Cell Membrane Proteins and Evasion of Lysis by Human Serum. mBio 2019, 10, e00068-19. [Google Scholar] [CrossRef]
- Bao, X.; Weiner, J., 3rd; Meckes, O.; Dommisch, H.; Schaefer, A.S. Entamoeba gingivalis Exerts Severe Pathogenic Effects on the Oral Mucosa. J. Dent. Res. 2021, 100, 771–776. [Google Scholar] [CrossRef]
- Lucht, E.; Evengard, B.; Skott, J.; Pehrson, P.; Nord, C.E. Entamoeba gingivalis in human immunodeficiency virus type 1-infected patients with periodontal disease. Clin. Infect. Dis. 1998, 27, 471–473. [Google Scholar] [CrossRef] [PubMed]
- Cembranelli, S.B.; Souto, F.O.; Ferreira-Paim, K.; Richinho, T.T.; Nunes, P.L.; Nascentes, G.A.; Ferreira, T.B.; Correia, D.; Lages-Silva, E. First evidence of genetic intraspecific variability and occurrence of Entamoeba gingivalis in HIV(+)/AIDS. PLoS ONE 2013, 8, e82864. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.; Kikuta, N.; Hashimoto, T.; Oyaizu, H.; Goto, N. Nucleotide sequence of the SrRNA gene of Entamoeba gingivalis: Applications for construction of a species-specific DNA probe and phylogenetic analysis. Microbiol. Immunol. 1995, 39, 185–192. [Google Scholar] [CrossRef]
- Garcia, G.; Ramos, F.; Martinez-Hernandez, F.; Hernandez, L.; Yanez, J.; Gaytan, P. A new subtype of Entamoeba gingivalis: “E. gingivalis ST2, kamaktli variant”. Parasitol. Res. 2018, 117, 1277–1284. [Google Scholar] [CrossRef]
- Garcia, G.; Ramos, F.; Maldonado, J.; Fernandez, A.; Yanez, J.; Hernandez, L.; Gaytan, P. Prevalence of two Entamoeba gingivalis ST1 and ST2-kamaktli subtypes in the human oral cavity under various conditions. Parasitol. Res. 2018, 117, 2941–2948. [Google Scholar] [CrossRef]
- Zhou, L.; Sriram, R.; Visvesvara, G.S.; Xiao, L. Genetic variations in the internal transcribed spacer and mitochondrial small subunit rRNA gene of Naegleria spp. J. Eukaryot. Microbiol. 2003, 50, 522–526. [Google Scholar] [CrossRef]
- Kohsler, M.; Leitner, B.; Blaschitz, M.; Michel, R.; Aspock, H.; Walochnik, J. ITS1 sequence variabilities correlate with 18S rDNA sequence types in the genus Acanthamoeba (Protozoa: Amoebozoa). Parasitol. Res. 2006, 98, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Hillis, D.M.; Dixon, M.T. Ribosomal DNA: Molecular evolution and phylogenetic inference. Q. Rev. Biol. 1991, 66, 411–453. [Google Scholar] [CrossRef] [PubMed]
- Som, I.; Azam, A.; Bhattacharya, A.; Bhattacharya, S. Inter- and intra-strain variation in the 5.8S ribosomal RNA and internal transcribed spacer sequences of Entamoeba histolytica and comparison with Entamoeba dispar, Entamoeba moshkovskii and Entamoeba invadens. Int. J. Parasitol. 2000, 30, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef]
- Zaffino, M.; Dubar, M.; Debourgogne, A.; Bisson, C.; Machouart, M. Development of a new TaqMan PCR assay for the detection of both Entamoeba gingivalis genotypes. Diagn Microbiol. Infect. Dis. 2019, 95, 114886. [Google Scholar] [CrossRef]
- Deng, Z.L.; Szafranski, S.P.; Jarek, M.; Bhuju, S.; Wagner-Dobler, I. Dysbiosis in chronic periodontitis: Key microbial players and interactions with the human host. Sci. Rep. 2017, 7, 3703. [Google Scholar] [CrossRef] [PubMed]
- Kikuta, N.; Yamamoto, A.; Goto, N. Detection and identification of Entamoeba gingivalis by specific amplification of rRNA gene. Can. J. Microbiol. 1996, 42, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, M.; Jahanimoghadam, F.; Babaei, Z.; Mohammadi, M.A.; Sharifi, F.; Hatami, N.; Danesh, M.; Poureslami, P.; Poureslami, H. Prevalence and Associated-Factors for Entamoeba gingivalis in Adolescents in Southeastern Iran by Culture and PCR, 2017. Iran J. Public Health 2020, 49, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Yaseen, A.; Mahafzah, A.; Dababseh, D.; Taim, D.; Hamdan, A.A.; Al-Fraihat, E.; Hassona, Y.; Sahin, G.O.; Santi-Rocca, J.; Sallam, M. Oral Colonization by Entamoeba gingivalis and Trichomonas tenax: A PCR-Based Study in Health, Gingivitis, and Periodontitis. Front. Cell Infect. Microbiol. 2021, 11, 782805. [Google Scholar] [CrossRef] [PubMed]
- Dover, G.A.; Strachan, T.; Coen, E.S.; Brown, S.D. Molecular drive. Science 1982, 218, 1069. [Google Scholar] [CrossRef]
- Simon, U.K.; Weiss, M. Intragenomic variation of fungal ribosomal genes is higher than previously thought. Mol. Biol. Evol. 2008, 25, 2251–2254. [Google Scholar] [CrossRef]
- Gong, J.; Dong, J.; Liu, X.; Massana, R. Extremely high copy numbers and polymorphisms of the rDNA operon estimated from single cell analysis of oligotrich and peritrich ciliates. Protist 2013, 164, 369–379. [Google Scholar] [CrossRef]
- Weber, A.A.; Pawlowski, J. Wide occurrence of SSU rDNA intragenomic polymorphism in foraminifera and its implications for molecular species identification. Protist 2014, 165, 645–661. [Google Scholar] [CrossRef] [PubMed]
- Windsor, J.J.; Macfarlane, L.; Clark, C.G. Internal transcribed spacer dimorphism and diversity in Dientamoeba fragilis. J. Eukaryot. Microbiol. 2006, 53, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Silberman, J.D.; Clark, C.G.; Diamond, L.S.; Sogin, M.L. Phylogeny of the genera Entamoeba and Endolimax as deduced from small-subunit ribosomal RNA sequences. Mol. Biol. Evol. 1999, 16, 1740–1751. [Google Scholar] [CrossRef]
- Tannich, E.; Horstmann, R.D.; Knobloch, J.; Arnold, H.H. Genomic DNA differences between pathogenic and nonpathogenic Entamoeba histolytica. Proc. Natl. Acad. Sci. USA 1989, 86, 5118–5122. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Lebbad, M.; Victory, E.L.; Verweij, J.J.; Tannich, E.; Alfellani, M.; Legarraga, P.; Clark, C.G. Increased sampling reveals novel lineages of Entamoeba: Consequences of genetic diversity and host specificity for taxonomy and molecular detection. Protist 2011, 162, 525–541. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Ascuna-Durand, K.; Chihi, A.; Belkessa, S.; Kurt, O.; El-Badry, A.; van der Giezen, M.; Clark, C.G. Further insight into the genetic diversity of Entamoeba coli and Entamoeba hartmanni. J. Eukaryot. Microbiol. 2022, 70, e12949. [Google Scholar] [CrossRef]
- Ibanez-Escribano, A.; Nogal-Ruiz, J.J.; Aran, V.J.; Escario, J.A.; Gomez-Barrio, A.; Alderete, J.F. Determination of internal transcribed spacer regions (ITS) in Trichomonas vaginalis isolates and differentiation among Trichomonas species. Parasitol. Int. 2014, 63, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Caccio, S.M.; Beck, R.; Almeida, A.; Bajer, A.; Pozio, E. Identification of Giardia species and Giardia duodenalis assemblages by sequence analysis of the 5.8S rDNA gene and internal transcribed spacers. Parasitology 2010, 137, 919–925. [Google Scholar] [CrossRef]
- De Almeida, M.E.; Steurer, F.J.; Koru, O.; Herwaldt, B.L.; Pieniazek, N.J.; da Silva, A.J. Identification of Leishmania spp. by molecular amplification and DNA sequencing analysis of a fragment of rRNA internal transcribed spacer 2. J. Clin. Microbiol. 2011, 49, 3143–3149. [Google Scholar] [CrossRef]
- Paul, J.; Bhattacharya, A.; Bhattacharya, S. Close sequence identity between ribosomal DNA episomes of the non-pathogenic Entamoeba dispar and pathogenic Entamoeba histolytica. J. Biosci. 2002, 27, 619–627. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Primer | Primer Sequence (5′-3′) | Amplicon Length (bp) | Average Tm (°C) |
|---|---|---|---|
| EGF1fw | CTGATGTTAAAGATTAAGCCATGC | ≈590 | 60.3 °C |
| EGF1rev | CGAGCCTTTTAATCACAACAAC | 58.4 °C | |
| EGF2fw | GTTGTTGTGATTAAAAGGCTCG | ≈507 | 58.4 °C |
| EGF2rev | GAAGTTCATACACTCAAGATTTCTC | 60.9 °C | |
| EGF3fw | GAGAAATCTTGAGTGTATGAACTTC | ≈470 | 60.9 °C |
| EGF3rev | CCAAGATGTCTAAGGGCATCAC | 59.5 °C | |
| EGF4fw | GTGATGCCCTTAGACATCTTG G | ≈360 | 59.5 °C |
| EGF4rev | CTCTAAATAAGGAGGTTCACATC | 59.2 °C | |
| EGITSfw | GATGTGAACCTCCTTATTTAGAG | ≈500 | 59.2 °C |
| EGITSrev | GATATGCTTAAGTTAAGAGAGTCAT | 59.2 °C | |
| ST1fw | GGAGTAAAAAGAAACAGTAGTAAG | ≈400 | 57.6 °C |
| ST1rev | CCAATTACTCACATTACAACAATC | 58.3 °C | |
| ST2fw | CTCAACGAAGACAATAGAGAAG | ≈420 | 58.4 °C |
| EGF2rev | GAAGTTCATACACTCAAGATTTCTC | 60.9 °C | |
| ST3fw | CTCTACGTAACTTGTTACAAGAGAGG | ≈330 | 64.6 °C |
| ST3rev | TAATTATCTCCATTTCTCTTCAAAATG | 59.2 °C |
| Total | Self-RGI 1 | Healthy | Age Range | Age Ø + SD | |
|---|---|---|---|---|---|
| Total | 59 | 18 | 41 | 21–81 | 44.5 ± 15 |
| Women | 30 | 11 | 19 | 21–79 | 43 ± 15 |
| Men | 29 | 7 | 22 | 22–81 | 46 ± 16 |
| Total | Positive | Negative | Self- | Pos./Self- | Neg./Self- | No GI 1 | Pos./H 2 | Neg./H 2 | |
|---|---|---|---|---|---|---|---|---|---|
| No. (%) | No. (%) | RGI 1 | RGI 1 No. (%) | RGI 1 No. (%) | No. (%) | No. (%) | |||
| Total | 59 | 27 (45.8%) | 32 (54.2%) | 18 | 14 (77%) | 4 (22%) | 41 | 13 (32%) | 28 (68%) |
| men | 29 | 10 (34.5%) | 19 (65.5%) | 7 | 6 (86%) | 1 (14%) | 22 | 4 (18%) | 18 (82%) |
| women | 30 | 17 (56.7%) | 13 (43.3%) | 11 | 8 (73%) | 3 (27%) | 19 | 9 (47%) | 10 (53%) |
| Age up to 35 | 20 | 9 (45.0%) | 11 (55.0%) | 5 | 5 (100%) | 0 | 15 | 4 (27%) | 11 (73%) |
| Age 35–50 | 21 | 11 (52.4%) | 10 (47.6%) | 6 | 6 (100%) | 0 | 15 | 5 (33%) | 10 (67%) |
| Age from 50 | 18 | 7 (38.9%) | 11 (61.1%) | 7 | 3 (43%) | 4 (57%) | 11 | 4 (36%) | 7 (64%) |
| Entamoeba ST | Comparison ST | Identity Including Gaps (%) | |||||
|---|---|---|---|---|---|---|---|
| 18S | EGI | 18S-28S | ITS1 | 5.8S | ITS2 | ||
| ST1 | ST1 | 98–99 | 93–100 | 97–99 | 88–100 | 99–100 | 90–100 |
| ST1 | ST2 | 88–89 | 78–81 | 86–87 | 61–69 | 91–92 | 66–74 |
| ST1 | ST3 | 94–95 | 85–86 | 92–93 | 65–68 | 97–98 | 80–86 |
| ST2 | ST2 | 98–99 | 92–100 | 96–99 | 84–100 | 96–100 | 87–100 |
| ST2 | ST3 | 88–89 | 80–81 | 86–87 | 64–68 | 91–94 | 69–72 |
| ST3 | ST3 | 100 | 99–100 | 99–100 | 99–100 | 99–100 | 97–100 |
| ST1–ST3 | E. suis | 84–85 | – | – | – | – | – |
| ST1–ST3 | E. histolytica | 76–77 | 54–58 | 71–72 | 42–52 | 67–69 | 31–36 |
| ST1–ST3 | E. dispar | 75–76 | 53–58 | 70–71 | 42–53 | 66–68 | 31–36 |
| Pos. | Pos. | Pos. | Pos. | Neg. | Total | |
|---|---|---|---|---|---|---|
| No. (%) | ST1 | ST2 | ST1/ST3 | No. (%) | ||
| Total | 27 (46%) | 10 (37%) | 8 (30%) | 9 (33%) | 32 (54%) | 59 |
| Men | 10 (34%) | 5 (50%) | 2 (20%) | 3 (30%) | 19 (66%) | 29 |
| Women | 17 (57%) | 5 (30%) | 6 (35%) | 6 (35%) | 13 (43%) | 30 |
| Age up to 35 | 9 (45%) | 4 (45%) | 3 (33%) | 2 (22%) | 11 (55%) | 20 |
| Age 35–50 | 11 (52%) | 3 (27%) | 2 (18%) | 6 (55%) | 10 (48%) | 21 |
| Age from 50 | 7 (39%) | 3 (43%) | 3 (43%) | 1 (14%) | 11 (61%) | 18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Köhsler, M.; Kniha, E.; Wagner, A.; Walochnik, J. Pilot Study on the Prevalence of Entamoeba gingivalis in Austria—Detection of a New Genetic Variant. Microorganisms 2023, 11, 1094. https://doi.org/10.3390/microorganisms11051094
Köhsler M, Kniha E, Wagner A, Walochnik J. Pilot Study on the Prevalence of Entamoeba gingivalis in Austria—Detection of a New Genetic Variant. Microorganisms. 2023; 11(5):1094. https://doi.org/10.3390/microorganisms11051094
Chicago/Turabian StyleKöhsler, Martina, Edwin Kniha, Angelika Wagner, and Julia Walochnik. 2023. "Pilot Study on the Prevalence of Entamoeba gingivalis in Austria—Detection of a New Genetic Variant" Microorganisms 11, no. 5: 1094. https://doi.org/10.3390/microorganisms11051094