
Physico-Chemical Characterization of an Exocellular Sugars Tolerant Β-Glucosidase from Grape Metschnikowia pulcherrima Isolates

Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Isolates
2.2. Typing by Molecular Techniques
2.3. Qualitative Screening of the β-Glucosidase Activity
2.4. Quantitative β-Glucosidase Assay
2.5. Effect of Sugars and Ethanol on β-Glucosidase Activity
2.6. Effect of Temperature and pH on β-Glucosidase Activity
2.7. Effect of Metal Ions on β-Glucosidase Activity
2.8. Winemaking
2.9. Determination of Terpene Compounds Liberated from Wine Incubated with M. pulcherrima Strains
2.10. Statistical Analyses
3. Results and Discussion
3.1. Molecular Identification of Isolates
3.2. Qualitative and Quantitative Detection of β-Glucosidase
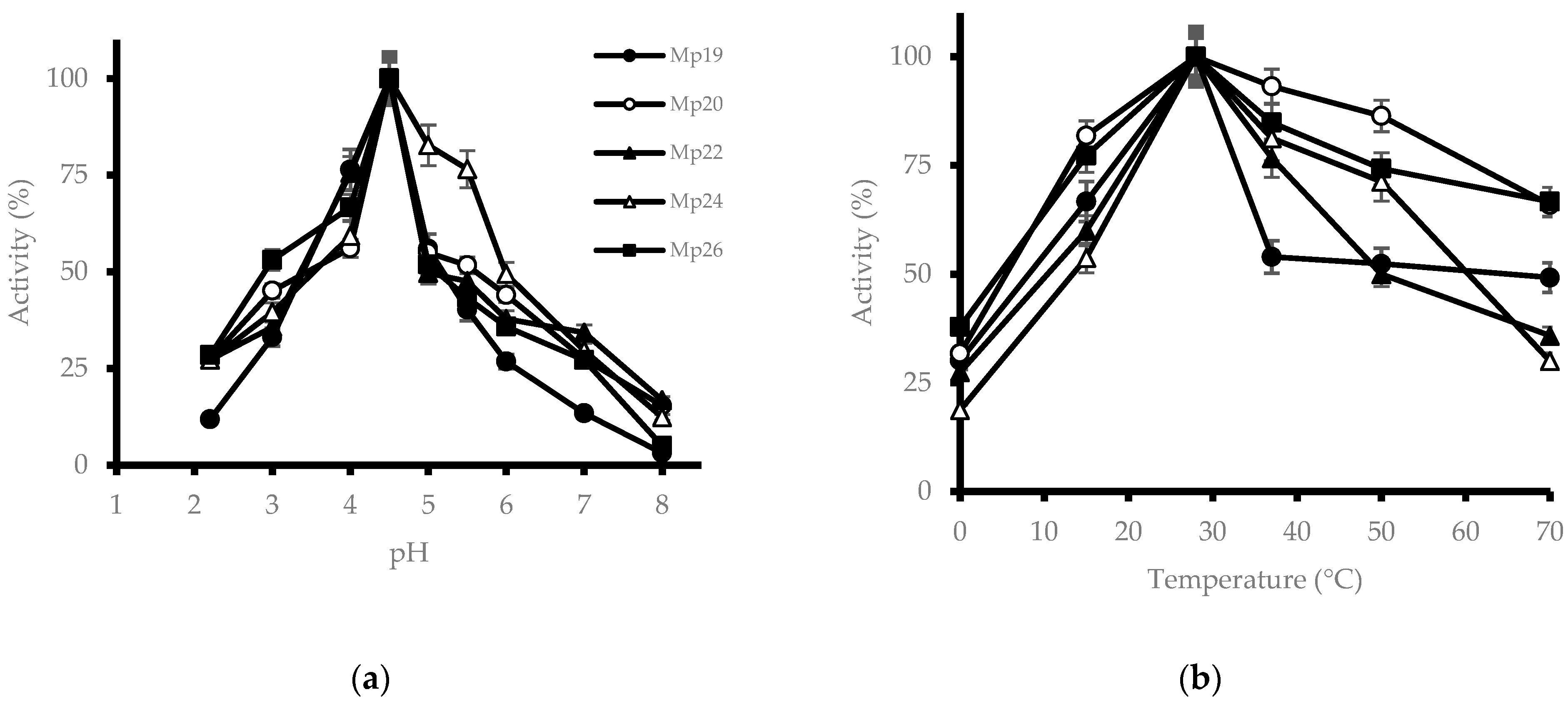
3.3. Effect of pH and Temperature on β-Glucosidase Activity
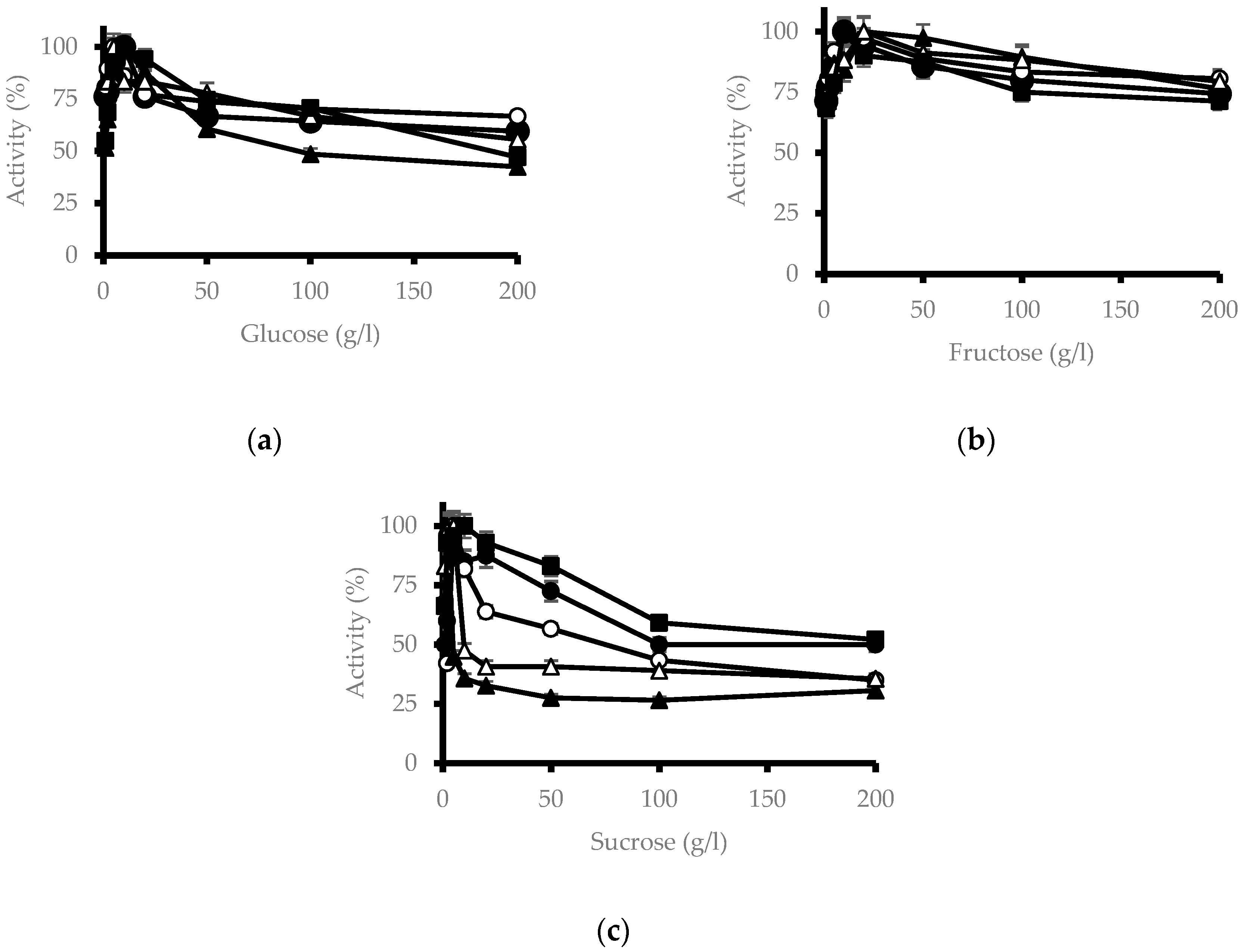
3.4. Effect of Sugars on β-Glucosidase Activity
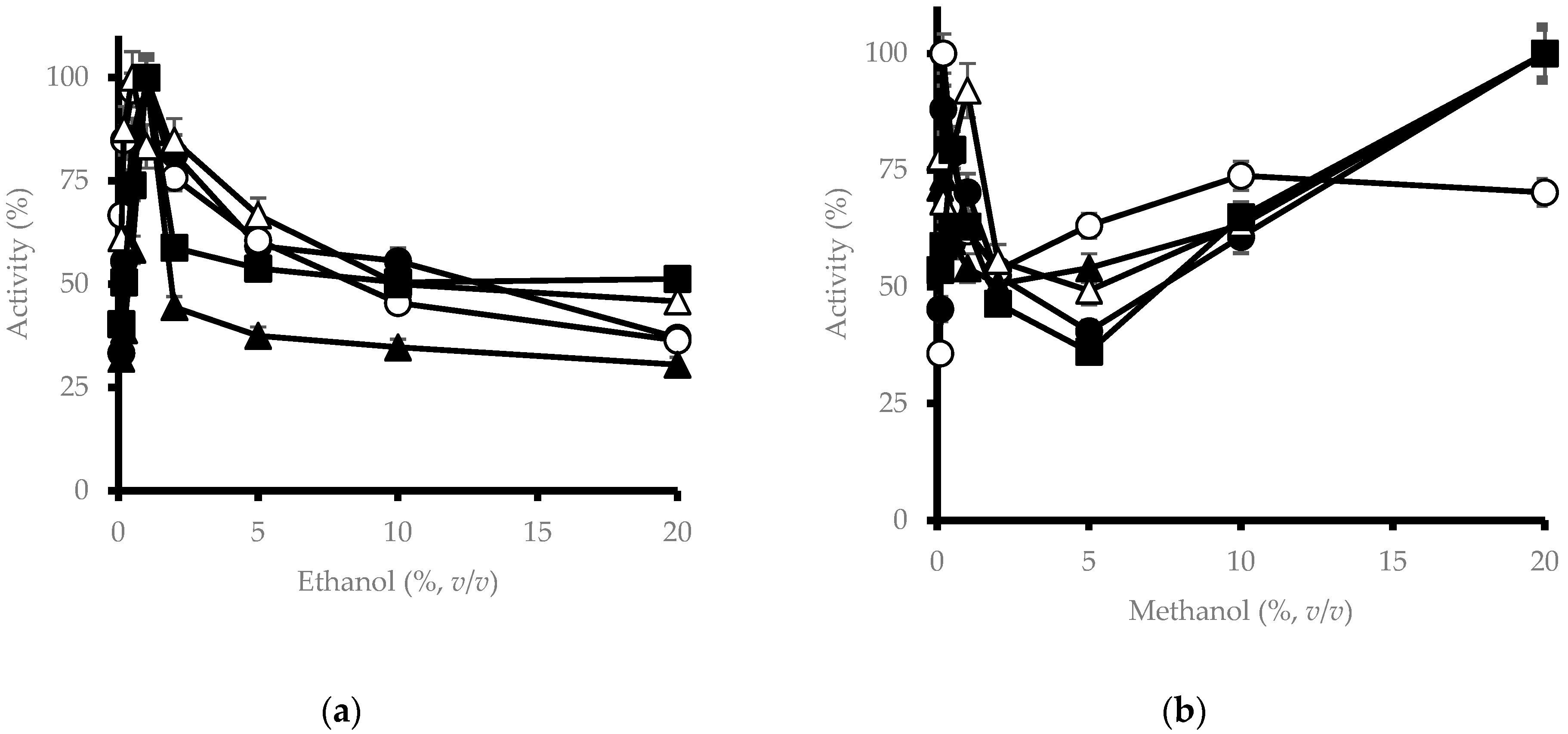
3.5. Effect of Alcohols on β-Glucosidase Activity
3.6. Effect of Chemicals on β-Glucosidase Activity
3.7. Determination of Terpene Compounds Liberated from Wine Treated with Different M. pulcherrima Yeasts
4. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- Romano, P.; Capece, A.; Jespersen, L. Taxonomic and Ecological Diversity of Food and Beverage Yeasts. In Yeasts in Food and Beverages; Querol, A., Fleet, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 13–53. [Google Scholar]
- Pitt, J.I.; Miller, M.W. Sporulation in Candida pulcherrima, Candida reukaufii and Chlamydozyma species—Their relationships with Metschnikowia. Mycologia 1968, 60, 663–685. [Google Scholar] [CrossRef]
- Gao, P.; Peng, S.; Sam, F.E.; Zhu, Y.; Liang, L.; Li, M.; Wang, J. Indigenous non-Saccharomyces yeasts with β-glucosidase activity in sequential fermentation with Saccharomyces cerevisiae: A strategy to improve the volatile composition and sensory characteristics of wines. Front. Microbiol. 2022, 13, 845837. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Loira, I.; Escott, C.; del Fresno, J.M.; Bañuelos, M.A.; Suárez-Lepe, J.A. Applications of Metschnikowia pulcherrima in wine biotechnology. Fermentation 2019, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Jolly, N.P.; Augustyn, O.P.H.; Pretorius, I.S. The role and use of non-Saccharomyces yeasts in wine production. S. Afr. J. Enol. Vitic. 2006, 27, 15–39. [Google Scholar] [CrossRef]
- Barbosa, C.; Lage, P.; Esteves, M.; Chambel, L.; Mendes-Faia, A.; Mendes-Ferreira, A. Molecular and phenotypic characterization of Metschnikowia pulcherrima strains from Douro wine region. Fermentation 2018, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Singhania, R.R.; Patel, A.K.; Sukumaran, R.K.; Larroche, C.; Pandey, A. Role and significance of beta-glucosidases in the hydrolysis of cellulose for bioethanol production. Bioresour. Technol. 2013, 127, 500–507. [Google Scholar] [CrossRef]
- Mateo, J.J.; Maicas, S. Non-Saccharomyces yeasts: An enzymatic unexplored world to be exploited. In Enzymes in Food Biotechnology; Kuddus, M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 433–450. [Google Scholar]
- Liew, K.J.; Lim, L.; Woo, H.Y.; Chan, K.; Shamsir, M.S.; Goh, K.M. Purification and characterization of a novel GH1 beta-glucosidase from Jeotgalibacillus malaysiensis. Int. J. Biol. Macromol. 2018, 115, 1094–1102. [Google Scholar] [CrossRef]
- Méndez-Líter, J.A.; Nieto-Domínguez, M.; de Toro, B.F.; Santana, A.G.; Prieto, A.; Asensio, J.L.; Cañada, F.J.; de Eugenio, L.I.; Martínez, M.J. A glucotolerant β-glucosidase from the fungus Talaromyces amestolkiae and its conversion into a glycosynthase for glycosylation of phenolic compounds. Microb. Cell Fact. 2020, 19, 127. [Google Scholar] [CrossRef]
- Mendes Ferreira, A.; Clímaco, M.C.; Mendes Faia, A. The role of non-Saccharomyces species in releasing glycosidic bound fraction of grape aroma components-a preliminary study. J. Appl. Microbiol. 2001, 91, 67–71. [Google Scholar] [CrossRef]
- Zott, K.; Thibon, C.; Bely, M.; Lonvaud-Funel, A.; Dubourdieu, D.; Masneuf-Pomarede, I. The grape must non-Saccharomyces microbial community: Impact on volatile thiol release. Int. J. Food Microbiol. 2011, 151, 210–215. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie Van Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef] [PubMed]
- López, S.; Mateo, J.J.; Maicas, S. Characterization of Hanseniaspora strains with aroma-enhancing properties in muscat wine. S. Afr. J. Enol. Vitic. 2014, 35, 292–303. [Google Scholar]
- Zhang, W.; Zhuo, X.; Hu, L.; Zhang, X. Effects of Crude β-Glucosidases from Issatchenkia terricola, Pichia kudriavzevii, Metschnikowia pulcherrima on the Flavor Complexity and Characteristics of Wines. Microorganisms 2020, 8, 953. [Google Scholar] [CrossRef]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef]
- Qin, T.; Liao, J.; Zheng, Y.; Zhang, W.; Zhang, X. Oenological Characteristics of Four Non-Saccharomyces Yeast Strains With β-Glycosidase Activity. Front. Microbiol. 2021, 12, 626920. [Google Scholar] [CrossRef]
- Gao, P.; Sam, F.E.; Zhang, B.; Peng, S.; Li, M.; Wang, J. Enzymatic Characterization of Purified β-Glucosidase from Non-Saccharomyces Yeasts and Application on Chardonnay Aging. Foods 2022, 11, 852. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.; Jing, S.; Zhang, H.; Yang, X.; Jin, G.; Tao, Y. Localization, purification, and characterization of a novel β-glucosidase from Hanseniaspora uvarum Yun268. J. Food Sci. 2022, 87, 886–894. [Google Scholar] [CrossRef]
- Hernandez-Guzman, A.; Flores-Martıinez, A.; Ponce-Noyola, P.; Julio, C.; Castro, V. Purification and characterization of an extracellular β-glucosidase from Sporothrix schenckii. FEBS Open Bio 2016, 6, 1067–1077. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Zhang, W.; Qin, T.; Liao, J.; Zhang, X. Effects of Purified β-Glucosidases from Issatchenkia terricola, Pichia kudriavzevii, Metschnikowia pulcherrima on the Flavor Complexity and Typicality of Wines. J. Fungi 2022, 8, 1057. [Google Scholar] [CrossRef]
- Vaz, J.E.; Rabelo, L.; Zaiter, M.A.; Pereira, W.E.S.; Metzker, G.; Boscolo, M.; da Silva, R.; Gomes, E.; da Silva, R.R. Functional properties and potential application of ethanol tolerant β-glucosidases from Pichia ofunaensis and Trichosporon multisporum yeasts. 3 Biotech 2021, 11, 467. [Google Scholar] [CrossRef]
- Zufferey, V.; Murisier, F.; Schultz, H.R. A model analysis of the photosynthetic response of Vitis vinifera L. cvs Riesling and Chasselas leaves in the field: I. Interaction of age, light and temperature. Vitis 2000, 39, 19–26. [Google Scholar]
- Türkel, S.; Ener, B. Isolation and characterization of new Metschnikowia pulcherrima strains as producer of the antimicrobial pigment pulcherrimin. Z. Naturforsch. C Biosci. 2009, 64, 405–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.A.; Yoon, K.H. Characterization of a Paenibacillus woosongensis beta–Xylosidase/alpha-Arabinofuranosidase produced by recombinant Escherichia coli. J. Microbiol. Biotechnol. 2010, 20, 1711–1716. [Google Scholar]
- Zhou, J.; Bao, L.; Chang, L.; Zhou, Y.; Hong, L. Biochemical and kinetic characterization of GH43 β-d-xylosidase/α-larabinofuranosidase and GH30 α–l–arabinofuranosidase/β-dxylosidase from rumen metagenome. J. Ind. Microbiol. Biotechnol. 2012, 39, 143–152. [Google Scholar] [CrossRef] [PubMed]
- González-Pombo, P.; Pérez, G.; Carrau, F.; Guisán, J.M.; Batista-Viera, F.; Brena, B.M. One-step purification and characterization of an intracellular β-glucosidase from Metschnikowia pulcherrima. Biotechnol. Lett. 2008, 30, 1469–1475. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Meng, T.; Li, Z.; Liu, P.; Wang, Y.; He, N.; Liang, D. Characterization of a beta-glucosidase from Bacillus licheniformis and its effect on bioflocculant degradation. AMB Express 2017, 7, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.M.; Lee, J.P.; Baek, S.C.; Kim, S.G.; Jo, Y.D.; Kim, J.; Kim, H. Characterization of two extracellular β-glucosidases produced from the cellulolytic fungus Aspergillus sp. YDJ216 and their potential applications for the hydrolysis of flavone glycosides. Int. J. Biol. Macromol. 2018, 111, 595–603. [Google Scholar]
- Mateo, J.J.; Andreu, L. Characterization of an exocellular ethanol-tolerant β-glucosidase from Quambalaria cyanescens isolates from unripened grapes. Eur. Food Res. Technol. 2020, 246, 2349–2357. [Google Scholar] [CrossRef]
- Sinha, S.K.; Reddy, K.P.; Datta, S. Understanding the glucose tolerance of an archaeon β-glucosidase from Thermococcus sp. Carbohydr. Res. 2019, 486, 107835. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, X.; Yin, Q.; Fang, W.; Fang, Z.; Wang, X.; Zhang, X.; Xiao, Y. A mechanism of glucose tolerance and stimulation of GH1 β-glucosidases. Sci. Rep. 2015, 5, 17296. [Google Scholar] [CrossRef] [Green Version]
- Guo, B.; Amano, Y.; Nozaki, K. Improvements in glucose sensitivity and stability of Trichoderma reesei β-glucosidase using site-directed mutagenesis. PLoS ONE 2016, 11, e0147301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, H.; Xiao, Z.; Yu, K.; Zhang, Q.; Lu, C.; Wang, G.; Wang, Y.; Liang, J.; Huang, W.; Huang, X.; et al. Use of a purified β-glucosidase from coral-associated microorganisms to enhance wine aroma. J. Sci. Food Agric. 2022, 102, 3467–3474. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.; Ruiz, J.; Belda, I.; Benito-Vázquez, I.; Marquina, D.; Calderón, F.; Santos, A.; Benito, S. The genus Metschnikowia in enology. Microorganisms 2020, 8, 1038. [Google Scholar] [CrossRef] [PubMed]
- González-Centeno, M.R.; Rosselló, C.; Simal, S.; Garau, M.C.; López, F.; Femenia, A. Physico-chemical properties of cell wall materials obtained from ten grape varieties and their byproducts: Grape pomaces and stems. Lebensm. Wiss. Technol. 2010, 43, 1580–1586. [Google Scholar] [CrossRef]
- Fallahi, E.; Conway, W.S.; Hickey, K.D.; Sams, C.E. The role of calcium and nitrogen in postharvest quality and disease resistance of apples. HortScience 1997, 32, 831–835. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, L.M.O.; Vici, A.C.; Pinheiro, M.P.; Heinen, P.R.; De Oliveira, A.H.C.; Ward, R.J.; Prade, R.A.; Buckeridge, M.S.; Polizeli, M.L. A highly glucose tolerant ß-glucosidase from Malbranchea pulchella (MpBg3) enables cellulose saccharification. Sci. Rep. 2020, 10, 6998. [Google Scholar] [CrossRef] [Green Version]
- Liao, J.; Zhang, S.; Zhang, X. Effects of Mixed Adding Crude Extracts of β-Glucosidases from Three Different Non-Saccharomyces Yeast Strains on the Quality of Cabernet Sauvignon Wines. J. Fungi 2022, 8, 710. [Google Scholar] [CrossRef]
- Oliveira, J.M.; Oliveira, P.; Baumes, R.L.; Maia, M.O. Volatile and glycosidically bound composition of Loureiro and Alvarinho wines. Food Sci. Technol. Int. 2008, 14, 341–353. [Google Scholar] [CrossRef] [Green Version]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Fernández-González, M.; Di Stefano, R.; Briones, A. Hydrolysis and transformation of terpene glycosides from Muscat must by different yeast species. Food Microbiol. 2003, 20, 35–41. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Isolate | Absorbance |
|---|---|
| Mp19 | 0.253 |
| Mp20 | 0.234 |
| Mp22 | 0.31 |
| Mp24 | 0.27 |
| Mp26 | 0.256 |
| Isolate | ||||||
|---|---|---|---|---|---|---|
| Compound | mM | Mp19 | Mp20 | Mp22 | Mp24 | Mp26 |
| Ca2+ | 2 | 0.211 | 0.222 | 0.224 | 0.220 | 0.189 |
| 10 | 0.502 * | 0.501 * | 0.537 * | 0.475 * | 0.485 * | |
| Na+ | 2 | 0.212 | 0.205 | 0.205 | 0.225 | 0.199 |
| 10 | 0.204 | 0.212 | 0.219 | 0.200 | 0.216 | |
| Mg2+ | 2 | 0.213 | 0.181 | 0.198 | 0.223 | 0.203 |
| 10 | 0.229 | 0.180 | 0.222 | 0.219 | 0.226 | |
| Zn2+ | 2 | 0.205 | 0.190 | 0.221 | 0.206 | 0.205 |
| 10 | 0.211 | 0.210 | 0.248 | 0.197 | 0.237 | |
| Tween 80 | 2 | 0.249 | 0.214 | 0.239 | 0.216 | 0.211 |
| 10 | 0.234 | 0.261 | 0.246 | 0.252 | 0.239 | |
| Hg2+ | 2 | 0.233 | 0.233 | 0.282 | 0.231 | 0.236 |
| Control | M. Pulcherrima Inoculated | |||
|---|---|---|---|---|
| Mp22 | Mp24 | Mp26 | ||
| Oxide A b | 29.7 (1.2) | 35.4 (2.1) | 32.7 (3.2) | 26.9 (3.4) |
| Oxide B c | nd | nd | nd | nd |
| Linalool | 20.0 (0.9) | 50.4 * (3.9) | 46.4 * (3.4) | 39.2 * (5.3) |
| Ho-trienol | 24.0 (3.2) | 55.3 * (5.3) | 39.1 * (4.2) | 34.9 * (0.6) |
| 2-Phenylethanol | 1890.2 (43.4) | 3117.5 * (39.8) | 2817.8 * (26.8) | 2638.5 * (45.6) |
| Oxide C d | nd | nd | nd | nd |
| Oxide D e | nd | nd | nd | nd |
| Terpineol | 53.3 (3.4) | 68.9 * (4.7) | 66.2 * (1.2) | 56.1 (3.9) |
| Nerol | 24.6 (2.8) | 24.8 (1.1) | 22.4 (3.1) | 25.3 (1.2) |
| Geraniol | 59.8 (5.0) | 60.9 (3.7) | 57.9 (1.7) | 58.2 (1.7) |
| Diol 1 f | 43.2 (4.7) | 90.9 * (2.1) | 88.2 * (2.1) | 85.6 * (3.2) |
| 4-Vinylphenol | 63.2 (1.2) | 86.3 * (2.4) | 77.8 * (5.8) | 63.1 (0.9) |
| Endiol g | nd | 51.2 * (2.1) | 50.1 * (3.4) | 33.7 * (4.2) |
| Diol 2 h | 12.0 (0.6) | 11.4 (0.9) | 9.8 (2.6) | 11.1 (0.9) |
| 2-Phenylethyl acetate | 28.0 (4.1) | 61.2 * (7.2) | 33.3 (1.2) | 35.8 (4.7) |
| 2-Methoxy-4-vinylphenol | 89.0 (6.1) | 112.0 * (5.3) | 116.4 * (6.5) | 104.1 (2.9) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mateo, J.J. Physico-Chemical Characterization of an Exocellular Sugars Tolerant Β-Glucosidase from Grape Metschnikowia pulcherrima Isolates. Microorganisms 2023, 11, 964. https://doi.org/10.3390/microorganisms11040964
Mateo JJ. Physico-Chemical Characterization of an Exocellular Sugars Tolerant Β-Glucosidase from Grape Metschnikowia pulcherrima Isolates. Microorganisms. 2023; 11(4):964. https://doi.org/10.3390/microorganisms11040964
Chicago/Turabian StyleMateo, José Juan. 2023. "Physico-Chemical Characterization of an Exocellular Sugars Tolerant Β-Glucosidase from Grape Metschnikowia pulcherrima Isolates" Microorganisms 11, no. 4: 964. https://doi.org/10.3390/microorganisms11040964