
A Review of the Bacterial Phosphoproteomes of Beneficial Microbes

Department of Pharmaceutical Engineering, Hoseo University, Asan-si 31499, Republic of Korea
Microorganisms 2023, 11(4), 931; https://doi.org/10.3390/microorganisms11040931
Submission received: 28 February 2023
/
Revised: 27 March 2023
/
Accepted: 31 March 2023
/
Published: 3 April 2023
(This article belongs to the Special Issue Latest Review Papers in Molecular Microbiology and Immunology 2023)
Abstract
:The number and variety of protein post-translational modifications (PTMs) found and characterized in bacteria over the past ten years have increased dramatically. Compared to eukaryotic proteins, most post-translational protein changes in bacteria affect relatively few proteins because the majority of modified proteins exhibit substoichiometric modification levels, which makes structural and functional analyses challenging. In addition, the number of modified enzymes in bacterial species differs widely, and degrees of proteome modification depend on environmental conditions. Nevertheless, evidence suggests that protein PTMs play essential roles in various cellular processes, including nitrogen metabolism, protein synthesis and turnover, the cell cycle, dormancy, spore germination, sporulation, persistence, and virulence. Additional investigations on protein post-translational changes will undoubtedly close knowledge gaps in bacterial physiology and create new means of treating infectious diseases. Here, we describe the role of the post-translation phosphorylation of major bacterial proteins and review the progress of research on phosphorylated proteins depending on bacterial species.
1. Introduction
Bacteria play vital roles in the environment, animals, and humans and perform many essential ecological functions, such as recycling organic materials and assisting the carbon and nitrogen cycles. In contrast to plant and animal cells, bacteria are frequently subjected to continuous changes in their physical and chemical surroundings [1]. Bacterial metabolism is controlled by intracellular signals and provides the energy required for cellular activity and adaptation to different environments [2]. Bacteria rapidly adapt to various environments through post-translational modifications (PTMs) or the allosteric binding of small molecules that play a key role in metabolism. This review focuses on protein phosphorylation in PTMs. Protein phosphorylation is the most common and well-studied PTM that bacteria use to regulate protein activity and underlies bacterial protein heterogeneity. Previous studies have shown that phosphorylation is utilized more by eukaryotes than prokaryotes. Nonetheless, research efforts have resulted in the discovery of a wealth of bacterial phosphoproteins, despite the low abundances of protein modifications [3,4,5].
2. Bacterial Protein Phosphorylation
Bacterial protein phosphorylation subserves diverse functions in bacteria related to antibiotic resistance, such as DNA replication, metabolism, heat shock response, biofilm formation, spore formation, anti-virulence, and the production of amino acids and antibiotics. Protein homeostasis and novel protein functions can be achieved by phosphorylation, which requires components of complex cellular signal detection and conversion networks. Protein phosphorylation (His, Asp, Ser, Thr, Tyr, and Arg), glycosylation (Arg, Asn, Ser, and Thr), acetylation (Lys), acylation (Lys), lipidation (Cys), oxidation (Met), and thiolation (Cys) are the most common PTMs [6], and protein phosphorylation is one of the best understood. Amino acid residue phosphorylation can control the activity of proteins by causing structural changes in active sites and modulating protein–protein interactions. For example, in bacteria, protein phosphorylation and dephosphorylation of various amino acids provide a variety of chemical characteristics [7], stabilities, and functionalities [5]. Furthermore, protein phosphorylation plays essential regulatory roles in the cell cycle, receptor-mediated signal transduction, differentiation, proliferation, transformation, and metabolism. Two types of protein phosphorylation systems are most common in bacteria: the so-called two-component systems (TCSs), which include bacterial protein kinases, and the protein phosphorylation system, which affects serine, threonine, and tyrosine side chains.
2.1. Two-Component Systems (TCSs)
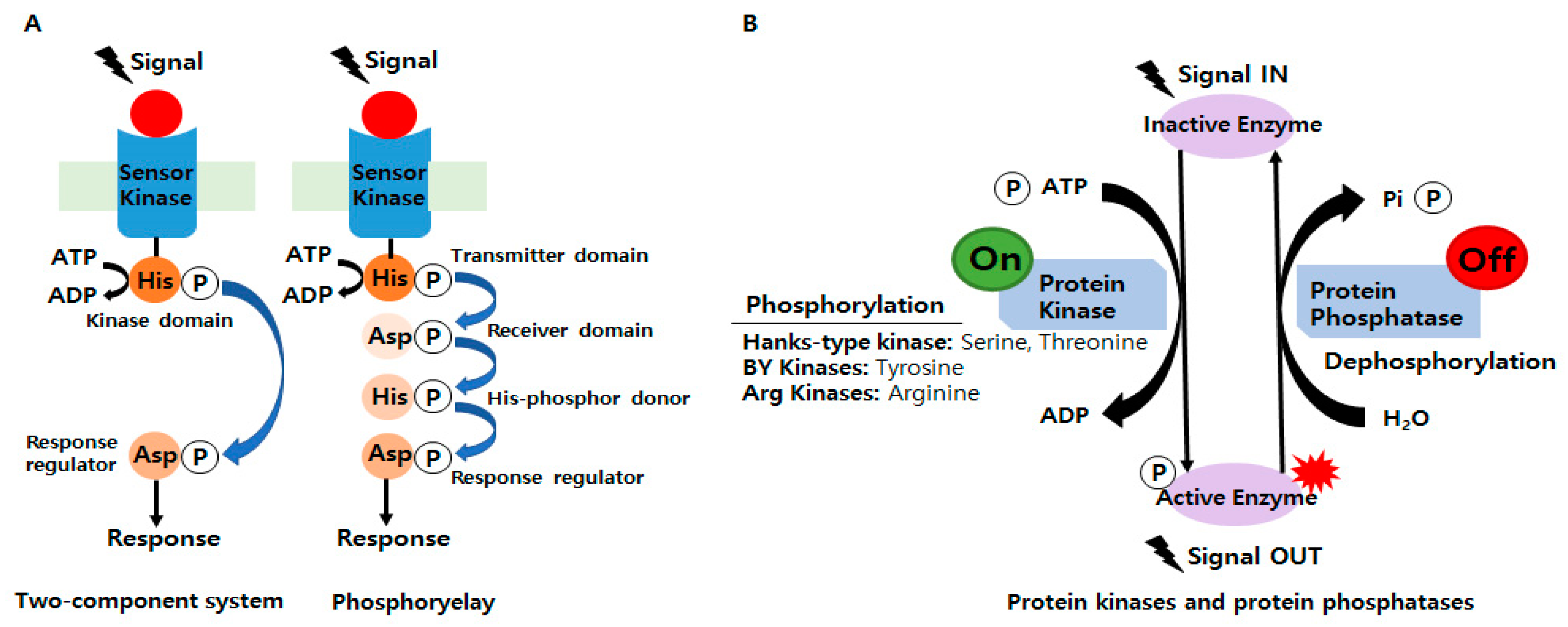
Since the publication of a breakthrough paper on bacterial signaling in 1986, researchers have been able to share their findings on various regulatory systems. In addition, changes in protein phosphorylation and the discovery of amino acid sequence similarities in TCSs have been actively studied [8]. Bacteria sense and respond to numerous external stimuli to survive in various environments [9] and adapt to environmental changes using TCSs and phosphorelays, which are critical mediators of bacterial signal transduction (Figure 1A). In phosphorylases, a sensor kinase first transfers the phosphoryl group to a response regulator with a conserved aspartate domain but no output domain, which is a more complicated type of TCS [10]. TCSs comprise at least two proteins: a sensor kinase and a response regulator. It was predicted that bacteria exhibit signaling phosphorylation mainly at His and Asp residues [11]. The former senses external stimuli, while the latter alters the expression profiles of bacterial genes required for survival and adaptation [12]. In other words, TCSs play a significant role in the general regulatory network by integrating external signals and information from stress pathways, central metabolism, and global regulators [13].
For example, the PhoQ/PhoP TCS detects several host stimuli, such as extracellular magnesium restriction, low pH, cationic antimicrobial peptides, and osmotic stress [5,14]. TCSs are critical for the coordinated expression of virulence factors and, in some situations, for bacterial viability and proliferation. Several studies have shown that TCSs regulate virulence and antibiotic resistance in pathogenic bacteria [9,15,16,17,18,19,20,21,22,23,24,25,26]. Furthermore, the mechanisms of specific TCSs inhibitors differ from those of existing antibiotics and might facilitate the development of effective drugs against drug-resistant bacteria [5,12,15,27,28]. Serine/threonine kinases, which usually have multiple substrates, can also phosphorylate TCS response regulators [5]. The reported regulatory modes of five TCSs in Escherichia coli (E. coli) exhibited novel relationships: MG1655, BaeSR, and CpxAR are stimulated by ethanol stress; KdpDE and PhoRB are induced by low levels of potassium and phosphate, respectively; and ZraSR is stimulated by zinc [29]. Human TCS genes have been compared to TCS genes in Francisella tularensis, a Gram-negative bacterium that causes disease in various hosts [13]. Furthermore, a recent report showed that TCSs utilize multiple mechanisms, such as cross-regulation, to integrate and coordinate input stimuli to control biofilm formation [30,31,32,33,34,35,36].
2.2. Involvements of Ser/Thr/Tyr/Arg Kinases in Bacterial Signaling and Regulation
Unlike TCS histidine kinases, which usually phosphorylate one response regulator, Hanks-type kinases and BY kinases tend to phosphorylate multiple protein substrates (Figure 1B) [5]. In phosphorylases, a sensor kinase first transfers the phosphoryl group to a response regulator with a conserved aspartate domain but no output domain, which is a more complicated type of TCS [10]. Phosphoproteomic surveys over the past decade in phylogenetically diverse bacteria have identified numerous proteins phosphorylated at Ser/Thr (ST) residues [11]. Furthermore, Tyr phosphorylation regulates several cellular processes in bacteria [37,38]. Historically, the phosphorylation of ST residues in bacterial proteins was first identified by pioneering experiments in the 1970s. However, by the early 1980s, most research focused on TCSs [39], HPr kinase/phosphorylases [40,41,42], and the isocitrate dehydrogenase (Icd) kinase/phosphatase system [43,44,45,46]. As a result, researchers arrived at the premature conclusion that eukaryotes possess only Ser/Thr/Tyr (STY) kinases and that bacteria possess mainly His/Asp kinases. On the other hand, with the advent of genomic sequencing in the 1990s, genes encoding ST kinases were widely identified in bacterial genomes [47], and this apparent complexity presented the challenge of identifying the substrates of these bacterial kinases. Comprehensive searches for proteins containing phosphorylated STY residues in E. coli, Bacillus subtilis (B. subtilis), and Lactococcus lactis (L. lactis) in 2007 gave rise to bacterial phosphoproteomics [3,4,47,48]. Since then, hundreds of homologous TCSs have now been identified in eukaryotic organisms. Reversible phosphorylation of STY residues has also been found in many prokaryotes identified as having equal or greater numbers of STYs than eukaryotes [47,49,50,51]. For example, numerous eukaryotic ST kinases that participate in complex signaling pathways help regulate the Myxococcus xanthus (M. xanthus) life cycle [52]. In addition, bacterial kinases with catalytic domains may share structural and functional homology with eukaryotic ST kinases [53,54]. Knowledge of protein kinases/phosphatases has expanded as researchers have further defined bacterial evolutionary conservation. Therefore, the roles of bacterial proteins containing phosphorylated STY residues corresponding to protein kinases/phosphatases during signal transduction need to be fully understood. Bacterial protein phosphorylation, which performs a variety of functions including antibiotic resistance, DNA replication and metabolism, heat shock response, biofilm formation, sporulation, and antitoxicity, is continuously investigated [50,55,56,57,58,59,60]. Several years ago, new evidence suggested that arginine phosphorylation and dephosphorylation are key regulators in bacteria, which implied these modifications might also be important physiologically [61,62,63,64,65,66,67,68,69,70,71].
3. Bacterial Phosphoproteomics
Phosphorylation represents a dynamic change, and phosphoproteins are commonly present at very low levels. As a result, precise and sensitive techniques are needed for phosphoproteome analysis. A large body of phosphoproteomic research has been conducted using mass spectrometry techniques in conjunction with specific phosphor-enrichment techniques [1]. Additionally, specific tools have been developed to study the many substrates of STY kinases. Traditional phosphoproteomics, utilized in bacteriology before 2007, relied on 1D- and 2D-gel 32P-radiolabeling or Western blotting with immunodetection followed by low-resolution mass spectrometry. Although 2D gel electrophoresis enables the simultaneous separation of hundreds of proteins, this tool has poor reproducibility, under-represents low-abundance and hydrophobic proteins, and has a poor dynamic range [72]. Furthermore, the ability of 2D gel electrophoresis to resolve integral membrane proteins is limited because of protein aggregation during the first isoelectric-focusing (IEF) migration, and this technique is particularly ineffective at identifying sites of phosphorylation. However, the efficient enrichment of phosphorylated peptides before mass spectrometry has revolutionized phosphoproteomics, and since 2007, high-resolution mass spectrometry coupled with gel-free analysis has led to the elucidation of site-specific STY bacterial phosphoproteomes in many bacterial species.
3.1. Phosphoproteome Analysis of Beneficial Microorganisms
The first phosphoproteome studies suggested [3,4,48] that phosphorylations are critical regulatory events of bacterial metabolism and showed that bacterial phosphoproteins and phosphorylated residues are associated with evolutionary conservation. Hundreds of biological meaningful phosphorylation sites in bacteria had been found by 2019 [73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88]. The immobilized metal ion affinity chromatography (IMAC) phosphopeptide enrichment technique was used to identify more than 2000 phosphorylated proteins [81,82,89]. In 2021, 19 phosphoproteomic studies on bacteria were reported, and the phosphoproteomes of 14 bacteria were analyzed and biologically interpreted [54,71,90,91,92,93,94,95,96,97,98]. Increasing evidence shows that bacterial phosphorylation sites are as versatile as those of eukaryotes. Furthermore, many studies have emphasized the utilities of protein phosphorylation events and their associated kinases/phosphatases for elucidating the associated physiological processes. Table 1 lists phosphoproteomics studies conducted on 35 bacterial species since the start of phosphoprotein research in 2007. In addition to STY phosphorylation in bacteria, recent research efforts have also studied the phosphorylation of histidine (His, the most abundant bacterial protein) [71,81,82,89,93,99,100] and arginine, which plays a significant role in bacteria [71,82,101]. Prior to 2014, experiments on sub-stoichiometric phosphopeptide enrichment were performed under strong acidic conditions, which explains why phosphorylated histidine residues were difficult to detect. However, phosphorylated His proteins can now be identified using recently developed methods [81,89]. Furthermore, several new methods have been devised to analyze arginine since it was discovered that arginine phosphorylation plays an important role in Gram-positive bacteria [70,71]. The information provided in Table 1 may be expansive, but it provides comprehensive reference information on research techniques and trends for those studying phosphorylated proteins. Furthermore, it provides information for researchers studying specific bacteria regarding the detection of phosphorylated proteins.
3.2. Phylogenetic Diagram of Beneficial Microorganisms
This review also provides an overview of useful microorganisms subjected to phosphoproteomic studies. Figure 2 lists the 35 bacterial species investigated, divides them into 8 phyla, 11 classes, 16 orders, 24 families, and 26 genera, and classifies them as Gram-positive bacteria (P, n = 16) or Gram-negative bacteria (N, n = 19).
Mycobacterium is a genus in the phylum Actinomycetota and is assigned its own family, Mycobacteriaceae. This genus includes pathogens known to cause serious diseases in mammals and tuberculosis in humans. Biochemical and signaling pathways involved in pathogenicity were investigated in virulent H37Rv and non-virulent H37Ra [122] strains to investigate protein phosphorylation networks using clinical isolates of M. tuberculosis [79]. In addition, a phosphoprotein study was undertaken to understand how antibiotic resistance develops [80] and to obtain insights into the regulatory roles of phosphoproteins in Mycobacterium growth and development [78,84].
Antibiotics, such as actinorhodin, methylenomycin, undecylprodigiosin, and perimycin, are produced by different Streptomyces strains [108]. Immobilized zirconium (IV) affinity chromatography and mass spectrometry were used to discover more phosphoproteins [95] and understand the roles of phosphoproteins in Streptomyces coelicolor (S. coelicolor) [106]. Bacterial differentiation and secondary metabolic activation in S. coelicolor were recently investigated using a quantitative mass spectrometry-based/proteomics/ phosphoproteomics approach [85].
Bacilli is a class of Gram-positive aerobic bacteria that includes the orders Bacillaes and Lactobacillales. Bacillales are a representative genus that includes Bacillus, Listeria, and Staphylococcus, and Bacillus subtilis (B. subtilis) is used as a model for research on bacterial cell differentiation and chromosome replication. This bacterium is used commercially to synthesize large amounts of enzymes [125,126,127], and B. subtilis 168 has been reported to contain a number of biologically significant phosphoproteins [3,54,77,93,115]. Listeria monocytogenes (L. monocytogenes) is a pathogenic soil bacterium, and after 143 phosphorylation sites [107] were discovered in this bacterium, an automated STY phosphopeptide enrichment method was devised to investigate the relationship between protein phosphorylation, toxicity mechanisms, and carbon metabolism, and as a result, 420 phosphorylation sites were detected [54]. Phosphorylated proteins in Staphylococcus aureus have been found to be associated with pathogenicity and virulence. An effective phosphopeptide enrichment technique was developed to understand how protein phosphorylation affects complex signaling networks associated with pathogenicity, and eight proteins phosphorylated on arginine residues have been identified [93,101]. Research has shown that arginine phosphorylation plays a significant and relevant role in metabolism [71]. Streptococcus is a genus of Gram-positive coccus or spherical bacteria belonging to the family Streptococcaceae, within the order Lactobacillales in the phylum Bacillota [128]. The pathogenic bacterium Streptococcus pneumoniae, which plays an essential regulatory role in complex protein phosphorylation metabolic pathways and bacterial virulence, has been studied [104]. A systematic study of ST kinases and phosphatases of the pathogen Streptococcus suis (S. suis) was performed using comparative phenotypic, proteomic, and phosphoproteomic assays [91]. In addition, studies were conducted to identify the proteins and pathways tagged by STY phosphorylation in Streptococcus thermophilous (S. thermophilous), a lactic acid bacterium used extensively for dairy fermentation [56]. The class Clostridia includes Clostridium acetobutylicum (C. acetobutylicum), which produces butanol, and Clostridioides difficile (C. difficile), a well-known enteropathogen. The extent and nature of phosphorylation in the Gram-positive enteropathogen C. difficile have not been well characterized. PTMs have been studied [98], and a promising study was conducted to provide detailed mapping of kinase–substrate relationships in C. difficile to identify novel biomarkers and therapeutic targets [96].
Cyanobacteria of the species Microcystis aeruginosa (M. aeruginosa) can play a crucial role in synthesizing cyanotoxins, particularly the potent liver poisons known as microcystins, and thus, the relation between toxin generation and phosphoproteomic profiles was studied in M. aeruginosa [123]. Cyanobacteria, such as Synechocystis sp., play important ecological roles. Ser, Thr, and Tyr phosphorylation contribute to the basic mechanisms that regulate homeostasis in cyanobacteria [113,121].
Thermus is a genus of thermophilic bacteria belonging to the Deinococcota phylum, and the research, biotechnological, and industrial potentials of thermostable enzymes isolated from members of the Thermus genus are of great interest. The phosphoproteins of Thermus thermophilus (T. thermophilus) HB8 identified using phosphoproteome analysis are involved in various cellular processes [112]. In a phosphoproteomic study on T. thermophilus HB27, phosphorylation affected PilF phosphorylation on type IV pilus and biofilm formation [75].
Mycoplasma pneumoniae (M. pneumoniae) belongs to the Mollicutes class and is a diminutive bacterium capable of host-independent life. In humans, M. pneumoniae causes mycoplasma pneumonia, a form of atypical bacterial pneumonia related to cold agglutinin disease. This bacterium exhibits little regulation of gene expression, which is why its phosphorylated proteins are biologically important [74].
Gammaproteobacteria, Alphaproteobacteria, and Betaproteobacteria are classes of bacteria in the phylum Pseudomonadota. Pseudomonas, Moraxella, and Acinetobacter species are pathogens that can cause disease in humans, animals, and plants. Acinetobacter baumannii (A. baumannii) can be pathogenic in individuals with a weakened immune system, and is garnering attention as a cause of nosocomial infections [129]. In one study, the STY phosphoprotein properties of two A. baumannii reference strains (ATCC17978) and a highly invasive, multidrug-resistant clinical isolate (Abh12O-A2) were compared, and the results obtained highlighted the roles of phosphoproteins in pathogenicity and drug resistance [76]. The roles of AmpC β-lactamase phosphorylation were also compared in a mipenem-susceptible Acinetobacter baumannii SK17-S and resistant SK17-R strain [100]. E. coli is a rod-shaped, Gram-negative, facultative anaerobic organism that can be grown and cultured easily and inexpensively in a laboratory environment [130], and studies have confirmed that specific phosphorylated bacterial proteins are involved in translational arrest, growth inhibition, and the induction of physiological dormancy [83]. Phosphoproteomics studies have generated large datasets of bacterial phosphorylated protein with the aim of understanding cellular processes [4,77,83,114,130]. Approximately 30% of Klebsiella pneumoniae (K. pneumoniae) strains naturally present in soil can fix nitrogen in anaerobic environments, and K. pneumoniae has been shown to increase crop yields via nitrogen fixation [131]. Encapsulated K. pneumoniae, an important pathogen in nosocomial infections, contains protein-tyrosine kinases and phosphatases, which are viewed as keys to deciphering its virulence [102]. An enrichment process was developed to identify more phosphopeptides in a single bacterial sample [77]. Rhodopseudomonas palustris (R. palustris) has a variable metabolism and can grow in photoheterotrophic and chemoheterotrophic conditions. This species is used to control carbon metabolism by phosphorylation at the threonine residue and produce hydrogen, lipids, and thus butanol [111]. In addition, the phosphoproteome of Bordetella pertussis, bronchiseptica, and parapertussis were characterized, and their potential roles in Bordetella biology and virulence were examined. Bordetella are pathogens that cause whooping cough or diseases resembling whooping cough. Globally, bordetella infections have increased, necessitating a greater understanding of these diseases and the developments of novel medications and vaccines [92].
4. Conclusions
Bacteria play vital roles in the environment, animals, and humans. Bacterial protein phosphorylation serves diverse functions in bacteria, such as antibiotic resistance, DNA replication and metabolism, heat shock response, biofilm formation, spore formation, anti-virulence, and the production of amino acids and antibiotics. Bacteria contain extremely small amounts of phosphoproteins, but despite this, phosphoproteins influence essential cellular processes. Research on two-component systems (TCSs) and the protein phosphorylated at Ser/Thr/Tyr (STY) residues began in 2008, and hundreds of biologically relevant phosphorylation sites have since been discovered in bacteria. Furthermore, increasing evidence indicates that bacterial phosphorylation sites are as versatile and rich as those in eukaryotes. Advances in proteomic technology have resulted in the discovery of many bacterial phosphoproteins, and advances in LC-MS/MS technology and phosphopeptide enrichment over the last 20 years have enabled the study of large datasets of Ser/Thr/Tyr/Arg phosphopeptides in bacteria. Prior to 2014, experiments on sub-stoichiometric phosphopeptide enrichment were done under strong acidic conditions, which explains why phosphorylated histidine residues were difficult to detect. However, phosphorylated His proteins can now be identified using recently developed methods [81,89]. Furthermore, several new methods have been devised to analyze arginine since it was discovered that arginine phosphorylation plays an important role in Gram-positive bacteria [70,71]. Because technological advances have enabled researchers to determine the biological significances of individual microbes, we undertook this review to summarize studies on the phosphorylation of proteins and the phylogeny of microbes. Table 1 provides a summary of the status of Ser/Thr/Tyr/His/Arg phosphorylated protein analyses conducted on beneficial microorganisms, and Figure 2 summarizes why researchers studied these microorganisms and findings of biological significance. Although this information may be somewhat expansive, it provides comprehensive reference information on research techniques and trends for those studying phosphorylated proteins. Furthermore, it provides information for researchers studying specific bacteria regarding the detection of phosphorylated proteins. This review article was also produced in part to help researchers find information on the biological significance of phosphoproteins and provide information on research ideas and trends.
Funding
This study was supported by a National Research Foundation of Korea (NRF) grant (Grant no. 2021R1G1A1010154) funded by the Korea government (MSIT).
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Thingholm, T.E.; Jensen, O.N.; Larsen, M.R. Analytical strategies for phosphoproteomics. Proteomics 2009, 9, 1451–1468. [Google Scholar] [CrossRef] [PubMed]
- Chubukov, V.; Gerosa, L.; Kochanowski, K.; Sauer, U. Coordination of microbial metabolism. Nat. Rev. Microbiol. 2014, 12, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Macek, B.; Mijakovic, I.; Olsen, J.V.; Gnad, F.; Kumar, C.; Jensen, P.R.; Mann, M. The serine/threonine/tyrosine phosphoproteome of the model bacterium Bacillus subtilis. Mol. Cell. Proteom. 2007, 6, 697–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macek, B.; Gnad, F.; Soufi, B.; Kumar, C.; Olsen, J.V.; Mijakovic, I.; Mann, M. Phosphoproteome Analysis of E. coli Reveals Evolutionary Conservation of Bacterial Ser/Thr/Tyr Phosphorylation. Mol. Cell. Proteom. 2008, 7, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Macek, B.; Forchhammer, K.; Hardouin, J.; Weber-Ban, E.; Grangeasse, C.; Mijakovic, I. Protein post-translational modifications in bacteria. Nat. Rev. Microbiol. 2019, 17, 651–664. [Google Scholar] [CrossRef]
- Ramazi, S.; Zahiri, J. Posttranslational modifications in proteins: Resources, tools and prediction methods. Database J. Biol. Databases Curation 2021, 2021, baab012. [Google Scholar] [CrossRef]
- Mijakovic, I.; Grangeasse, C.; Turgay, K. Exploring the diversity of protein modifications: Special bacterial phosphorylation systems. FEMS Microbiol. Rev. 2016, 40, 398–417. [Google Scholar] [CrossRef] [Green Version]
- Bourret, R.B.; Silversmith, R.E. Two-component signal transduction. Curr. Opin. Microbiol. 2010, 13, 113–115. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.; Jamal, S.B.; Hassan, S.S.; Carvalho, P.; Almeida, S.; Barh, D.; Ghosh, P.; Silva, A.; Castro, T.L.P.; Azevedo, V. Two-Component Signal Transduction Systems of Pathogenic Bacteria As Targets for Antimicrobial Therapy: An Overview. Front. Microbiol. 2017, 8, 1878. [Google Scholar] [CrossRef] [Green Version]
- Mitrophanov, A.Y.; Groisman, E.A. Signal integration in bacterial two-component regulatory systems. Genes Dev. 2008, 22, 2601–2611. [Google Scholar] [CrossRef] [Green Version]
- Rajagopalan, K.; Dworkin, J.; Nagle, E. Identification and Biochemical Characterization of a Novel Protein Phosphatase 2C-Like Ser/Thr Phosphatase in Escherichia coli. J. Bacteriol. 2018, 200, e00225-18, Erratum in J. Bacteriol. 2019, 201, e00648-19. [Google Scholar] [CrossRef] [Green Version]
- Hirakawa, H.; Kurushima, J.; Hashimoto, Y.; Tomita, H. Progress Overview of Bacterial Two-Component Regulatory Systems as Potential Targets for Antimicrobial Chemotherapy. Antibiotics 2020, 9, 635. [Google Scholar] [CrossRef] [PubMed]
- van Hoek, M.L.; Hoang, K.V.; Gunn, J.S. Two-Component Systems in Francisella Species. Front. Cell Infect. Microbiol. 2019, 9, 198. [Google Scholar] [CrossRef]
- Yuan, J.; Jin, F.; Glatter, T.; Sourjik, V. Osmosensing by the bacterial PhoQ/PhoP two-component system. Proc. Natl. Acad. Sci. USA 2017, 114, E10792–E10798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasuhiro Gotoh, Y.E.; Watanabe, T.; Okamoto, S.; Doi, A.; Utsumi, R. Two-component signal transduction as potential drug targets in pathogenic bacteria. Curr. Opin. Microbiol. 2010, 13, 232–239. [Google Scholar] [CrossRef]
- Huang, J.; Li, C.; Song, J.; Velkov, T.; Wang, L.; Zhu, Y.; Li, J. Regulating polymyxin resistance in Gram-negative bacteria: Roles of two-component systems PhoPQ and PmrAB. Future Microbiol. 2020, 15, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Lingzhi, L.; Haojie, G.; Dan, G.; Hongmei, M.; Yang, L.; Mengdie, J.; Chengkun, Z.; Xiaohui, Z. The role of two-component regulatory system in beta-lactam antibiotics resistance. Microbiol. Res. 2018, 215, 126–129. [Google Scholar] [CrossRef]
- Takada, H.; Yoshikawa, H. Essentiality and function of WalK/WalR two-component system: The past, present, and future of research. Biosci. Biotechnol. Biochem. 2018, 82, 741–751. [Google Scholar] [CrossRef]
- Cardona, S.T.; Choy, M.; Hogan, A.M. Essential Two-Component Systems Regulating Cell Envelope Functions: Opportunities for Novel Antibiotic Therapies. J. Membr. Biol. 2018, 251, 75–89. [Google Scholar] [CrossRef]
- Bhagirath, A.Y.; Li, Y.; Patidar, R.; Yerex, K.; Ma, X.; Kumar, A.; Duan, K. Two Component Regulatory Systems and Antibiotic Resistance in Gram-Negative Pathogens. Int. J. Mol. Sci. 2019, 20, 1781. [Google Scholar] [CrossRef] [Green Version]
- Tierney, A.R.; Rather, P.N. Roles of two-component regulatory systems in antibiotic resistance. Future Microbiol. 2019, 14, 533–552. [Google Scholar] [CrossRef]
- Murret-Labarthe, C.; Kerhoas, M.; Dufresne, K.; Daigle, F. New Roles for Two-Component System Response Regulators of Salmonella enterica Serovar Typhi during Host Cell Interactions. Microorganisms 2020, 8, 722. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Du, C.; Huang, H.; Gu, H.; Dong, X.; Hu, Y. TCS response regulator OmpR plays a major role in stress resistance, antibiotic resistance, motility, and virulence in Edwardsiella piscicida. Aquaculture 2022, 559, 738441. [Google Scholar] [CrossRef]
- Shaw, C.; Hess, M.; Weimer, B.C. Two-component systems regulate bacterial virulence in response to the host gastrointestinal environment and metabolic cues. Virulence 2022, 13, 1666–1680. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Dhankhar, P.; Kumar, P. Bacterial histidine kinases as potential antibacterial drug targets. In Protein Kinase Inhibitors; Elsevier: Amsterdam, The Netherlands, 2022; pp. 711–734. [Google Scholar]
- Kwiecinski, J.M.; Jelani, D.A.; Fuentes, E.J.; Horswill, A.R. Therapeutic Inhibition of Staphylococcus aureus ArlRS Two-Component Regulatory System Blocks Virulence. Antimicrob. Agents Chemother. 2022, 66, e00187-22. [Google Scholar] [CrossRef]
- Kundu, M. The role of two-component systems in the physiology of Mycobacterium tuberculosis. IUBMB Life 2018, 70, 710–717. [Google Scholar] [CrossRef] [Green Version]
- Schaefers, M.M. Regulation of Virulence by Two-Component Systems in Pathogenic Burkholderia. Infect. Immun. 2020, 88, e00927-19. [Google Scholar] [CrossRef]
- Choudhary, K.S.; Kleinmanns, J.A.; Decker, K.; Sastry, A.V.; Gao, Y.; Szubin, R.; Seif, Y.; Palsson, B.O. Elucidation of Regulatory Modes for Five Two-Component Systems in Escherichia coli Reveals Novel Relationships. mSystems 2020, 5, e00980-20. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Sun, D.; Zhu, J.; Liu, W. Two-Component Signal Transduction Systems: A Major Strategy for Connecting Input Stimuli to Biofilm Formation. Front. Microbiol. 2018, 9, 3279. [Google Scholar] [CrossRef]
- Badal, D.; Jayarani, A.V.; Kollaran, M.A.; Kumar, A.; Singh, V. Pseudomonas aeruginosa biofilm formation on endotracheal tubes requires multiple two-component systems. J. Med. Microbiol. 2020, 69, 906–919. [Google Scholar] [CrossRef]
- Kera, K.; Yoshizawa, Y.; Shigehara, T.; Nagayama, T.; Tsujii, M.; Tochigi, S.; Uozumi, N. Hik36-Hik43 and Rre6 act as a two-component regulatory system to control cell aggregation in Synechocystis sp. PCC6803. Sci. Rep. 2020, 10, 19405. [Google Scholar] [CrossRef]
- Sionov, R.V.; Steinberg, D. Targeting the Holy Triangle of Quorum Sensing, Biofilm Formation, and Antibiotic Resistance in Pathogenic Bacteria. Microorganisms 2022, 10, 1239. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, V.; Tiwari, M.; Joshi, R.; Tiwari, V. Therapeutic strategies against potential antibiofilm targets of multidrug-resistant Acinetobacter baumannii. J. Cell. Physiol. 2022, 237, 2045–2063. [Google Scholar] [CrossRef]
- Jabbour, N.; Morello, E.; Camiade, E.; Lartigue, M.-F. Biofilm Formation in Streptococcus agalactiae Is Inhibited by a Small Regulatory RNA Regulated by the Two-Component System CiaRH. Microbiol. Spectr. 2022, 10, e0063522. [Google Scholar] [CrossRef]
- Ballén, V.; Cepas, V.; Ratia, C.; Gabasa, Y.; Soto, S.M. Clinical Escherichia coli: From Biofilm Formation to New Antibiofilm Strategies. Microorganisms 2022, 10, 1103. [Google Scholar] [CrossRef] [PubMed]
- Whitmore, S.E.; Lamont, R.J. Tyrosine phosphorylation and bacterial virulence. Int. J. Oral Sci. 2012, 4, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cozzone, A.J. Role of protein phosphorylation on serine/threonine and tyrosine in the virulence of bacterial pathogens. J. Mol. Microbiol. Biotechnol. 2005, 9, 198–213. [Google Scholar] [CrossRef] [PubMed]
- Stock, A.M.; Robinson, V.L.; Goudreau, P.N. Two-component signal transduction. Annu. Rev. Biochem. 2000, 69, 183–215. [Google Scholar] [CrossRef] [Green Version]
- Van Dijk, A.A.; De Lange, L.C.M.; Bachovchin, W.W.; Robillard, G.T. Effect of phosphorylation on hydrogen-bonding interactions of the active site histidine of the phosphocarrier protein HPr of the phosphoenolpyruvate-dependent phosphotransferase system determined by nitrogen-15 NMR spectroscopy. Biochemistry 1990, 29, 8164–8171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reizer, J.; Romano, A.H.; Deutscher, J. The role of phosphorylation of HPr, a phosphocarrier protein of the phosphotransferase system, in the regulation of carbon metabolism in gram-positive bacteria. J. Cell Biochem. 1993, 51, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Mijakovic, I.; Poncet, S.; Galinier, A.; Monedero, V.; Fieulaine, S.; Janin, J.; Nessler, S.; Marquez, J.A.; Scheffzek, K.; Hasenbein, S.; et al. Pyrophosphate-producing protein dephosphorylation by HPr kinase/phosphorylase: A relic of early life? Proc. Natl. Acad. Sci. USA 2002, 99, 13442–13447. [Google Scholar] [CrossRef] [Green Version]
- LaPorte, D.C. The isocitrate dehydrogenase phosphorylation cycle: Regulation and enzymology. J. Cell. Biochem. 1993, 51, 14–18. [Google Scholar] [CrossRef]
- Laporte, D.C.; Stueland, C.S.; Ikeda, T.P. Isocitrate dehydrogenase kinase/phosphatase. Biochimie 1989, 71, 1051–1057. [Google Scholar] [CrossRef]
- Garnak, M.; Reeves, H.C. Phosphorylation of Isocitrate dehydrogenase of Escherichia coli. Science 1979, 203, 1111–1112. [Google Scholar] [CrossRef] [PubMed]
- Stueland, C.S.; Gorden, K.; LaPorte, D.C. The isocitrate dehydrogenase phosphorylation cycle. Identification of the primary rate-limiting step. J. Biol. Chem. 1988, 263, 19475–19479. [Google Scholar] [CrossRef] [PubMed]
- Mijakovic, I. Protein phosphorylation in bacteria. Microbe 2010, 5, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Soufi, B.; Gnad, F.; Jensen, P.R.; Petranovic, D.; Mann, M.; Mijakovic, I.; Macek, B. The Ser/Thr/Tyr phosphoproteome of Lactococcus lactis IL1403 reveals multiply phosphorylated proteins. Proteomics 2008, 8, 3486–3493. [Google Scholar] [CrossRef]
- Chao, J.D.; Wong, D.; Av-Gay, Y. Microbial protein-tyrosine kinases. J. Biol. Chem. 2014, 289, 9463–9472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellinzoni, M.; Wehenkel, A.M.; Duran, R.; Alzari, P.M. Novel mechanistic insights into physiological signaling pathways mediated by mycobacterial Ser/Thr protein kinases. Microbes Infect. 2019, 21, 222–229. [Google Scholar] [CrossRef]
- Getz, L.J.; Runte, C.S.; Rainey, J.K.; Thomas, N.A. Tyrosine Phosphorylation as a Widespread Regulatory Mechanism in Prokaryotes. J. Bacteriol. 2019, 201, e00205-19. [Google Scholar] [CrossRef] [Green Version]
- Nariya, H.; Inouye, S. Identification of a protein Ser/Thr kinase cascade that regulates essential transcriptional activators in Myxococcus xanthus development. Mol. Microbiol. 2005, 58, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.F.; Goss, L.; Dworkin, J. Eukaryote-like serine/threonine kinases and phosphatases in bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 192–212. [Google Scholar] [CrossRef] [Green Version]
- Birk, M.S.; Charpentier, E.; Frese, C.K. Automated Phosphopeptide Enrichment for Gram-Positive Bacteria. J. Proteome Res. 2021, 20, 4886–4892. [Google Scholar] [CrossRef]
- Bonne Kohler, J.; Jers, C.; Senissar, M.; Shi, L.; Derouiche, A.; Mijakovic, I. Importance of protein Ser/Thr/Tyr phosphorylation for bacterial pathogenesis. FEBS Lett. 2020, 594, 2339–2369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, C.; Haller, L.; Blein-Nicolas, M.; Zivy, M.; Canette, A.; Verbrugghe, M.; Mezange, C.; Boulay, M.; Gardan, R.; Samson, S.; et al. Identification of Hanks-Type Kinase PknB-Specific Targets in the Streptococcus thermophilus Phosphoproteome. Front. Microbiol. 2019, 10, 1329. [Google Scholar] [CrossRef] [PubMed]
- Szoke, T.; Albocher, N.; Govindarajan, S.; Nussbaum-Shochat, A.; Amster-Choder, O. Tyrosine phosphorylation-dependent localization of TmaR that controls activity of a major bacterial sugar regulator by polar sequestration. Proc. Natl. Acad. Sci. USA 2021, 118, e2016017118. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Sammartino, J.C.; Costantino, L.; Gelain, A.; Meneghetti, F.; Villa, S.; Chiarelli, L.R. An Overview on the Potential Antimycobacterial Agents Targeting Serine/Threonine Protein Kinases from Mycobacterium tuberculosis. Curr. Top. Med. Chem. 2019, 19, 646–661. [Google Scholar] [CrossRef]
- Hirschfeld, C.; Gomez-Mejia, A.; Bartel, J.; Hentschker, C.; Rohde, M.; Maass, S.; Hammerschmidt, S.; Becher, D. Proteomic Investigation Uncovers Potential Targets and Target Sites of Pneumococcal Serine-Threonine Kinase StkP and Phosphatase PhpP. Front. Microbiol. 2019, 10, 3101. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Garcia, T.; Poncet, S.; Cuenot, E.; Douché, T.; Gianetto, Q.G.; Peltier, J.; Courtin, P.; Chapot-Chartier, M.-P.; Matondo, M.; Dupuy, B.; et al. Ser/Thr kinase-dependent phosphorylation of the peptidoglycan hydrolase CwlA controls its export and modulates cell division in Clostridioides difficile. bioRxiv 2020, 12, e00519-21. [Google Scholar] [CrossRef]
- Andrews, L.D.; Graham, J.; Snider, M.J.; Fraga, D. Characterization of a novel bacterial arginine kinase from Desulfotalea psychrophila. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 150, 312–319. [Google Scholar] [CrossRef]
- Elsholz, A.K.; Turgay, K.; Michalik, S.; Hessling, B.; Gronau, K.; Oertel, D.; Mäder, U.; Bernhardt, J.; Becher, D.; Hecker, M. Global impact of protein arginine phosphorylation on the physiology of Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2012, 109, 7451–7456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.; Trentini, D.B.; Spiess, S.; Fuhrmann, J.; Ammerer, G.; Mechtler, K.; Clausen, T. Quantitative phosphoproteomics reveals the role of protein arginine phosphorylation in the bacterial stress response. Mol. Cell. Proteom. 2014, 13, 537–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trentini, D.B.; Suskiewicz, M.J.; Heuck, A.; Kurzbauer, R.; Deszcz, L.; Mechtler, K.; Clausen, T. Arginine phosphorylation marks proteins for degradation by a Clp protease. Nature 2016, 539, 48–53. [Google Scholar] [CrossRef]
- Junker, S.; Maaß, S.; Otto, A.; Hecker, M.; Becher, D.R. Toward the quantitative characterization of arginine phosphorylations in Staphylococcus aureus. J. Proteome Res. 2018, 18, 265–279. [Google Scholar] [PubMed]
- Junker, S.; Maaβ, S.; Otto, A.; Michalik, S.; Morgenroth, F.; Gerth, U.; Hecker, M.; Becher, D. Spectral library based analysis of arginine phosphorylations in Staphylococcus aureus. Mol. Cell. Proteom. 2018, 17, 335–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.; Choi, Y.; Lee, D.; Seo, J.K.; Kee, J.M. Distinct phosphorylation and dephosphorylation dynamics of protein arginine kinases revealed by fluorescent activity probes. Chem. Commun. 2019, 55, 7482–7485. [Google Scholar] [CrossRef]
- Suskiewicz, M.J.; Hajdusits, B.; Beveridge, R.; Heuck, A.; Vu, L.D.; Kurzbauer, R.; Hauer, K.; Thoeny, V.; Rumpel, K.; Mechtler, K. Structure of McsB, a protein kinase for regulated arginine phosphorylation. Nat. Chem. Biol. 2019, 15, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Semanjski, M.; Orlovetskie, N.; Bhattacharya, S.; Alon, S.; Argaman, L.; Jarrous, N.; Zhang, Y.; Macek, B.; Sinai, L. Arginine dephosphorylation propels spore germination in bacteria. Proc. Natl. Acad. Sci. USA 2019, 116, 14228–14237. [Google Scholar] [CrossRef] [Green Version]
- Ogbonna, E.C.; Anderson, H.R.; Schmitz, K.R. Identification of Arginine Phosphorylation in Mycolicibacterium smegmatis. Microbiol. Spectr. 2022, 10, e0204222. [Google Scholar] [CrossRef] [PubMed]
- Prust, N.; van Breugel, P.C.; Lemeer, S. Widespread Arginine Phosphorylation in Staphylococcus aureus. Mol. Cell. Proteom. 2022, 21, 100232. [Google Scholar] [CrossRef]
- Kolkman, A.; Daran-Lapujade, P.; Fullaondo, A.; Olsthoorn, M.M.A.; Pronk, J.T.; Slijper, M.; Heck, A.J.R. Proteome analysis of yeast response to various nutrient limitations. Mol. Syst. Biol. 2006, 2, 2006.0026. [Google Scholar] [CrossRef]
- Voisin, S.; Watson, D.C.; Tessier, L.; Ding, W.; Foote, S.; Bhatia, S.; Kelly, J.F.; Young, N.M. The cytoplasmic phosphoproteome of the Gram-negative bacterium Campylobacter jejuni: Evidence for modification by unidentified protein kinases. Proteomics 2007, 7, 4338–4348. [Google Scholar] [CrossRef]
- Schmidl, S.R.; Gronau, K.; Pietack, N.; Hecker, M.; Becher, D.; Stulke, J. The phosphoproteome of the minimal bacterium Mycoplasma pneumoniae: Analysis of the complete known Ser/Thr kinome suggests the existence of novel kinases. Mol. Cell Proteom. 2010, 9, 1228–1242. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.L.; Liao, J.H.; Lin, G.H.; Lin, M.H.; Chang, Y.C.; Liang, S.Y.; Yang, F.L.; Khoo, K.H.; Wu, S.H. Phosphoproteomic analysis reveals the effects of PilF phosphorylation on type IV pilus and biofilm formation in Thermus thermophilus HB27. Mol. Cell Proteom. 2013, 12, 2701–2713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, N.C.; Spat, P.; Mendez, J.A.; Nakedi, K.; Aranda, J.; Bou, G. Ser/Thr/Tyr phosphoproteome characterization of Acinetobacter baumannii: Comparison between a reference strain and a highly invasive multidrug-resistant clinical isolate. J. Proteom. 2014, 102, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-H.; Sugiyama, N.; Ishihama, Y. Systematic profiling of the bacterial phosphoproteome reveals bacterium-specific features of phosphorylation. Sci. Signal. 2015, 8, rs10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakedi, K.C.; Nel, A.J.; Garnett, S.; Blackburn, J.M.; Soares, N.C. Comparative Ser/Thr/Tyr phosphoproteomics between two mycobacterial species: The fast growing Mycobacterium smegmatis and the slow growing Mycobacterium bovis BCG. Front. Microbiol. 2015, 6, 237. [Google Scholar] [CrossRef] [Green Version]
- Fortuin, S.; Tomazella, G.G.; Nagaraj, N.; Sampson, S.L.; Gey van Pittius, N.C.; Soares, N.C.; Wiker, H.G.; de Souza, G.A.; Warren, R.M. Phosphoproteomics analysis of a clinical Mycobacterium tuberculosis Beijing isolate: Expanding the mycobacterial phosphoproteome catalog. Front. Microbiol. 2015, 6, 6. [Google Scholar] [CrossRef]
- de Keijzer, J.; Mulder, A.; de Beer, J.; de Ru, A.H.; van Veelen, P.A.; van Soolingen, D. Mechanisms of Phenotypic Rifampicin Tolerance in Mycobacterium tuberculosis Beijing Genotype Strain B0/W148 Revealed by Proteomics. J. Proteome Res. 2016, 15, 1194–1204. [Google Scholar] [CrossRef] [PubMed]
- Potel, C.M.; Lin, M.H.; Heck, A.J.R.; Lemeer, S. Defeating Major Contaminants in Fe3+-Immobilized Metal Ion Affinity Chromatography (IMAC) Phosphopeptide Enrichment. Mol. Cell Proteom. 2018, 17, 1028–1034. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.H.; Potel, C.M.; Tehrani, K.; Heck, A.J.R.; Martin, N.I.; Lemeer, S. A New Tool to Reveal Bacterial Signaling Mechanisms in Antibiotic Treatment and Resistance. Mol. Cell Proteom. 2018, 17, 2496–2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semanjski, M.; Germain, E.; Bratl, K.; Kiessling, A.; Gerdes, K.; Macek, B. The kinases HipA and HipA7 phosphorylate different substrate pools in Escherichia coli to promote multidrug tolerance. Sci. Signal. 2018, 11, eaat5750. [Google Scholar] [CrossRef] [PubMed]
- Albeldas, C.; Ganief, N.; Calder, B.; Nakedi, K.C.; Garnett, S.; Nel, A.J.M.; Blackburn, J.M.; Soares, N.C. Global proteome and phosphoproteome dynamics indicate novel mechanisms of vitamin C induced dormancy in Mycobacterium smegmatis. J. Proteom. 2018, 180, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rioseras, B.; Shliaha, P.V.; Gorshkov, V.; Yagüe, P.; López-García, M.T.; Gonzalez-Quiñonez, N.; Kovalchuk, S.; Rogowska-Wrzesinska, A.; Jensen, O.N.; Manteca, A. Quantitative Proteome and Phosphoproteome Analyses of Streptomyces coelicolor Reveal Proteins and Phosphoproteins Modulating Differentiation and Secondary Metabolism. Mol. Cell Proteom. 2018, 17, 1591–1611. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Hou, L.; Liu, M.; Xu, X.; Gao, Q.; Deng, J.; Xiang, S.; Cao, Q.; Zhou, M.; Yang, Q.; et al. Phosphoproteomic Analysis of Spiroplasma eriocheiris and Crosstalk with Acetylome Reveals the Role of Post-Translational Modifications in Metabolism. Curr. Proteom. 2019, 17, 392–403. [Google Scholar] [CrossRef]
- Lim, S. Quantitative Bacterial Phosphoproteomics, Electronic, Scholarly Journal. Ph.D. Thesis, The University of Queensland, Brisbane, Australia, 2015. [Google Scholar] [CrossRef]
- Yague, P.; Gonzalez-Quinonez, N.; Fernanez-Garcia, G.; Alonso-Fernandez, S.; Manteca, A. Goals and Challenges in Bacterial Phosphoproteomics. Int. J. Mol. Sci. 2019, 20, 5678. [Google Scholar] [CrossRef] [Green Version]
- Potel, C.M.; Lin, M.-H.; Heck, A.J.R.; Lemeer, S. Widespread bacterial protein histidine phosphorylation revealed by mass spectrometry-based proteomics. Nat. Methods 2018, 15, 187–190. [Google Scholar] [CrossRef]
- Baros-Steyl, S.S.; Nakedi, K.C.; Ganief, T.A.; Okendo, J.O.; Tabb, D.L.; Soares, N.C.; Blackburn, J.M. Phosphoproteomics reveals new insights into the role of PknG during the persistence of pathogenic mycobacteria in host macrophages. bioRxiv 2021. [Google Scholar] [CrossRef]
- Hu, Q.; Yao, L.; Liao, X.; Zhang, L.-S.; Li, H.-T.; Li, T.-T.; Jiang, Q.-G.; Tan, M.-F.; Li, L.; Draheim, R.R.; et al. Comparative Phenotypic, Proteomic, and Phosphoproteomic Analysis Reveals Different Roles of Serine/Threonine Phosphatase and Kinase in the Growth, Cell Division, and Pathogenicity of Streptococcus suis. Microorganisms 2021, 9, 2442. [Google Scholar] [CrossRef]
- Luu, L.D.W.; Zhong, L.; Kaur, S.; Raftery, M.J.; Lan, R. Comparative Phosphoproteomics of Classical Bordetellae Elucidates the Potential Role of Serine, Threonine and Tyrosine Phosphorylation in Bordetella Biology and Virulence. Front. Cell. Infect. Microbiol. 2021, 11, 660280. [Google Scholar] [CrossRef]
- Prust, N.; van der Laarse, S.; van den Toorn, H.W.P.; van Sorge, N.M.; Lemeer, S. In-Depth Characterization of the Staphylococcus aureus Phosphoproteome Reveals New Targets of Stk1. Mol. Cell Proteom. 2021, 20, 100034. [Google Scholar] [CrossRef] [PubMed]
- Sultan, A.; Jers, C.; Ganief, T.A.; Shi, L.; Senissar, M.; Kohler, J.B.; Macek, B.; Mijakovic, I. Phosphoproteome Study of Escherichia coli Devoid of Ser/Thr Kinase YeaG during the Metabolic Shift from Glucose to Malate. Front. Microbiol. 2021, 12, 657562. [Google Scholar] [CrossRef]
- Alonso-Fernández, S.; Arribas-Díez, I.; Fernández-García, G.; González-Quiñónez, N.; Jensen, O.N.; Manteca, A. Quantitative phosphoproteome analysis of Streptomyces coelicolor by immobilized zirconium (IV) affinity chromatography and mass spectrometry reveals novel regulated protein phosphorylation sites and sequence motifs. J. Proteom. 2022, 269, 104719. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Garcia, T.; Douché, T.; Giai Gianetto, Q.; Poncet, S.; El Omrani, N.; Smits, W.K.; Cuenot, E.; Matondo, M.; Martin-Verstraete, I. In-Depth Characterization of the Clostridioides difficile Phosphoproteome to Identify Ser/Thr Kinase Substrates. Mol. Cell. Proteom. 2022, 21, 100428. [Google Scholar] [CrossRef] [PubMed]
- Šarić, E.; Quinn, G.A.; Nalpas, N.; Paradžik, T.; Kazazić, S.; Filić, Ž.; Šemanjski, M.; Herron, P.; Hunter, I.; Maček, B.; et al. Phosphoproteome Dynamics of Streptomyces rimosus during Submerged Growth and Antibiotic Production. mSystems 2022, 7, e0019922. [Google Scholar] [CrossRef]
- Smits Wiep, K.; Mohammed, Y.; de Ru Arnoud, H.; Cordo, V.; Friggen Annemieke, H.; van Veelen Peter, A.; Hensbergen Paul, J. Clostridioides difficile Phosphoproteomics Shows an Expansion of Phosphorylated Proteins in Stationary Growth Phase. mSphere 2022, 7, e0091121. [Google Scholar] [CrossRef]
- Licona-Cassani, C.; Lim, S.; Marcellin, E.; Nielsen, L.K. Temporal dynamics of the Saccharopolyspora erythraea phosphoproteome. Mol. Cell Proteom. 2014, 13, 1219–1230. [Google Scholar] [CrossRef] [Green Version]
- Lai, J.H.; Yang, J.T.; Chern, J.; Chen, T.L.; Wu, W.L.; Liao, J.H.; Tsai, S.F.; Liang, S.Y.; Chou, C.C.; Wu, S.H. Comparative Phosphoproteomics Reveals the Role of AmpC β-lactamase Phosphorylation in the Clinical Imipenem-resistant Strain Acinetobacter baumannii SK17. Mol. Cell Proteom. 2016, 15, 12–25. [Google Scholar] [CrossRef] [Green Version]
- Basell, K.; Otto, A.; Junker, S.; Zuhlke, D.; Rappen, G.M.; Schmidt, S.; Hentschker, C.; Macek, B.; Ohlsen, K.; Hecker, M.; et al. The phosphoproteome and its physiological dynamics in Staphylococcus aureus. Int. J. Med. Microbiol. 2014, 304, 121–132. [Google Scholar] [CrossRef]
- Lin, M.H.; Hsu, T.L.; Lin, S.Y.; Pan, Y.J.; Jan, J.T.; Wang, J.T.; Khoo, K.H.; Wu, S.H. Phosphoproteomics of Klebsiella pneumoniae NTUH-K2044 reveals a tight link between tyrosine phosphorylation and virulence. Mol. Cell Proteom. 2009, 8, 2613–2623. [Google Scholar] [CrossRef] [Green Version]
- Ravichandran, A.; Sugiyama, N.; Tomita, M.; Swarup, S.; Ishihama, Y. Ser/Thr/Tyr phosphoproteome analysis of pathogenic and non-pathogenic Pseudomonas species. Proteomics 2009, 9, 2764–2775. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Ge, F.; Xiao, C.-L.; Yin, X.-F.; Ge, R.; Zhang, L.-H.; He, Q.-Y. Phosphoproteomic Analysis Reveals the Multiple Roles of Phosphorylation in Pathogenic Bacterium Streptococcus pneumoniae. J. Proteome Res. 2010, 9, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Prisic, S.; Dankwa, S.; Schwartz, D.; Chou, M.F.; Locasale, J.W.; Kang, C.-M.; Bemis, G.; Church, G.M.; Steen, H.; Husson, R.N. Extensive phosphorylation with overlapping specificity by Mycobacterium tuberculosis serine/threonine protein kinases. Proc. Natl. Acad. Sci. USA 2010, 107, 7521–7526. [Google Scholar] [CrossRef] [Green Version]
- Parker, J.L.; Jones, A.M.; Serazetdinova, L.; Saalbach, G.; Bibb, M.J.; Naldrett, M.J. Analysis of the phosphoproteome of the multicellular bacterium Streptomyces coelicolor A3(2) by protein/peptide fractionation, phosphopeptide enrichment and high-accuracy mass spectrometry. Proteomics 2010, 10, 2486–2497. [Google Scholar] [CrossRef]
- Misra, S.K.; Milohanic, E.; Ake, F.; Mijakovic, I.; Deutscher, J.; Monnet, V.; Henry, C. Analysis of the serine/threonine/tyrosine phosphoproteome of the pathogenic bacterium Listeria monocytogenes reveals phosphorylated proteins related to virulence. Proteomics 2011, 11, 4155–4165. [Google Scholar] [CrossRef] [PubMed]
- Manteca, A.; Ye, J.; Sanchez, J.; Jensen, O.N. Phosphoproteome analysis of Streptomyces development reveals extensive protein phosphorylation accompanying bacterial differentiation. J. Proteome Res. 2011, 10, 5481–5492. [Google Scholar] [CrossRef]
- Ge, R.; Sun, X.; Xiao, C.; Yin, X.; Shan, W.; Chen, Z.; He, Q.Y. Phosphoproteome analysis of the pathogenic bacterium Helicobacter pylori reveals over-representation of tyrosine phosphorylation and multiply phosphorylated proteins. Proteomics 2011, 11, 1449–1461. [Google Scholar] [CrossRef]
- Bai, X.; Ji, Z. Phosphoproteomic investigation of a solvent producing bacterium Clostridium acetobutylicum. Appl. Microbiol. Biotechnol. 2012, 95, 201–211. [Google Scholar] [CrossRef]
- Hu, C.-W.; Lin, M.-H.; Huang, H.-C.; Ku, W.-C.; Yi, T.-H.; Tsai, C.-F.; Chen, Y.-J.; Sugiyama, N.; Ishihama, Y.; Juan, H.-F.; et al. Phosphoproteomic Analysis of Rhodopseudomonas palustris Reveals the Role of Pyruvate Phosphate Dikinase Phosphorylation in Lipid Production. J. Proteome Res. 2012, 11, 5362–5375. [Google Scholar] [CrossRef]
- Takahata, Y.; Inoue, M.; Kim, K.; Iio, Y.; Miyamoto, M.; Masui, R.; Ishihama, Y.; Kuramitsu, S. Close proximity of phosphorylation sites to ligand in the phosphoproteome of the extreme thermophile Thermus thermophilus HB8. Proteomics 2012, 12, 1414–1430. [Google Scholar] [CrossRef]
- Yang, M.K.; Qiao, Z.X.; Zhang, W.Y.; Xiong, Q.; Zhang, J.; Li, T.; Ge, F.; Zhao, J.D. Global phosphoproteomic analysis reveals diverse functions of serine/threonine/tyrosine phosphorylation in the model cyanobacterium Synechococcus sp. strain PCC 7002. J. Proteome Res. 2013, 12, 1909–1923. [Google Scholar] [CrossRef]
- Soares, N.C.; Spat, P.; Krug, K.; Macek, B. Global dynamics of the Escherichia coli proteome and phosphoproteome during growth in minimal medium. J. Proteome Res. 2013, 12, 2611–2621. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, V.; Shi, L.; Krug, K.; Derouiche, A.; Jers, C.; Cousin, C.; Kobir, A.; Mijakovic, I.; Macek, B. Quantitative phosphoproteome analysis of Bacillus subtilis reveals novel substrates of the kinase PrkC and phosphatase PrpC. Mol. Cell Proteom. 2014, 13, 1965–1978. [Google Scholar] [CrossRef] [Green Version]
- Ouidir, T.; Jarnier, F.; Cosette, P.; Jouenne, T.; Hardouin, J. Extracellular Ser/Thr/Tyr phosphorylated proteins of Pseudomonas aeruginosa PA14 strain. Proteomics 2014, 14, 2017–2030. [Google Scholar] [CrossRef] [PubMed]
- Misra, S.K.; Moussan Désirée Aké, F.; Wu, Z.; Milohanic, E.; Cao, T.N.; Cossart, P.; Deutscher, J.; Monnet, V.; Archambaud, C.; Henry, C. Quantitative proteome analyses identify PrfA-responsive proteins and phosphoproteins in Listeria monocytogenes. J. Proteome Res. 2014, 13, 6046–6057. [Google Scholar] [CrossRef]
- Liu, T.; Tian, C.F.; Chen, W.X. Site-Specific Ser/Thr/Tyr Phosphoproteome of Sinorhizobium meliloti at Stationary Phase. PLoS ONE 2015, 10, e0139143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, A.; Soufi, B.; Ravikumar, V.; Soares, N.C.; Krug, K.; Smith, Y.; Macek, B.; Ben-Yehuda, S. Phosphoproteome dynamics mediate revival of bacterial spores. BMC Biol. 2015, 13, 76. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.; Marcellin, E.; Jacob, S.; Nielsen, L.K. Global dynamics of Escherichia coli phosphoproteome in central carbon metabolism under changing culture conditions. J. Proteom. 2015, 126, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Spät, P.; Maček, B.; Forchhammer, K. Phosphoproteome of the cyanobacterium Synechocystis sp. PCC 6803 and its dynamics during nitrogen starvation. Front. Microbiol. 2015, 6, 248. [Google Scholar] [CrossRef] [Green Version]
- Verma, R.; Pinto, S.M.; Patil, A.H.; Advani, J.; Subba, P.; Kumar, M.; Sharma, J.; Dey, G.; Ravikumar, R.; Buggi, S.; et al. Quantitative Proteomic and Phosphoproteomic Analysis of H37Ra and H37Rv Strains of Mycobacterium tuberculosis. J. Proteome Res. 2017, 16, 1632–1645. [Google Scholar] [CrossRef]
- Qu, J.; Shen, L.; Zhao, M.; Li, W.; Jia, C.; Zhu, H.; Zhang, Q. Determination of the Role of Microcystis aeruginosa in Toxin Generation Based on Phosphoproteomic Profiles. Toxins 2018, 10, 304. [Google Scholar] [CrossRef] [Green Version]
- Tatli, M.; Hebert, A.S.; Coon, J.J.; Amador-Noguez, D. Genome Wide Phosphoproteome Analysis of Zymomonas mobilis under Anaerobic, Aerobic, and N(2)-Fixing Conditions. Front. Microbiol. 2019, 10, 1986. [Google Scholar] [CrossRef] [PubMed]
- Errington, J.; Aart, L.T.V. Microbe Profile: Bacillus subtilis: Model organism for cellular development, and industrial workhorse. Microbiology 2020, 166, 425–427. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.I.; Rahman, M.M.; Salam, M.A.; Khan, M.A.R.; Islam, M.T. Identification of marine sponge-associated bacteria of the Saint Martin’s island of the Bay of Bengal emphasizing on the prevention of motile Aeromonas septicemia in Labeo rohita. Aquaculture 2021, 545, 737156. [Google Scholar] [CrossRef]
- Rahman, M.M.; Paul, S.I.; Akter, T.; Tay, A.C.Y.; Foysal, M.J.; Islam, M.T. Whole-Genome Sequence of Bacillus subtilis WS1A, a Promising Fish Probiotic Strain Isolated from Marine Sponge of the Bay of Bengal. Microbiol. Resour. Announc. 2020, 9, e00641-20. [Google Scholar] [CrossRef]
- Sriskandan, S.; Slater, J.D. Invasive disease and toxic shock due to zoonotic Streptococcus suis: An emerging infection in the East? PLoS Med. 2006, 3, e187. [Google Scholar] [CrossRef] [Green Version]
- Antunes, L.C.; Visca, P.; Towner, K.J. Acinetobacter baumannii: Evolution of a global pathogen. Pathog. Dis. 2014, 71, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Mateus, A.; Hevler, J.; Bobonis, J.; Kurzawa, N.; Shah, M.; Mitosch, K.; Goemans, C.V.; Helm, D.; Stein, F.; Typas, A.; et al. The functional proteome landscape of Escherichia coli. Nature 2020, 588, 473–478. [Google Scholar] [CrossRef]
- Riggs, P.J.; Chelius, M.; Iniguez, A.L.; Kaeppler, S.; Triplett, E. Enhanced maize productivity by inoculation with diazotrophic bacteria. Aust. J. Plant Physiol. 2001, 28, 829–836. [Google Scholar] [CrossRef]
Figure 1.
(A) Schematics of the prototypical two-component systems (TCSs) and phosphorelay systems in bacteria. (B) The overall mechanism of protein phosphorylation regulated by protein kinases and protein phosphatase.
Figure 1.
(A) Schematics of the prototypical two-component systems (TCSs) and phosphorelay systems in bacteria. (B) The overall mechanism of protein phosphorylation regulated by protein kinases and protein phosphatase.

Figure 2.
Taxonomic classifications of beneficial microorganisms subjected to phosphorylation studies. (P): Gram-positive bacteria and (N): Gram-negative bacteria.
Figure 2.
Taxonomic classifications of beneficial microorganisms subjected to phosphorylation studies. (P): Gram-positive bacteria and (N): Gram-negative bacteria.


{kind=link}
{kind=link}
Table 1.
Bacterial Ser/Thr/Tyr/His/Arg phosphoprotemics studies.
| Organism | Strain | Year | P-pro. (ea) | P-pep. (ea) | P-site (ea) | Ser (%) | Thr (%) | Tyr (%) | Arg (%) | His (%) | Refs. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| C. jejuni | 11168 | 2007 | 36 | 58 | 35 | 30.3 | 72.7 | 9.1 | [73] | ||
| B. subtilis | 168 | 2007 | 78 | 103 | 78 | 69.2 | 20.5 | 10.3 | [3] | ||
| L. lactis | Il1403 | 2008 | 63 | 102 | 79 | 46.5 | 50.6 | 2.7 | [48] | ||
| E. coli K12 | MG1665 | 2008 | 79 | 105 | 81 | 67.9 | 23.5 | 8.6 | [4] | ||
| K. pneumoniae | K2044 | 2009 | 81 | 117 | 93 | 31.2 | 15.4 | 25.8 | [102] | ||
| P. putida | MK25 | 2009 | 40 | 56 | 53 | 52.8 | 39.6 | 7.5 | [103] | ||
| P. aeruginosa | PAO1 | 2009 | 23 | 57 | 55 | 52.7 | 32.7 | 14.5 | [103] | ||
| M. pneumoniae | M129 | 2010 | 63 | 16 | 16 | 53.3 | 46.7 | 0 | [74] | ||
| S. pneumoniae | D39 | 2010 | 84 | 102 | 163 | 47.2 | 43.8 | 9 | [104] | ||
| M. tuberculosis | H37Rv | 2010 | 301 | 381 | 506 | 40 | 60 | 0 | [105] | ||
| S. coelicolor | A3(2) | 2010 | 40 | 44 | 46 | 34.1 | 52.3 | 13.6 | [106] | ||
| L. monocytogenes | EGDe | 2011 | 112 | 155 | 143 | 93 | 43 | 7 | [107] | ||
| S. coelicolor | M145 | 2011 | 127 | 260 | 289 | 46.8 | 48 | 5.2 | [108] | ||
| H. pylori | 26695 | 2011 | 67 | 80 | 124 | 42.8 | 38.7 | 18.5 | [109] | ||
| C. acetobutylicum | ATCC824 | 2012 | 61 | 82 | 107 | 42 | 47.6 | 10.6 | [110] | ||
| R. palustris(Ch) | CGA010 | 2012 | 54 | 100 | 63 | 63.3 | 16.1 | 19.4 | [111] | ||
| R. palustris(Ph) | CGA010 | 2012 | 42 | 74 | 59 | 58.9 | 23.2 | 17.9 | [111] | ||
| T. thermophilus | HB8 | 2012 | 48 | 52 | 46 | 30 | 12 | 4 | [112] | ||
| T. thermophilus | HB27 | 2013 | 53 | 93 | 67 | 57 | 36 | 7 | [75] | ||
| Synechococcus sp. | PCC7002 | 2013 | 245 | 280 | 410 | 43.9 | 42.4 | 13.6 | [113] | ||
| E. coli K12 | BW25113 | 2013 | 133 | 150 | 108 | 75.9 | 16.7 | 7.4 | [114] | ||
| S. aureus | COL | 2014 | 108 | 68 | 50 | 25 | 15 | 10 | [101] | ||
| A. baumannii | AbH120A2 | 2014 | 70 | 80 | 70.8 | 25.2 | 3.8 | [76] | |||
| A. baumannii | 17978 | 2014 | 41 | 48 | 68.9 | 24.1 | 5.2 | [76] | |||
| B. subtilis | 168 | 2014 | 177 | 155 | 74.6 | 18.6 | 7.3 | [115] | |||
| S. erythraea | NRRL2338 | 2014 | 88 | 109 | 47 | 45 | 8 | 5.3 | [99] | ||
| P. aeruginosa | PA14 | 2014 | 28 | 43 | 59 | 49 | 24 | 27 | [116] | ||
| L. monocytogenes | ∆PrfA | 2014 | 191 | 256 | 242 | 155 | 75 | 12 | [117] | ||
| S. meliloti | CCBAU | 2015 | 77 | 88 | 96 | 63 | 28 | 5 | [118] | ||
| B. subtilis | Spore | 2015 | 124 | 155 | 77.41 | 22.6 | [119] | ||||
| B. subtilis | 168 | 2015 | 175 | 441 | 339 | 74.8 | 17.7 | 7.1 | [77] | ||
| E. coli K12 | BW25113 | 2015 | 392 | 1212 | 1088 | 69.5 | 21.8 | 7.7 | [77] | ||
| E. coli K12 | MG1655 | 2015 | 71 | 82 | [120] | ||||||
| K. pneumoniae | K2044 | 2015 | 286 | 663 | 559 | 72.9 | 13.7 | 12.9 | [77] | ||
| Synechocystis sp. | PCC 6803 | 2015 | 188 | 242 | 262 | [121] | |||||
| M. tuberculosis | SAW5527 | 2015 | 214 | 303 | 414 | 38 | 59 | 3 | [79] | ||
| M. smegmatis | mc2155 | 2015 | 2462 | 464 | 185 | 39.5 | 57.1 | 3.5 | [78] | ||
| M. bovis BCG | 1173P2 | 2015 | 1765 | 402 | 442 | 35 | 61.6 | 3.1 | [78] | ||
| M. tuberculosis | B0/W148 | 2016 | 132 | 180 | 191 | 22 | 76 | 2 | [80] | ||
| A. baumannii | SK17-S | 2016 | 248 | 351 | 410 | 47 | 27.6 | 12.4 | 4.9 | [100] | |
| A. baumannii | SK17-R | 2016 | 211 | 240 | 285 | 41.4 | 29.5 | 17.5 | 4.9 | [100] | |
| M. tuberculosis | H37Ra | 2017 | 257 | 512 | 29 | 68 | 3 | [122] | |||
| M. smegmatis | mc2155 | 2018 | 154 | 222 | 242 | 24.8 | 74.0 | 1.2 | [84] | ||
| M. aeruginosa | FACHB-469 | 2018 | 37 | 59 | [123] | ||||||
| M. aeruginosa | FACHB-905 | 2018 | 18 | 26 | [123] | ||||||
| S. coelicolor | M145 | 2018 | 48 | 92 | 85 | 50.6 | 47.4 | 2 | [85] | ||
| E. coli K12 | MG1665 | 2018 | 632 | 1178 | 1183 | [83] | |||||
| E. coli K12 | W3110 | 2018 | 861 | 2446 | 57.2 | 25.3 | 8.5 | 9 | [81] | ||
| E. coli K12 | W3110 | 2018 | 781 | 2057 | 2129 | 1220 | 501 | 162 | 246 | [89] | |
| E. coli K12 | W3110 | 2018 | 2248 | 56 | 20 | 13 | 5 | 5 | [82] | ||
| Z. mobilis | ZM4,31821 | 2019 | 125 | 177 | 73 | 21 | 6 | [124] | |||
| S. thermophilus | LMD9 | 2019 | 106 | 410 | 161 | 43 | 33 | 23 | [56] | ||
| S. eriocheiris | M207170 | 2019 | 245 | 465 | [86] | ||||||
| E. coli K12 | 1655, ∆yea | 2021 | 83 | 127 | 67.7 | 28.3 | 3.9 | [94] | |||
| B. subtilis | 168 | 2021 | 146 | 283 | 267 | 73 | 12.7 | 7.5 | 6.7 | [93] | |
| S. aureus | USA300 | 2021 | 859 | 3800 | 3771 | 55.2 | 29.6 | 7.3 | 7.8 | [93] | |
| B. subtilis | 168 | 2021 | 153 | 214 | 67 | 28 | 5 | [54] | |||
| S. pyogenes | M1 | 2021 | 205 | 449 | 41 | 55 | 4 | [54] | |||
| L. monocytogenes | EGDe | 2021 | 241 | 420 | 56 | 35 | 9 | [54] | |||
| B. pertussis | L1423 | 2021 | 45 | 53 | 54 | 72 | 17 | 11 | [92] | ||
| B. bronchiseptica | RB50 | 2021 | 23 | 28 | 29 | 69 | 21 | 10 | [92] | ||
| B. parapertussis | 12822 | 2021 | 42 | 50 | 50 | 80 | 12 | 8 | [92] | ||
| M. bovis | BCG, ΔPknG | 2021 | 914 | 1371 | 1401 | 85.3 | 13.4 | 1.3 | [90] | ||
| S. suis | WT, Δstp | 2021 | 50 | 73 | [91] | ||||||
| S. suis | WT, Δstk | 2021 | 67 | 87 | [91] | ||||||
| S. aureus | NE98, ΔSdrE | 2022 | 953 | 4407 | 45.5 | 24 | 5 | 20.2 | 5.4 | [71] | |
| S. aureus | NE217, ΔStk1 | 2022 | 903 | 3779 | 48.1 | 22 | 6.7 | 18 | 5.2 | [71] | |
| S. aureus | NE1919, ΔStp1 | 2022 | 951 | 4085 | 40.2 | 21.2 | 6.1 | 26 | 6.5 | [71] | |
| C. difficile | 630WT | 2022 | 700 | 2994 | 1759 | 75 | 20 | 5 | [98] | ||
| C. difficile | 630WT, Δ erm | 2022 | 504 | 1061 | 117 | 76.6 | 17.8 | 5.6 | [96] | ||
| S. rimosus | G7, 10970 | 2022 | 230 | 273 | 417 | 41.3 | 53.5 | 5.3 | [97] | ||
| S. coelicolor | A3(2) | 2022 | 187 | 351 | 361 | 41 | 56.2 | 2.8 | [95] | ||
Experimental phosphoproteome coverage is shown in terms of identified phosphorylated proteins (P-pro.), phosphopeptides (P-pep.), and phosphorylated sites (P-site). Data were extracted from research publications or databases. Blank areas: not reported; (Ch) chemoheterotrophic growth; (Ph) photoheterotrophic growth.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lim, S. A Review of the Bacterial Phosphoproteomes of Beneficial Microbes. Microorganisms 2023, 11, 931. https://doi.org/10.3390/microorganisms11040931
AMA Style
Lim S. A Review of the Bacterial Phosphoproteomes of Beneficial Microbes. Microorganisms. 2023; 11(4):931. https://doi.org/10.3390/microorganisms11040931
Chicago/Turabian StyleLim, Sooa. 2023. "A Review of the Bacterial Phosphoproteomes of Beneficial Microbes" Microorganisms 11, no. 4: 931. https://doi.org/10.3390/microorganisms11040931
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.