Figure 1.
Log counts of Streptococcus thermophilus showing its acid tolerance as influenced by different ingredients over an incubation period of 120 min. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
Figure 1.
Log counts of Streptococcus thermophilus showing its acid tolerance as influenced by different ingredients over an incubation period of 120 min. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
Figure 2.
Log counts of Lactobacillus bulgaricus showing its acid tolerance as influenced by different ingredients over an incubation period of 120 min. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
Figure 2.
Log counts of Lactobacillus bulgaricus showing its acid tolerance as influenced by different ingredients over an incubation period of 120 min. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
Figure 3.
Log counts of Streptococcus thermophilus showing its bile tolerance as influenced by treatments over 8 h. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Average of three replicates. A,B Values with different letters at 0 h are significantly different (p < 0.05). a,b,c Values with different letters at 4 h are significantly different (p < 0.05). A,B,C Values with different letters at 8 h are significantly different (p < 0.05). Error bars represent SE.
Figure 3.
Log counts of Streptococcus thermophilus showing its bile tolerance as influenced by treatments over 8 h. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Average of three replicates. A,B Values with different letters at 0 h are significantly different (p < 0.05). a,b,c Values with different letters at 4 h are significantly different (p < 0.05). A,B,C Values with different letters at 8 h are significantly different (p < 0.05). Error bars represent SE.
Figure 4.
Log counts of Lactobacillus bulgaricus showing its bile tolerance as influenced by ingredients over 8 h. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Average of three replicates. a: Values with different letters at 4 h are significantly different (p < 0.05). A There were no significant (p > 0.05) differences at 0 h. A,B, Values with different letters at 4 h are significantly different (p < 0.05). A,B Values with different letters at 8 h are significantly different (p < 0.05). Error bars represent SE.
Figure 4.
Log counts of Lactobacillus bulgaricus showing its bile tolerance as influenced by ingredients over 8 h. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Average of three replicates. a: Values with different letters at 4 h are significantly different (p < 0.05). A There were no significant (p > 0.05) differences at 0 h. A,B, Values with different letters at 4 h are significantly different (p < 0.05). A,B Values with different letters at 8 h are significantly different (p < 0.05). Error bars represent SE.
Figure 5.
Log counts of Streptococcus thermophilus showing its growth as influenced by ingredients over the incubation period of 16 h. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
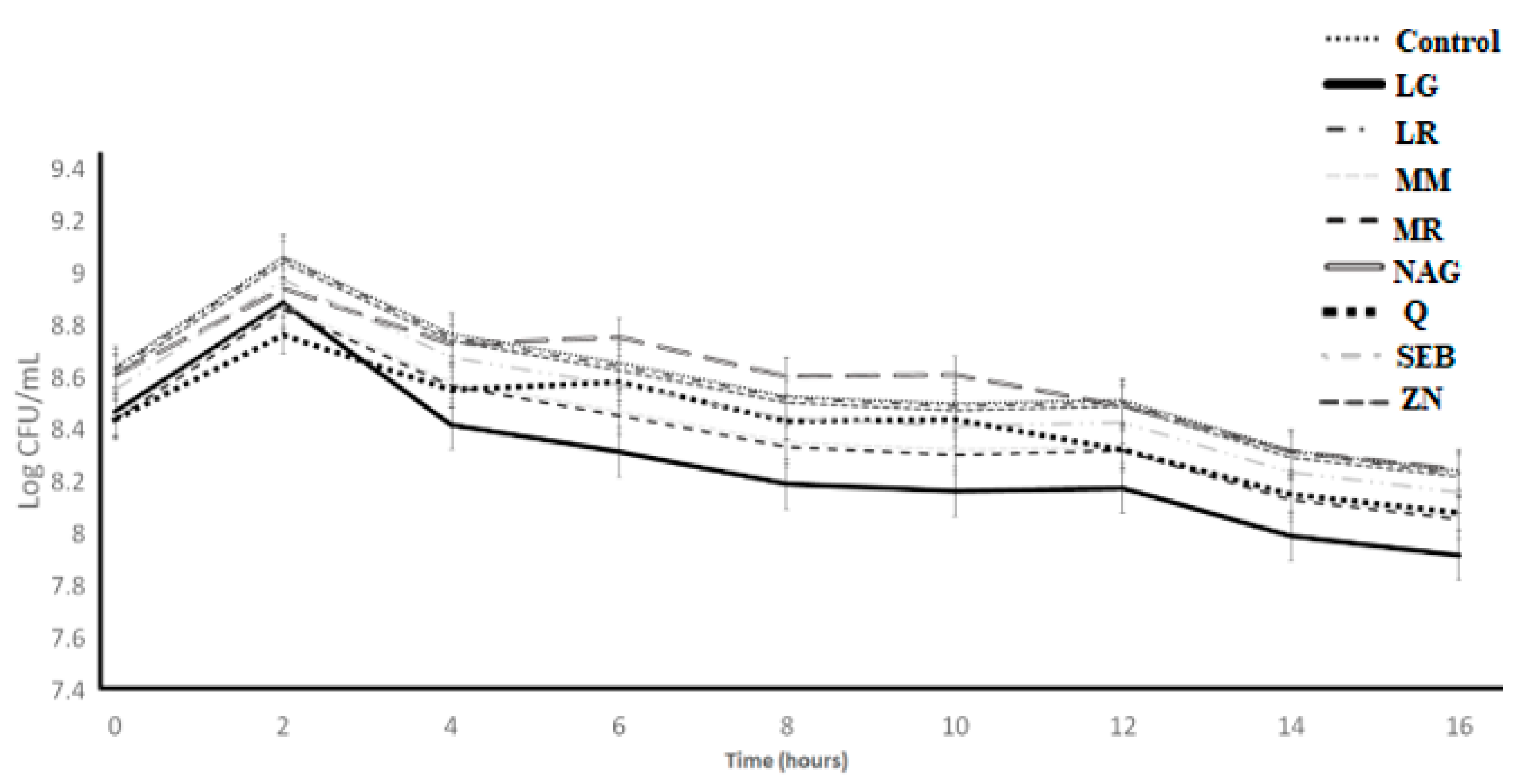
Figure 5.
Log counts of Streptococcus thermophilus showing its growth as influenced by ingredients over the incubation period of 16 h. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
Figure 6.
Log counts of Lactobacillus bulgaricus showing its growth as influenced by ingredients over the incubation period of 16 h. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
Figure 6.
Log counts of Lactobacillus bulgaricus showing its growth as influenced by ingredients over the incubation period of 16 h. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
Figure 7.
Log counts of Streptococcus thermophilus showing its resistance to simulated gastric juice as influenced by ingredients over different pH conditions. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
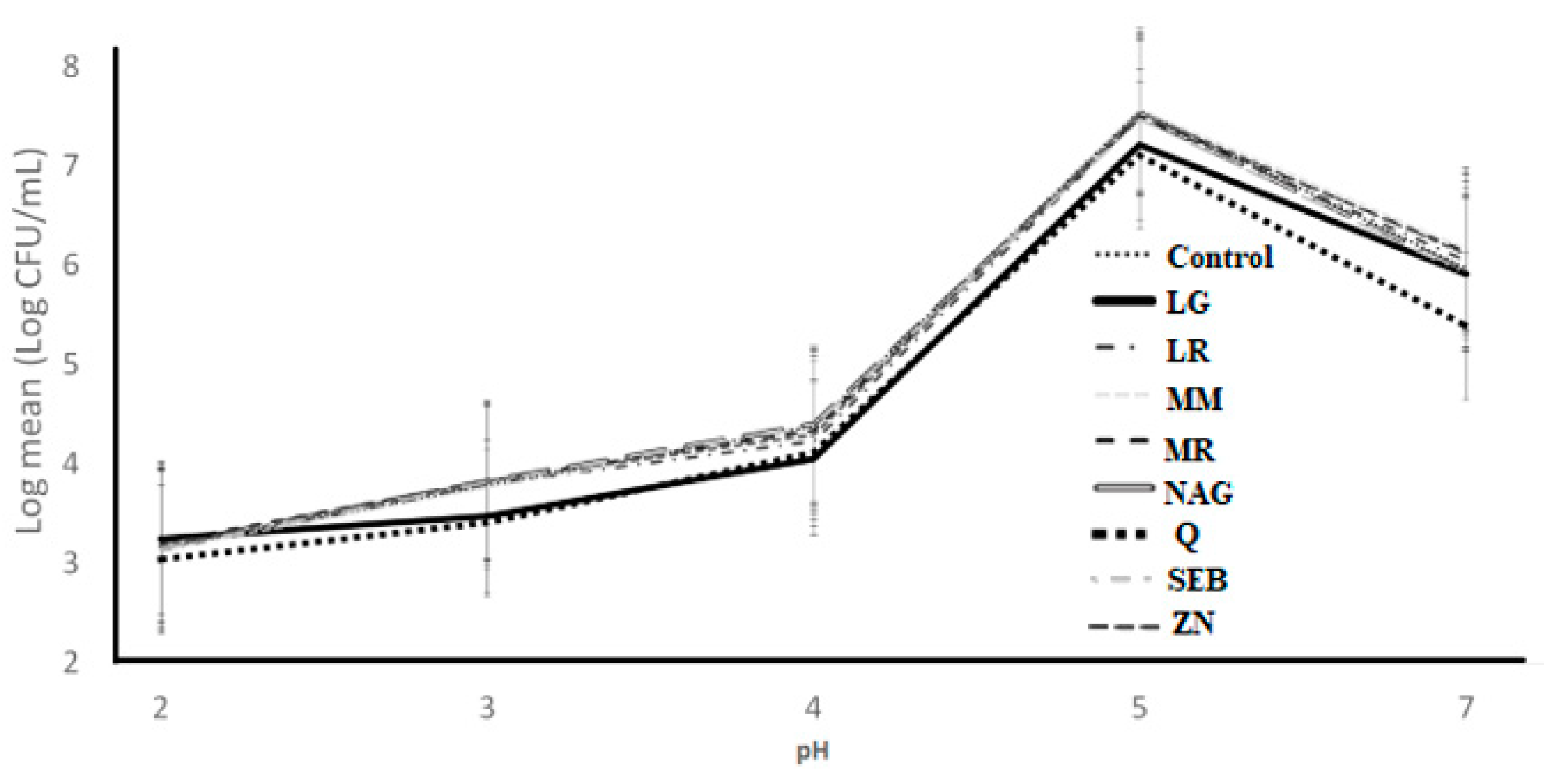
Figure 7.
Log counts of Streptococcus thermophilus showing its resistance to simulated gastric juice as influenced by ingredients over different pH conditions. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
Figure 8.
Log counts of Lactobacillus bulgaricus showing its resistance to simulated gastric juice as influenced by ingredients over different pH conditions. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
Figure 8.
Log counts of Lactobacillus bulgaricus showing its resistance to simulated gastric juice as influenced by ingredients over different pH conditions. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
Figure 9.
Log counts of Streptococcus thermophilus showing its resistance to lysozyme as influenced by ingredients during an incubation time of 120 min. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
Figure 9.
Log counts of Streptococcus thermophilus showing its resistance to lysozyme as influenced by ingredients during an incubation time of 120 min. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
Figure 10.
Log counts of Lactobacillus bulgaricus showing its resistance to lysozyme as influenced by ingredients during an incubation time of 120 min. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
Figure 10.
Log counts of Lactobacillus bulgaricus showing its resistance to lysozyme as influenced by ingredients during an incubation time of 120 min. C = control, LG = L-glutamine (7 g/L), Q = quercetin (700 mg/L), SEB = slippery elm bark (210 mg/L), MR = marshmallow root (1340 mg/L), NAG = N-acetyl-D-glucosamine (210 mg/L), LR = licorice root (210 mg/L), MM = maitake mushrooms (42 mg/L), and ZN = zinc orotate (70 mg/L). Error bars represent SE.
Table 1.
The p-value > F-value of ingredient, time or pH, and their interaction for bacterial viability, bile tolerance, acid tolerance, resistance to gastric juices, protease activity, and lysozyme resistance S. thermophilus and L. bulgaricus.
Table 1.
The p-value > F-value of ingredient, time or pH, and their interaction for bacterial viability, bile tolerance, acid tolerance, resistance to gastric juices, protease activity, and lysozyme resistance S. thermophilus and L. bulgaricus.
| Effect | S. thermophilus STI-06 | L. bulgaricus LB-12 |
|---|
| Viability |
| Ingredient | <0.0001 | <0.0001 |
| Time (Hours) | <0.0001 | <0.0001 |
| Ingredient × time | <0.0001 | <0.0001 |
| Bile tolerance |
| Ingredient | <0.0001 | <0.0001 |
| Time (Hours) | <0.0001 | <0.0001 |
| Ingredient × time | <0.0001 | <0.0001 |
| Acid tolerance |
| Ingredient | <0.0001 | 0.0576 |
| Time (Minutes) | <0.0001 | <0.0001 |
| Ingredient × time | <0.0001 | 0.0775 |
| Resistance to gastric juices |
| Ingredient | <0.0001 | 0.0765 |
| pH | <0.0001 | <0.0001 |
| Ingredient × pH | <0.0001 | 0.1450 |
| Protease activity |
| Ingredient | <0.0001 | 0.0579 |
| Time (Hours) | <0.0001 | <0.0001 |
| Ingredient × time | <0.0001 | 0.4460 |
| Lysozyme resistance |
| Ingredient | <0.0001 | <0.0001 |
| Time (Minutes) | <0.0001 | <0.0001 |
| Ingredient × time | <0.0001 | <0.0001 |
Table 2.
Least squares means for acid and bile tolerance of L. bulgaricus and S. thermophilus as influenced by ingredients.
Table 2.
Least squares means for acid and bile tolerance of L. bulgaricus and S. thermophilus as influenced by ingredients.
| Ingredient | S. thermophilus | L. bulgaricus |
|---|
| Acid tolerance |
| C | 5.848 c | NS |
| LG | 4.460 d | NS |
| Q | 5.790 c | NS |
| SEB | 5.054 c | NS |
| MR | 5.908 bc | NS |
| NAG | 5.944 bc | NS |
| LR | 5.977 ab | NS |
| MM | 6.003 a | NS |
| ZN | 5.866 c | NS |
| Bile tolerance |
| C | 7.656 a | 5.543 a |
| LG | 5.132 b | 4.960 b |
| Q | 7.087 a | 5.124 a |
| SEB | 7.654 a | 5.254 a |
| MR | 7.754 a | 5.548 a |
| NAG | 7.287 a | 5.439 a |
| LR | 7.940 a | 5.487 a |
| MM | 7.476 a | 5.487 a |
| ZN | 7.245 a | 5.557 a |
Table 3.
Mean log difference (log cfu/mL) in the acid tolerance and resistance to lysozyme counts of Streptococcus salivarius ssp. thermophilus ST-M5.
Table 3.
Mean log difference (log cfu/mL) in the acid tolerance and resistance to lysozyme counts of Streptococcus salivarius ssp. thermophilus ST-M5.
| Ingredient | S. thermophilus |
|---|
| Acid Tolerance |
| C | 3.72 |
| LG | 2.93 |
| Q | 3.53 |
| SEB | 3.77 |
| MR | 2.99 |
| NAG | 2.99 |
| LR | 3.02 |
| MM | 3.70 |
| ZN | 3.07 |
| Resistance to Lysozyme |
| C | 4.01 |
| LG | 4.18 |
| Q | 4.77 |
| SEB | 4.23 |
| MR | 4.49 |
| NAG | 3.82 |
| LR | 3.87 |
| MM | 3.92 |
| ZN | 3.89 |
Table 4.
Least squares means for viability and gastric juices tolerance of L. bulgaricus and S. thermophilus as influenced by ingredients.
Table 4.
Least squares means for viability and gastric juices tolerance of L. bulgaricus and S. thermophilus as influenced by ingredients.
| Ingredient | S. thermophilus | L. bulgaricus |
|---|
| Viability |
| C | 8.772 a | 7.965 a |
| LG | 8.578 b | 7.645 b |
| Q | 8.832 a | 7.95 a |
| SEB | 8.876 a | 7.834 a |
| MR | 8.765 a | 8.023 a |
| NAG | 8.786 a | 7.934 a |
| LR | 8.785 a | 7.911 a |
| MM | 8.865 a | 7.845 a |
| ZN | 8.874 a | 7.928 a |
| Resistance to gastric juices |
| C | 5.176 a | NS |
| LG | 4.654 b | NS |
| Q | 5.434 a | NS |
| SEB | 5.156 a | NS |
| MR | 5.467 a | NS |
| NAG | 5.215 a | NS |
| LR | 5.237 a | NS |
| MM | 5.125 a | NS |
| ZN | 5.190 a | NS |
| Resistance to lysozyme |
| C | 5.006 b | 4.861 b |
| LG | 4.967 b | 4.811 b |
| Q | 5.562 a | 5.785 a |
| SEB | 5.102 b | 5.766 a |
| MR | 5.498 a | 5.531 a |
| NAG | 5.054 b | 4.954 b |
| LR | 5.106 b | 5.544 a |
| MM | 5.142 b | 5.046 b |
| ZN | 5.121 b | 4.883 b |
Table 5.
Least square means for protease activity (absorbance) of Streptococcus thermophilus STI-06 over an incubation period of 24 h as influenced by added ingredients.
Table 5.
Least square means for protease activity (absorbance) of Streptococcus thermophilus STI-06 over an incubation period of 24 h as influenced by added ingredients.
| Sample | 0 h | 12 h | 24 h |
|---|
| C | 0.148 ± 0.005 a | 0.155 ± 0.005 d | 0.202 ± 0.011 c |
| LG | 0.151 ± 0.013 a | 0.170 ± 0.013 c | 0.221 ± 0.017 c |
| Q | 0.155 ± 0.019 a | 0.205 ± 0.019 b | 0.217 ± 0.024 c |
| SEB | 0.154 ± 0.021 a | 0.186 ± 0.021 c | 0.232 ± 0.028 bc |
| MR | 0.150 ± 0.023 a | 0.247 ± 0.023 a | 0.320 ± 0.027 a |
| NAG | 0.157 ± 0.012 a | 0.207 ± 0.012 b | 0.271 ± 0.015 b |
| LR | 0.152 ± 0.010 a | 0.175 ± 0.010 c | 0.227 ± 0.022 c |
| MM | 0.155 ± 0.017 a | 0.192 ± 0.017 bc | 0.249 ± 0.013 b |
| ZN | 0.150 ± 0.007 a | 0.173 ± 0.007 c | 0.225 ± 0.022 c |
Table 6.
Least square means for protease activity (absorbance) of Lactobacillus bulgaricus LB-12 over an incubation period of 24 h as influenced by added ingredients.
Table 6.
Least square means for protease activity (absorbance) of Lactobacillus bulgaricus LB-12 over an incubation period of 24 h as influenced by added ingredients.
| Sample | 0 h | 12 h | 24 h |
|---|
| C | 0.160 ± 0.009 | 0.313 ± 0.006 | 0.414 ± 0.017 |
| LG | 0.163 ± 0.007 | 0.320 ± 0.013 | 0.433 ± 0.021 |
| Q | 0.162 ± 0.005 | 0.335 ± 0.020 | 0.447 ± 0.016 |
| SEB | 0.159 ± 0.005 | 0.327 ± 0.015 | 0.441 ± 0.012 |
| MR | 0.160 ± 0.006 | 0.321 ± 0.017 | 0.427 ± 0.023 |
| NAG | 0.164 ± 0.007 | 0.333 ± 0.014 | 0.439 ± 0.020 |
| LR | 0.166 ± 0.004 | 0.343 ± 0.015 | 0.428 ± 0.018 |
| MM | 0.161 ± 0.007 | 0.330 ± 0.019 | 0.427 ± 0.020 |
| ZN | 0.159 ± 0.011 | 0.327 ± 0.022 | 0.432 ± 0.015 |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}