
Detection of Pathogens and Ticks on Sedentary and Migratory Birds in Two Corsican Wetlands (France, Mediterranean Area)
 , , and
, , and
Abstract
:1. Introduction
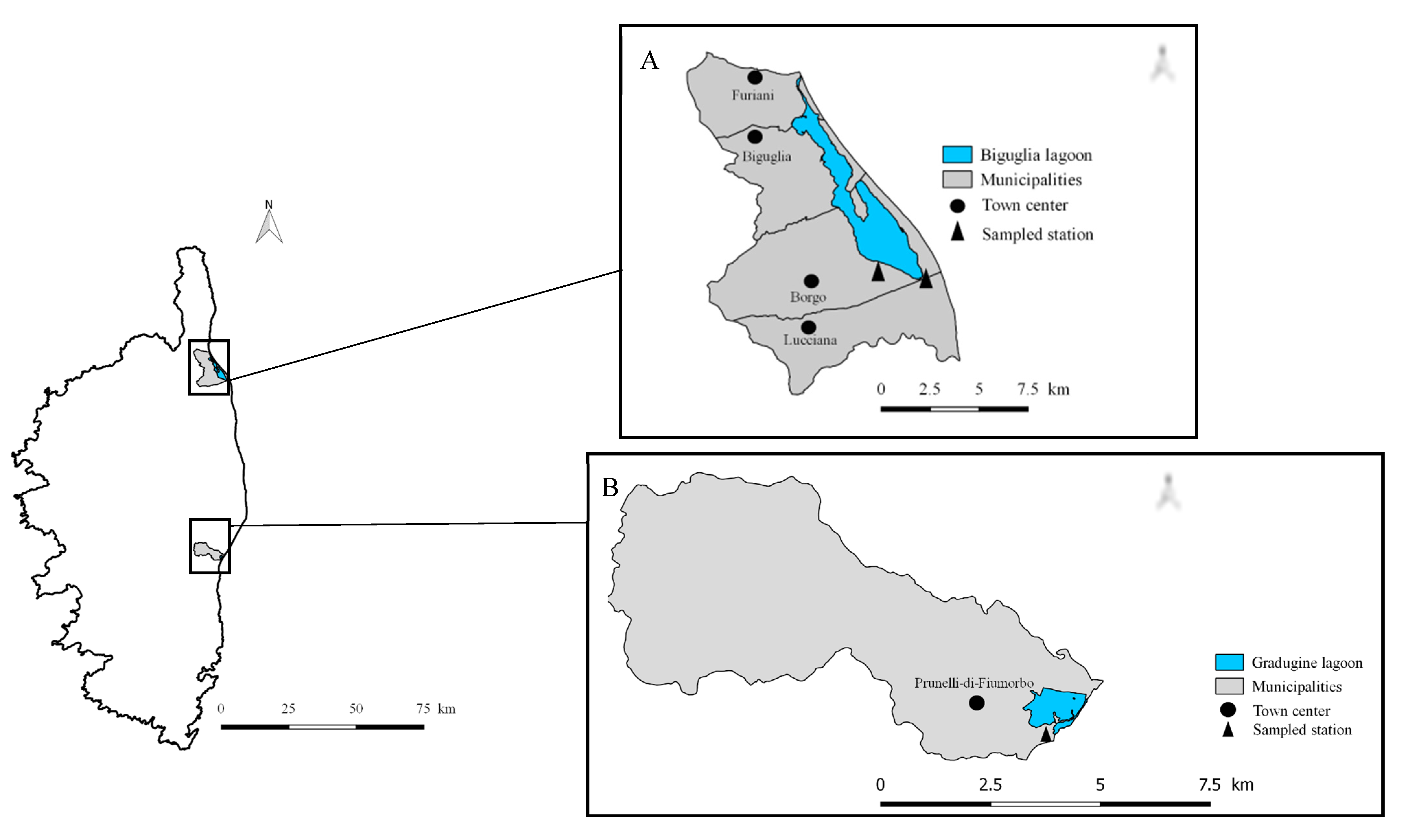
2. Materials and Methods
3. Results
3.1. Ectoparasite Carriers of Pathogens in Birds
3.2. Pathogens Detected in Blood Samples
4. Discussion
4.1. Bird Ectoparasites and Their Pathogens
4.1.1. Louse Flies
4.1.2. Ticks
4.2. Pathogens Found in Birds’ Blood
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmidt, R.; Reavill, D.; Phalen, D. Pathology of Pet and Aviary Birds, 2nd ed.; Wiley Blackwell: Hoboken, NJ, USA, 2015; p. 298. [Google Scholar] [CrossRef]
- Battisti, E.; Urach, K.; Hodžić, A.; Fusani, L.; Hufnagl, P.; Felsberger, G.; Ferroglio, E.; Duscher, G.G. Zoonotic Pathogens in Ticks from Migratory Birds, Italy. Emerg. Infect. Dis. 2020, 26, 2986–2988. [Google Scholar] [CrossRef] [PubMed]
- Klaus, C.; Gethmann, J.; Hoffmann, B.; Ziegler, U.; Heller, M.; Beer, M. Tick infestation in birds and prevalence of pathogens in ticks collected from different places in Germany. Parasitol. Res. 2016, 115, 2729–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liechti, B.B.E.F. Bird migration across the Mediterranean. Proc. Int. Ornithol. Congr. 1999, 22, 1983–1999. [Google Scholar]
- Mellone, U. Sea crossing as a major determinant for the evolution of migratory strategies in soaring birds. J. Anim. Ecol. 2020, 89, 1298–1301. [Google Scholar] [CrossRef]
- Maggini, I.; Cardinale, M.; Sundberg, J.H.; Spina, F.; Fusani, L. Recent phenological shifts of migratory birds at a Mediterranean spring stopover site: Species wintering in the Sahel advance passage more than tropical winterers. PLoS ONE 2020, 15, e0239489. [Google Scholar] [CrossRef]
- Vagliano, C. The Continental and Island Migration Route of the Southeast Mediterranean: Problems and Propositions; ICBP Tech. Publ.: Athens, Greece, 1985; No. 5; pp. 263–268. [Google Scholar]
- Jourdain, E.; Gauthier-Clerc, M.; Bicout, D.J.; Sabatier, P. Bird Migration Routes and Risk for Pathogen Dispersion into Western Mediterranean Wetlands. Emerg. Infect. Dis. 2007, 13, 365–372. [Google Scholar] [CrossRef]
- Bacon, P.R. Use of wetlands for tourism in the insular Caribbean. Ann. Tour. Res. 1987, 14, 104–117. [Google Scholar] [CrossRef]
- Hammer, D.A. Constructed Wetlands for Wastewater Treatment: Municipal, Industrial and Agricultural; CRC Press: Boca Raton, FL, USA, 1989. [Google Scholar]
- Abd Rabou, A.F.N.; Yassin, M.M.; Sharif, F.A.; Al-Agha, M.R.; Daher, K.S.A.; Ali, A.K.S.; Hamad, D.M. Inhabitants Knowledge on Mosquito Proliferation and Control in the Wetland Ecosystem of Wadi Gaza, Gaza Strip, Palestine. IUG J. Nat. Stud. 2020, 16, 2. [Google Scholar]
- Rey, J.R.; Walton, W.E.; Wolfe, R.J.; Connelly, C.R.; O’Connell, S.M.; Berg, J.; Sakolsky-Hoopes, G.E.; Laderman, A.D.; Connelly, R. North American Wetlands and Mosquito Control. Int. J. Environ. Res. Public Health 2012, 9, 4537–4605. [Google Scholar] [CrossRef]
- Dudek, K. Impact of biodiversity on tick-borne diseases. Prz. Epidemiol. 2014, 68, 681–684. [Google Scholar]
- Levison, M.E. Diseases Transmitted by Birds. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Pantin-Jackwood, M.J.; Swayne, D.E. Pathogenesis and pathobiology of avian influenza virus infection in birds. Rev. Sci. Tech. (Int. Off. Epizoot.) 2009, 28, 113–136. [Google Scholar] [CrossRef]
- Hildebrandt, A.; Franke, J.; Meier, F.; Sachse, S.; Dorn, W.; Straube, E. The potential role of migratory birds in transmission cycles of Babesia spp., Anaplasma phagocytophilum, and Rickettsia spp. Ticks Tick-borne Dis. 2010, 1, 105–107. [Google Scholar] [CrossRef]
- O’Donoghue, P. Haemoprotozoa: Making biological sense of molecular phylogenies. Int. J. Parasitol. Parasites Wildl. 2017, 6, 241–256. [Google Scholar] [CrossRef]
- Maquart, M.; Dahmani, M.; Marié, J.-L.; Gravier, P.; Leparc-Goffart, I.; Davoust, B. First Serological Evidence of West Nile Virus in Horses and Dogs from Corsica Island, France. Vector-Borne Zoonotic Dis. 2017, 17, 275–277. [Google Scholar] [CrossRef]
- Vilibic-Cavlek, T.; Petrovic, T.; Savic, V.; Barbic, L.; Tabain, I.; Stevanovic, V.; Klobucar, A.; Mrzljak, A.; Ilic, M.; Bogdanic, M.; et al. Epidemiology of Usutu Virus: The European Scenario. Pathogens 2020, 9, 699. [Google Scholar] [CrossRef]
- Benskin, C.; Wilson, K.; Jones, K.J.; Hartley, I.R. Bacterial pathogens in wild birds: A review of the frequency and effects of infection. Biol. Rev. 2009, 84, 349–373. [Google Scholar] [CrossRef]
- De la Fuente, J.; Estrada-Pena, A.; Venzal, J.M.; Kocan, K.M.; E Sonenshine, D. Overview: Ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 2008, 13, 6938–6946. [Google Scholar] [CrossRef] [Green Version]
- Sonenshine, D.E. Biology of Ticks; Oxford University Press: Oxford, UK, 1991. [Google Scholar]
- Hasle, G. Transport of ixodid ticks and tick-borne pathogens by migratory birds. Front. Cell. Infect. Microbiol. 2013, 3, 48. [Google Scholar] [CrossRef] [Green Version]
- Cicculli, V.; de Lamballerie, X.; Charrel, R.; Falchi, A. First molecular detection of Rickettsia africae in a tropical bont tick, Amblyomma variegatum, collected in Corsica, France. Exp. Appl. Acarol. 2019, 77, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Grech-Angelini, S.; Stachurski, F.; Lancelot, R.; Boissier, J.; Allienne, J.-F.; Marco, S.; Maestrini, O.; Uilenberg, G. Ticks (Acari: Ixodidae) infesting cattle and some other domestic and wild hosts on the French Mediterranean island of Corsica. Parasites Vectors 2016, 9, 1–11. [Google Scholar] [CrossRef]
- Reeves, W.K.; Lloyd, J.E. Louse Flies, Keds, and Bat Flies (Hippoboscoidea). In Medical and Veterinary Entomology, 3rd ed.; Mullen, G.R., Durden, L.A., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 421–438. [Google Scholar] [CrossRef]
- Foley, J.E.; Hasty, J.M.; Lane, R.S. Diversity of rickettsial pathogens in Columbian black-tailed deer and their associated keds (Diptera: Hippoboscidae) and ticks (Acari: Ixodidae). J. Vector Ecol. 2016, 41, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Halos, L.; Jamal, T.; Maillard, R.; Girard, B.; Guillot, J.; Chomel, B.; Vayssier-Taussat, M.; Boulouis, H.-J. Role of Hippoboscidae Flies as Potential Vectors of Bartonella spp. Infecting Wild and Domestic Ruminants. Appl. Environ. Microbiol. 2004, 70, 6302–6305. [Google Scholar] [CrossRef] [Green Version]
- Werszko, J.; Steiner-Bogdaszewska, Ż.; Jeżewski, W.; Szewczyk, T.; Kuryło, G.; Wołkowycki, M.; Wróblewski, P.; Karbowiak, G. Molecular detection of Trypanosoma spp. in Lipoptena cervi and Lipoptena fortisetosa (Diptera: Hippoboscidae) and their potential role in the transmission of pathogens. Parasitology 2020, 147, 1629–1635. [Google Scholar] [CrossRef]
- Žiegytė, R.; Platonova, E.; Bernotienė, R.; Valkiūnas, G.; Palinauskas, V. Complete sporogony of the blood parasite Haemoproteus nucleocondensus in common biting midges: Why is its transmission interrupted in Europe? Parasitology 2020, 147, 593–600. [Google Scholar] [CrossRef]
- Estrada-Peña, A. Ticks of Domestic Animals in the Mediterranean Region: A Guide to Identification of Species; University of Zaragoza: Zaragoza, Spain, 2004. [Google Scholar]
- Pérez-Eid, C. Les Tiques: Identification, Biologie, Importance Médicale et Vétérinaire; Lavoisier: Cachan, France, 2007. [Google Scholar]
- Emichelet, L.; Delannoy, S.; Edevillers, E.; Umhang, G.; Aspán, A.; Ejuremalm, M.; Echirico, J.; van der Wal, F.; Sprong, H.; Pihl, T.; et al. High-throughput screening of tick-borne pathogens in Europe. Front. Cell. Infect. Microbiol. 2014, 4, 103. [Google Scholar] [CrossRef]
- Ehricht, R.; Slickers, P.; Goellner, S.; Hotzel, H.; Sachse, K. Optimized DNA microarray assay allows detection and genotyping of single PCR-amplifiable target copies. Mol. Cell. Probes 2006, 20, 60–63. [Google Scholar] [CrossRef]
- Gondard, M.; Michelet, L.; Nisavanh, A.; Devillers, E.; Delannoy, S.; Fach, P.; Aspan, A.; Ullman, K.; Chirico, J.; Hoffmann, B.; et al. Prevalence of tick-borne viruses in Ixodes ricinus assessed by high-throughput real-time PCR. Pathog. Dis. 2018, 76, fty003. [Google Scholar] [CrossRef] [Green Version]
- Grech-Angelini, S.; Stachurski, F.; Vayssier-Taussat, M.; Devillers, E.; Casabianca, F.; Lancelot, R.; Uilenberg, G.; Moutailler, S. Tick-borne pathogens in ticks (Acari: Ixodidae) collected from various domestic and wild hosts in Corsica (France), a Mediterranean island environment. Transbound. Emerg. Dis. 2020, 67, 745–757. [Google Scholar] [CrossRef]
- Linke, S.; Ellerbrok, H.; Niedrig, M.; Nitsche, A.; Pauli, G. Detection of West Nile virus lineages 1 and 2 by real-time PCR. J. Virol. Methods 2007, 146, 355–358. [Google Scholar] [CrossRef]
- Thibault, J.C.; Bonaccorsi, G. The Birds of Corsica: An Annotated Checklist; British Ornithologists’ Union: Tring, UK, 2022. [Google Scholar]
- Rahola, N.; Goodman, S.; Robert, V. The Hippoboscidae (Insecta: Diptera) from Madagascar, with new records from the “Parc National de Midongy Befotaka”. Parasite 2011, 18, 127–140. [Google Scholar] [CrossRef]
- Oboňa, J.; Sychra, O.; Greš, S.; Heřman, P.; Manko, P.; Roháček, J.; Šestáková, A.; Šlapák, J.; Hromada, M. A revised annotated checklist of louse flies (Diptera, Hippoboscidae) from Slovakia. Zookeys 2019, 862, 129–152. [Google Scholar] [CrossRef]
- Eeva, T.; Andersson, T.; Berglund, M.M.; Brommer, J.E.; Hyvönen, R.; Klemola, T.; Laaksonen, T.; Loukola, O.; Morosinotto, C.; Rainio, K.; et al. Species and abundance of ectoparasitic flies (Diptera) in pied flycatcher nests in Fennoscandia. Parasites Vectors 2015, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Sychra, O.; Literák, I.; Podzemný, P.; Benedikt, V. Insect ectoparasites from wild passerine birds in the Czech Republic. Parasite 2008, 15, 599–604. [Google Scholar] [CrossRef] [Green Version]
- Maa, T.C. A revised checklist and concise host index of Hippoboscidae (Diptera). Pac. Insects Monogr. 1969, 20, 261–299. [Google Scholar]
- la_mouche_plate_n_6.pdf. Consulté Le: 11 Janvier 2022. [En ligne]. Available online: https://crbpo.mnhn.fr/IMG/pdf/la_mouche_plate_n_6.pdf (accessed on 11 January 2022).
- Bezerra-Santos, M.A.; Otranto, D. Keds, the enigmatic flies and their role as vectors of pathogens. Acta Trop. 2020, 209, 105521. [Google Scholar] [CrossRef]
- Buss, M.; Case, L.; Kearney, B.; Coleman, C.; Henning, J. Detection of Lyme disease and anaplasmosis pathogens via PCR in Pennsylvania deer ked. J. Vector Ecol. 2016, 41, 292–294. [Google Scholar] [CrossRef]
- Karadjian, G.; Puech, M.-P.; Duval, L.; Chavatte, J.-M.; Snounou, G.; Landau, I. Haemoproteus syrnii in Strix aluco from France: Morphology, stages of sporogony in a hippoboscid fly, molecular characterization and discussion on the identification of Haemoproteus species. Parasite 2013, 20, 32. [Google Scholar] [CrossRef] [Green Version]
- Mungomba, L.M.; Molyneux, D.H.; Wallbanks, K.R. Host-parasite relationship ofTrypanosoma corvi inOrnithomyia avicularia. Parasitol. Res. 1989, 75, 167–174. [Google Scholar] [CrossRef]
- Santolíková, A.; Brzoňová, J.; Čepička, I.; Svobodová, M. Avian Louse Flies and Their Trypanosomes: New Vectors, New Lineages and Host–Parasite Associations. Microorganisms 2022, 10, 584. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Jameson, L.; Medlock, J.; Vatansever, Z.; Tishkova, F. Unraveling the Ecological Complexities of Tick-Associated Crimean-Congo Hemorrhagic Fever Virus Transmission: A Gap Analysis for the Western Palearctic. Vector-Borne Zoonotic Dis. 2012, 12, 743–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rataud, A.; Henry, P.; Moutailler, S.; Marsot, M. Research effort on birds’ reservoir host potential for Lyme borreliosis: A systematic review and perspectives. Transbound. Emerg. Dis. 2022, 69, 2512–2522. [Google Scholar] [CrossRef]
- Wallménius, K.; Barboutis, C.; Fransson, T.; Jaenson, T.G.; Lindgren, P.-E.; Nyström, F.; Olsen, B.; Salaneck, E.; Nilsson, K. Spotted fever Rickettsia species in Hyalomma and Ixodes ticks infesting migratory birds in the European Mediterranean area. Parasites Vectors 2014, 7, 318. [Google Scholar] [CrossRef] [Green Version]
- Capligina, V.; Salmane, I.; Keišs, O.; Vilks, K.; Japina, K.; Baumanis, V.; Ranka, R. Prevalence of tick-borne pathogens in ticks collected from migratory birds in Latvia. Ticks Tick-borne Dis. 2014, 5, 75–81. [Google Scholar] [CrossRef]
- Baráková, I.; Derdáková, M.; Selyemová, D.; Chvostáč, M.; Špitalská, E.; Rosso, F.; Collini, M.; Rosà, R.; Tagliapietra, V.; Girardi, M.; et al. Tick-borne pathogens and their reservoir hosts in northern Italy. Ticks Tick-borne Dis. 2017, 9, 164–170. [Google Scholar] [CrossRef]
- Klitgaard, K.; Højgaard, J.; Isbrand, A.; Madsen, J.J.; Thorup, K.; Bødker, R. Screening for multiple tick-borne pathogens in Ixodes ricinus ticks from birds in Denmark during spring and autumn migration seasons. Ticks Tick-borne Dis. 2019, 10, 546–552. [Google Scholar] [CrossRef]
- Keskin, A.; Erciyas-Yavuz, K. A Preliminary Investigation on Ticks (Acari: Ixodidae) Infesting Birds in Kızılırmak Delta, Turkey: Table 1. J. Med. Èntomol. 2015, 53, 217–220. [Google Scholar] [CrossRef]
- Peña, A.E.; Osácar, J.J.; Pichon, B.; Gray, J.S. Hosts and Pathogen Detection for Immature Stages of Ixodes ricinus (Acari: Ixodidae) in North-Central Spain. Exp. Appl. Acarol. 2005, 37, 257–268. [Google Scholar] [CrossRef]
- Palomar, A.M.; Portillo, A.; Santibáñez, P.; Mazuelas, D.; Roncero, L.; Gutiérrez, Ó.; Oteo, J.A. Presence ofBorrelia turdiandBorrelia valaisiana(Spirochaetales: Spirochaetaceae) in Ticks Removed From Birds in the North of Spain, 2009–2011. J. Med. Èntomol. 2016, 54, 243–246. [Google Scholar] [CrossRef]
- Ogden, N.H.; Lindsay, L.R.; Hanincová, K.; Barker, I.K.; Bigras-Poulin, M.; Charron, D.F.; Heagy, A.; Francis, C.M.; O’Callaghan, C.J.; Schwartz, I.; et al. Role of Migratory Birds in Introduction and Range Expansion of Ixodes scapularis Ticks and of Borrelia burgdorferi and Anaplasma phagocytophilum in Canada. Appl. Environ. Microbiol. 2008, 74, 1780–1790. [Google Scholar] [CrossRef] [Green Version]
- Stuen, S.; Granquist, E.G.; Silaghi, C. Anaplasma phagocytophilum—A widespread multi-host pathogen with highly adaptive strategies. Front. Cell. Infect. Microbiol. 2013, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Palomar, A.M.; Portillo, A.; Santibáñez, P.; Mazuelas, D.; Roncero, L.; García-Álvarez, L.; Gutiérrez, Ó.; Oteo, J.A. Detection of tick-borne Anaplasma bovis, Anaplasma phagocytophilum and Anaplasma centrale in Spain. Med. Veter-Èntomol. 2015, 29, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Jahfari, S.; Coipan, E.C.; Fonville, M.; van Leeuwen, A.D.; Hengeveld, P.; Heylen, D.; Heyman, P.; van Maanen, C.; Butler, C.M.; Földvári, G.; et al. Circulation of four Anaplasma phagocytophilum ecotypes in Europe. Parasites Vectors 2014, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, E.; Tsao, J.I.; Muñoz, J.D.; Owen, J. Anaplasma phagocytophilum Infection in American Robins and Gray Catbirds: An Assessment of Reservoir Competence and Disease in Captive Wildlife. J. Med. Èntomol. 2013, 50, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, T.; Wilhelmsson, P.; Barboutis, C.; Fransson, T.; Jaenson, T.G.; Lindgren, P.-E.; Von Loewenich, F.D.; Lundkvist, Å.; Olsen, B.; Salaneck, E. A divergent Anaplasma phagocytophilum variant in an Ixodes tick from a migratory bird; Mediterranean basin. Infect. Ecol. Epidemiol. 2020, 10, 1729653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cicculli, V.; DeCarreaux, D.; Ayhan, N.; Casabianca, F.; De Lamballerie, X.; Charrel, R.; Falchi, A. Molecular screening of Anaplasmataceae in ticks collected from cattle in Corsica, France. Exp. Appl. Acarol. 2020, 81, 561–574. [Google Scholar] [CrossRef] [PubMed]
- Hauck, D.; Jordan, D.; Springer, A.; Schunack, B.; Pachnicke, S.; Fingerle, V.; Strube, C. Transovarial transmission of Borrelia spp., Rickettsia spp. and Anaplasma phagocytophilum in Ixodes ricinus under field conditions extrapolated from DNA detection in questing larvae. Parasites Vectors 2020, 13, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madison-Antenucci, S.; Kramer, L.D.; Gebhardt, L.L.; Kauffman, E. Emerging Tick-Borne Diseases. Clin. Microbiol. Rev. 2020, 33, e00083-18. [Google Scholar] [CrossRef]
- Paddock, C.D.; Childs, J.E. Ehrlichia chaffeensis: A Prototypical Emerging Pathogen. Clin. Microbiol. Rev. 2003, 16, 37–64. [Google Scholar] [CrossRef] [Green Version]
- Špitalská, E.; Literak, I.; Sparagano, O.A.E.; Golovchenko, M.; Kocianová, E. Ticks (Ixodidae) from passerine birds in the Carpathian region. Wien. Klin. Wochenschr. 2006, 118, 759–764. [Google Scholar] [CrossRef]
- Toma, L.; Mancini, F.; DI Luca, M.; Cecere, J.G.; Bianchi, R.; Khoury, C.; Quarchioni, E.; Manzia, F.; Rezza, G.; Ciervo, A. Detection of Microbial Agents in Ticks Collected from Migratory Birds in Central Italy. Vector-Borne Zoonotic Dis. 2014, 14, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Påhlson, C.; Lu, X.; Ott, M.; Nilsson, K. Characteristics of in vitro infection of human monocytes, by Rickettsia helvetica. Microbes Infect. 2020, 23, 104776. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.-E.; et al. Update on Tick-Borne Rickettsioses around the World: A Geographic Approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biernat, B.; Stańczak, J.; Michalik, J.; Sikora, B.; Cieniuch, S. Rickettsia helvetica and R. monacensis infections in immature Ixodes ricinus ticks derived from sylvatic passerine birds in west-central Poland. Parasitol. Res. 2016, 115, 3469–3477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elfving, K.; Olsen, B.; Bergström, S.; Waldenström, J.; Lundkvist, Å.; Sjöstedt, A.; Mejlon, H.; Nilsson, K. Dissemination of Spotted Fever Rickettsia Agents in Europe by Migrating Birds. PLoS ONE 2010, 5, e8572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heylen, D.; Fonville, M.; Van Leeuwen, A.D.; Stroo, A.; Duisterwinkel, M.; Van Wieren, S.; Diuk-Wasser, M.; De Bruin, A.; Sprong, H. Pathogen communities of songbird-derived ticks in Europe’s low countries. Parasites Vectors 2017, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Kováts, D.; Csörgő, T.; Meli, M.L.; Gönczi, E.; Hadnagy, Z.; Takács, N.; Farkas, R.; Hofmann-Lehmann, R. Birds as potential reservoirs of tick-borne pathogens: First evidence of bacteraemia with Rickettsia helvetica. Parasites Vectors 2014, 7, 128. [Google Scholar] [CrossRef] [Green Version]
- Hornok, S.; Boldogh, S.A.; Takács, N.; Juhász, A.; Kontschán, J.; Földi, D.; Koleszár, B.; Morandini, P.; Gyuranecz, M.; Szekeres, S. Anaplasmataceae closely related to Ehrlichia chaffeensis and Neorickettsia helminthoeca from birds in Central Europe, Hungary. Antonie Van Leeuwenhoek 2020, 113, 1067–1073. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Liu, Z.; Niu, Q.; Tian, Z.; Liu, J.; Guan, G.; Liu, G.; Luo, J.; Wang, X.; Yin, H. Tick-borne zoonotic pathogens in birds in Guangxi, Southwest China. Parasites Vectors 2015, 8, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.L.; Ross, T.M.; Evans, J.D. West Nile Virus. Clin. Lab. Med. 2010, 30, 47–65. [Google Scholar] [CrossRef]
- Bakonyi, T.; Haussig, J.M. West Nile virus keeps on moving up in Europe. Eurosurveillance 2020, 25, 2001938. [Google Scholar] [CrossRef] [PubMed]
- Zeller, H.G.; Schuffenecker, I. West Nile Virus: An Overview of Its Spread in Europe and the Mediterranean Basin in Contrast to Its Spread in the Americas. Eur. J. Clin. Microbiol. Infect. Dis. 2004, 23, 147–156. [Google Scholar] [CrossRef]
- Pallari, C.T.; Efstathiou, A.; Moysi, M.; Papanikolas, N.; Christodoulou, V.; Mazeris, A.; Koliou, M.; Kirschel, A.N. Evidence of West Nile virus seropositivity in wild birds on the island of Cyprus. Comp. Immunol. Microbiol. Infect. Dis. 2020, 74, 101592. [Google Scholar] [CrossRef] [PubMed]
- Magurano, F.; Remoli, M.E.; Baggieri, M.; Fortuna, C.; Marchi, A.; Fiorentini, C.; Bucci, P.; Benedetti, E.; Ciufolini, M.G.; Rizzo, C.; et al. Circulation of West Nile virus lineage 1 and 2 during an outbreak in Italy. Clin. Microbiol. Infect. 2012, 18, E545–E547. [Google Scholar] [CrossRef] [Green Version]
- Fotakis, E.A.; Mavridis, K.; Kampouraki, A.; Balaska, S.; Tanti, F.; Vlachos, G.; Gewehr, S.; Mourelatos, S.; Papadakis, A.; Kavalou, M.; et al. Mosquito population structure, pathogen surveillance and insecticide resistance monitoring in urban regions of Crete, Greece. PLoS Neglected Trop. Dis. 2022, 16, e0010186. [Google Scholar] [CrossRef] [PubMed]
- Beck, C.; Goffart, I.L.; Franke, F.; Gonzalez, G.; Dumarest, M.; Lowenski, S.; Blanchard, Y.; Lucas, P.; Lamballerie, X.; Grard, G.; et al. Contrasted Epidemiological Patterns of West Nile Virus Lineages 1 and 2 Infections in France from 2015 to 2019. Pathogens 2020, 9, 908. [Google Scholar] [CrossRef] [PubMed]
- Valkiūnas, G.; Iezhova, T.A. Keys to the avian Haemoproteus parasites (Haemosporida, Haemoproteidae). Malar. J. 2022, 21, 1–69. [Google Scholar] [CrossRef]
- Krone, O.; Waldenström, J.; Valkiūnas, G.; Lessow, O.; Müller, K.; Iezhova, T.A.; Fickel, J.; Bensch, S. Haemosporidian Blood Parasites in European Birds of Prey and Owls. J. Parasitol. 2008, 94, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Londoño, J.; Cardona-Romero, M.; Martínez-Sánchez, E.T.; Ossa-López, P.A.; Pérez-Cárdenas, J.E.; Gonzalez, A.D.; Rivera-Páez, F.A.; Castaño-Villa, G.J. Avian haemosporidian (Haemosporida: Plasmodium and Haemoproteus) in the department of Arauca, Colombian Orinoquia region. Parasitol. Res. 2022, 121, 1775–1787. [Google Scholar] [CrossRef]
- Pellegrino, I.; Ilahiane, L.; Boano, G.; Cucco, M.; Pavia, M.; Prestridge, H.; Voelker, G. Avian Haemosporidian Diversity on Sardinia: A First General Assessment for the Insular Mediterranean. Diversity 2021, 13, 75. [Google Scholar] [CrossRef]


{kind=link}
| Pathogens | Name | Sequence | Length (Nucleotide) | Gene | Gene References |
|---|---|---|---|---|---|
| Aujeszky’s disease virus | Aujv_gp50_F | CTTTATCGAGTACGCCGACTG | 225 | gp50 | Y14834.1 |
| Av_gp50_R | AACGGGCACTCTTGCCCC | ||||
| Av_gp50_P | CAGATCTTTGGGCGCTGCCGGC | ||||
| Haemoproteus spp. | Hae_cytB_F | ATATGCATGCTACTGGTGCTAC | 240 | cytochrome B | AF465579.1 |
| Hae_cytB_R | CAAATCCATGAAACAAGTCCAGG | ||||
| Hae_cytB_P | CGGTTGCACCCCAGAAACTCATTGAC | ||||
| Leptospira spp. | Lep_Lipl32_F | CTCTATGTTTGGATTCCTGCC | 158 | LipL32 | MK541891.1 |
| Lep_Lipl32_R | CCAAGTATCAAACCAATGTGGC | ||||
| Lep_Lipl32_P | ATTGATTTTTCTTCTGGGGTAGCCGCTTTG | ||||
| Leucocytozoon spp. | Leu_cytB_F | GGGTTATGTCTTACCATGGGG | 177 | cytochrome B | KF717066.1 |
| Leu_cytB_R | AATTGCTAGTGCTACGAATGGG | ||||
| Leu_cytB_P | AAATGAGTTTTTGGGGAGCAACCGTTATTAC | ||||
| Plasmodium spp. | Pla_ssrRNA_F | ATATAGAAACTGCGAACGGCTC | 339 | ssu rRNA | MK650620.1 |
| Pla_ssrRNA_R | TTTCTCAGGCTCCCTCTCC | ||||
| Pla_ssrRNA_P | CTCTAATTCCCCGTTACCCGTCATAGC | ||||
| Rickettsia monacensis | Ric_mon_F | CTCGGTGCCGGTACTTTAAAC | 192 | ompB | KU961543.1 |
| Ric_mon_R | GAGCACCGCCAATAAGAGC | ||||
| Ric_mon_P | AGTGCCGATGCAAATACTCCGGTGAC | ||||
| Trypanosoma spp. | Try_18SRNA_F | GTAATTCCAGCTCCAAAAGCG | 178 | 18S rRNA | EU596263.1 |
| Try_18SRNA_R | TCAGGAAGGAACCACTCCC | ||||
| Try_18SRNA_P | ACCTCAAGGGCATGGGTCACCAATCC |
| Pathogens | Name | Sequence | Length (Nucleotide) | Genes | Genes References |
|---|---|---|---|---|---|
| Aujeszky’s disease virus | Aujv_gp50_F2 | AACATCCTCACCGACTTCATG | 158 | gp50 | Y14834.1 |
| Av_gp50_R2 | CTGGTAGAACGGCGTCAGG | ||||
| Av_gp50_P2 | AATCGCATCACGTCCACGCCCCC | ||||
| Haemoproteus spp. | Hae_cytB_F2 | CCTTGGGGTCAAATGAGTTTC | 231 | cytochrome B | AF465579.1 |
| Hae_cytB_R2 | AAGCCGTATCATATCCTAAAGG | ||||
| Hae_cytB_P2 | CCTGGACTTGTTTCATGGATTTGTGGAGG | ||||
| Leucocytozoon spp. | Leu_cytB_F2 | GAGTTTCTGGGGAGCTACTG | 197 | cytochrome B | KU842391.1 |
| Leu_cytB_R2 | GGATTAGTGCTACCTTGAATATG | ||||
| Leu_cytB_P2 | TGAATAAATACAATTGCTAGTGCTACGAATGG | ||||
| Plasmodium spp. | Pla_ssrRNA_F2 | TCGAGTTTCTGACCTATCAGC | 264 | ssrRNA | MK650620.1 |
| Pla_ssrRNA_R2 | AGACTTGCCCTCCAATTGTTAC | ||||
| Pla_ssrRNA_P2 | TGGCCTTGCATTGTTATTTCTTGTCACTACC | ||||
| Trypanosoma spp. | Try_18SRNA_F2 | CAACACGGGGAACTTTACCAG | 141 | 18SRNA | EU596263.1 |
| Try_18SRNA_R2 | ATCCTACTGGGCAGCTTGG | ||||
| Try_18SRNA_P2 | CAGGGTGAGGATTGACAGATTGAGTGTTC |
| Bird Species | Lagoon | Blood Samples | Ixodes spp. Pools/Positive Pools | I. accumanitus/ventalloi Pools/Positive Pools | I. arboricola/lividus Pools/Positive Pools | I. frontalis Pools/Positive Pools | I. ricinus Pools/Positive Pools | O. biloba Individuals/Positive Individuals | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Adult | Nymph | Larva | Adult | Nymph | Larva | Adult | Nymph | Larva | Adult | Nymph | Larva | Adult | Nymph | Larva | Adult | |||
| Sedentary | ||||||||||||||||||
| Parus major | Biguglia, Gradugine | 80/2 WNV‡ | - | - | - | - | - | - | - | 1/0 | - | - | - | - | - | - | - | - |
| Migratory | ||||||||||||||||||
| Acrocephalus arundinaceus | Biguglia, Gradugine | 18 | - | - | - | - | - | - | - | - | 1/0 | - | - | - | - | 1/0 | - | - |
| Hirondo rustica | Biguglia, Gradugine | 177 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 37/1 T‡ |
| Luscinia megarhynchos | Biguglia, Gradugine | 15 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1/1 R‡ | - |
| Prunella modularis | Biguglia, Gradugine | 1 | - | - | - | - | - | - | - | - | - | - | 1/0 | - | - | 1/0 | - | - |
| Sedentary/Migratory | ||||||||||||||||||
| Cettia cetti | Biguglia, Gradugine | 95 | - | - | - | 1/1 Rh‡ | - | - | - | - | - | - | - | - | - | 3/0 | - | - |
| Erithacus rubecula | Biguglia, Gradugine | 68/1 Ec | - | 1/0 | 1/0 | - | - | - | - | - | 1/0 | - | - | 1/1 Rh‡ | 1/0 | 4/1 Rh‡ | 3/1 Ec‡ | - |
| Saxicola rubicola | Gradugine | 1 | - | - | - | - | - | - | - | 1/1 R‡ | - | - | - | - | - | - | - | - |
| Sylvia atricapilla | Biguglia, Gradugine | 148 | - | - | - | - | - | - | - | - | - | - | - | - | - | 1/1 Ap‡ | - | - |
| Turdus merula | Biguglia, Gradugine | 17 | - | - | 1/0 | - | - | - | - | - | - | - | - | - | - | 2/1 R‡ | - | - |
| Total | 762/2 WNV‡, 1 Ec‡ | - | 1/0 | 2/0 | 1/1 Rh‡ | - | - | - | 2/1 R‡ | 2/0 | - | 1/0 | 1/0 | 1/0 | 11/1 Ap‡, 1 R‡, 1 Rh‡ | 4/ 1 Ec‡, 1 R‡ | 37/1 T‡ | |
| Bird Species | Tick Species | Louse Fly Species | Number of Ectoparasites (Number of Infested Birds) | Mean Infestation Intensity | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ixodes spp. | Ixodes accuminatus/ventalloi | Ixodes arboricola/lividus | Ixodes frontalis | Ixodes ricinus | Orythomya. biloba | |||||||||||||
| Adults | Nymphs | Larvae | Adults | Nymphs | Larvae | Adults | Nymphs | Larvae | Adults | Nymphs | Larvae | Adults | Nymphs | Larvae | Adults | |||
| Acrocephalus arundinaceus | - | - | - | - | - | - | - | - | 1 | - | - | - | - | 1 | - | - | 2 (2) | 1.0 |
| Cettia cetti | - | - | - | 1 | - | - | - | - | - | - | - | - | - | 7 | - | - | 8 (3) | 2.66 |
| Erithacus rubecula | - | 1 | 1 | - | - | - | - | - | 2 | - | - | 1 | 1 | 7 | 3 | - | 16 (11) | 1.45 |
| Hirundo rustica | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 37 | 37 (35) | 1.06 |
| Luscinia megarhyncos | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | 1 (1) | 1.0 |
| Parus major | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | 1 (1) | 1.0 |
| Prunella modularis | - | - | - | - | - | - | - | - | - | - | 1 | - | - | 1 | - | - | 2 (2) | 1.0 |
| Saxicola rubicola | - | - | - | - | - | - | - | 10 | - | - | - | - | - | - | - | - | 10 (1) | 10 |
| Sylvia atricapilla | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | 1 (1) | 1 |
| Turdus merula | - | - | 1 | - | - | - | - | - | - | - | - | - | - | 2 | - | - | 3 (2) | 1.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Defaye, B.; Moutailler, S.; Vollot, B.; Galon, C.; Gonzalez, G.; Moraes, R.A.; Leoncini, A.-S.; Rataud, A.; Le Guillou, G.; Pasqualini, V.; et al. Detection of Pathogens and Ticks on Sedentary and Migratory Birds in Two Corsican Wetlands (France, Mediterranean Area). Microorganisms 2023, 11, 869. https://doi.org/10.3390/microorganisms11040869
Defaye B, Moutailler S, Vollot B, Galon C, Gonzalez G, Moraes RA, Leoncini A-S, Rataud A, Le Guillou G, Pasqualini V, et al. Detection of Pathogens and Ticks on Sedentary and Migratory Birds in Two Corsican Wetlands (France, Mediterranean Area). Microorganisms. 2023; 11(4):869. https://doi.org/10.3390/microorganisms11040869
Chicago/Turabian StyleDefaye, Baptiste, Sara Moutailler, Benjamin Vollot, Clémence Galon, Gaëlle Gonzalez, Rayane Amaral Moraes, Antoine-Simon Leoncini, Amalia Rataud, Gilles Le Guillou, Vanina Pasqualini, and et al. 2023. "Detection of Pathogens and Ticks on Sedentary and Migratory Birds in Two Corsican Wetlands (France, Mediterranean Area)" Microorganisms 11, no. 4: 869. https://doi.org/10.3390/microorganisms11040869