
Evaluation of Bacillus paramycoides Strains Isolated from Channa Fish sp. on Growth Performance of Labeo rohita Fingerlings Challenged by Fish Pathogen Aeromonas hydrophila MTCC 12301

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
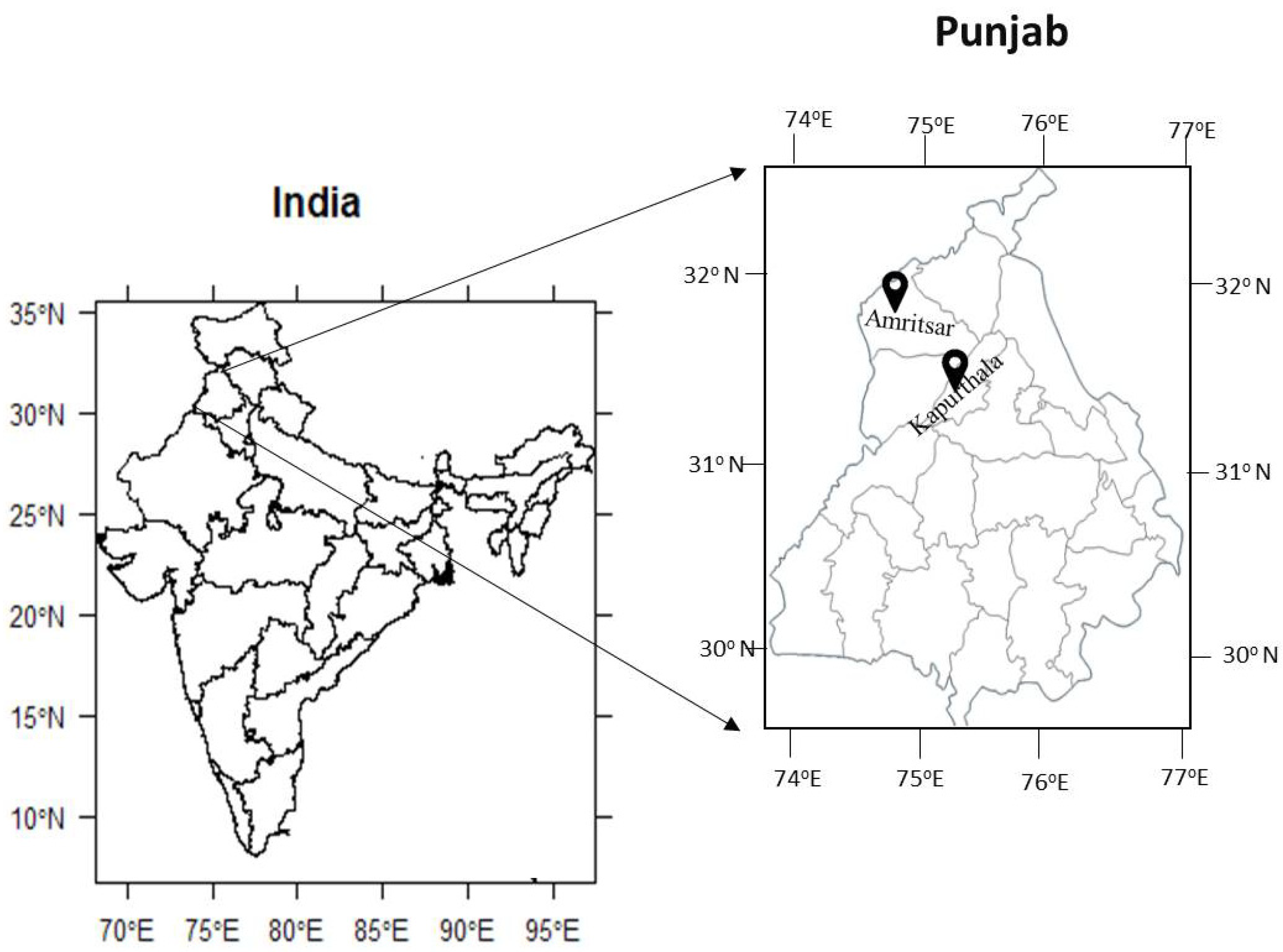
2.1. Sample Collection
2.2. Isolation of Gut Bacteria
2.3. Evaluation of Antimicrobial Activity
2.4. pH and Bile Tolerance
2.5. Antibiotic Susceptibility Test
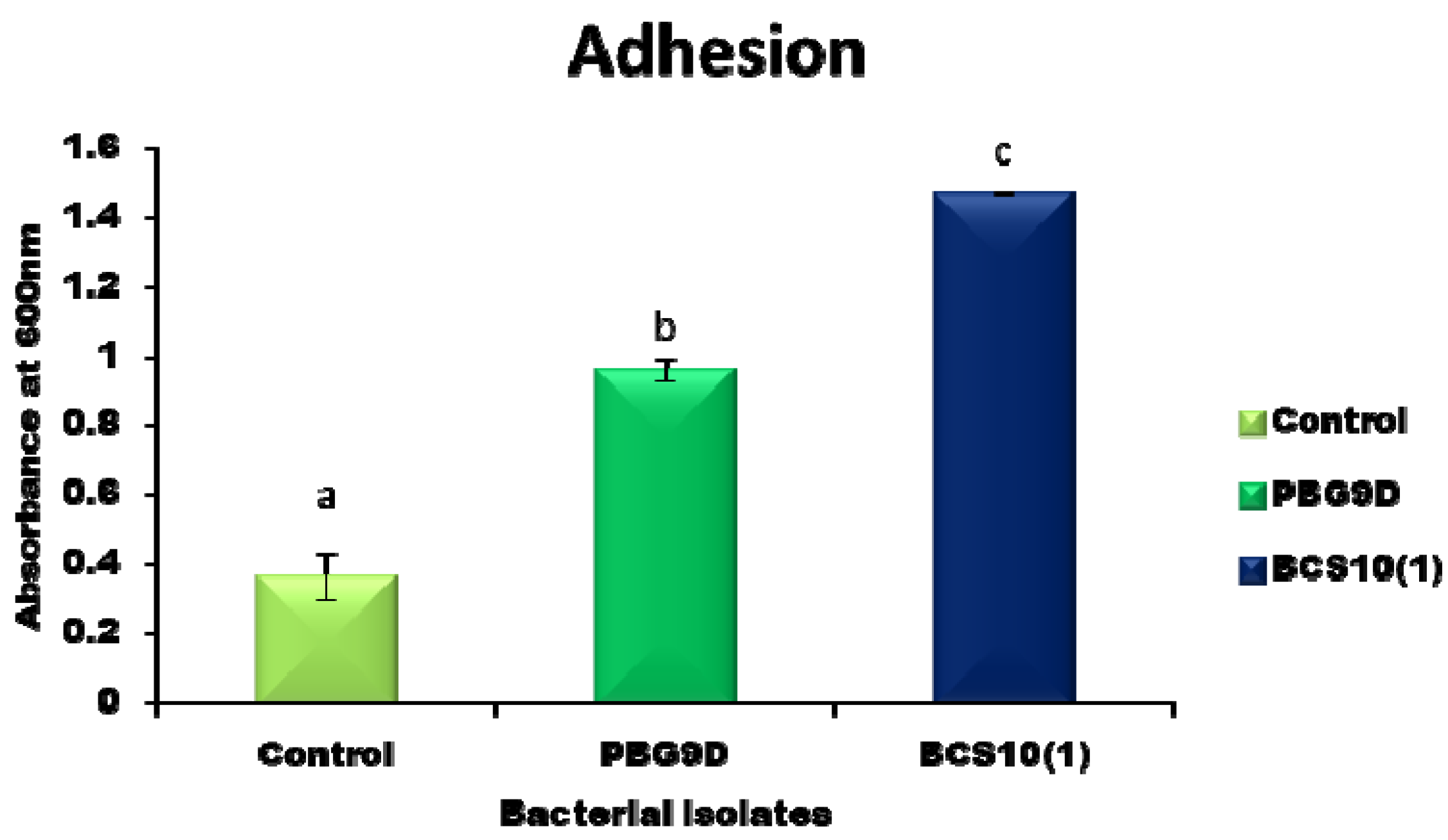
2.6. Adhesion and Biofilm Formation
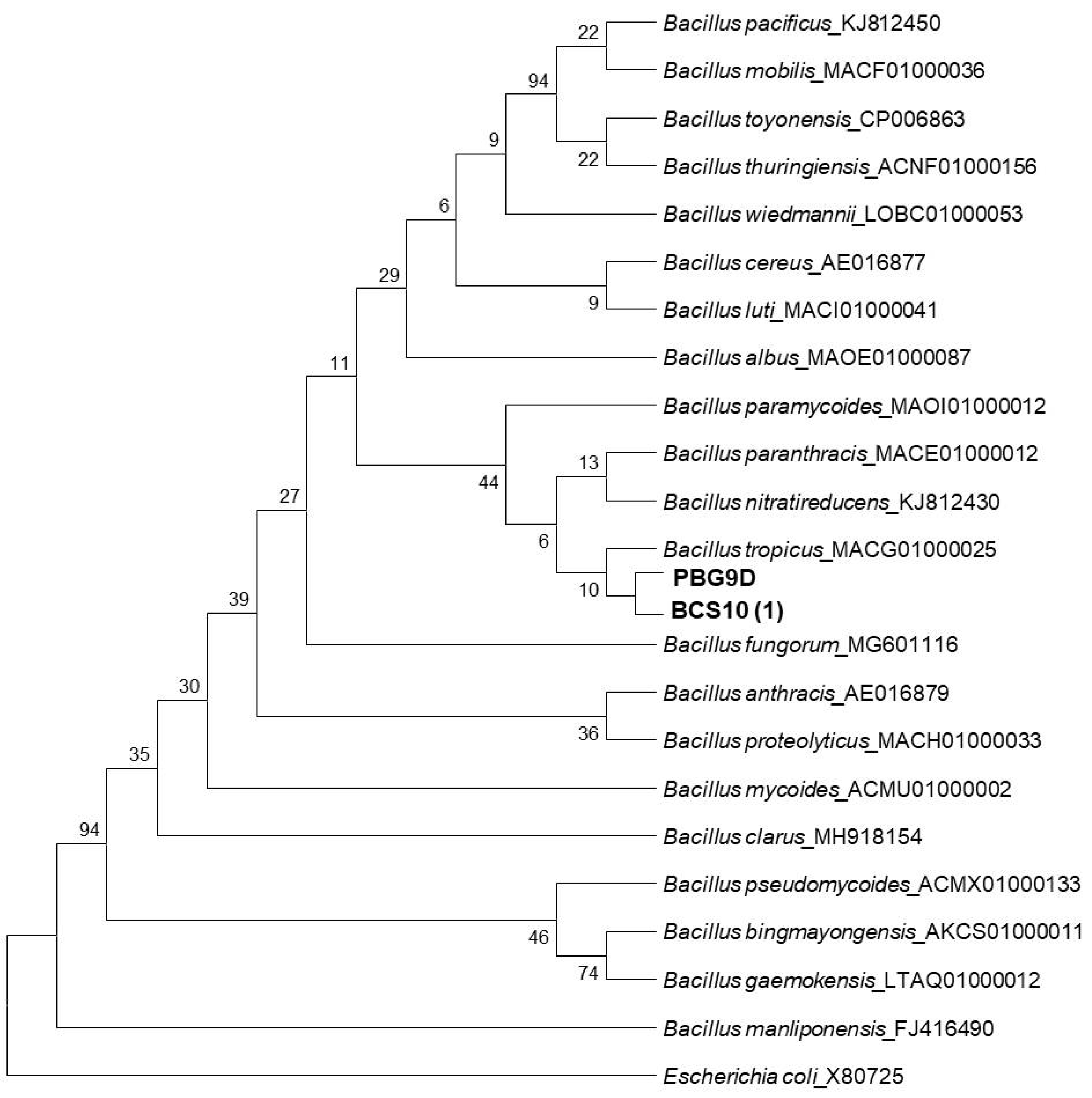
2.7. Identification of Bacterial Isolates
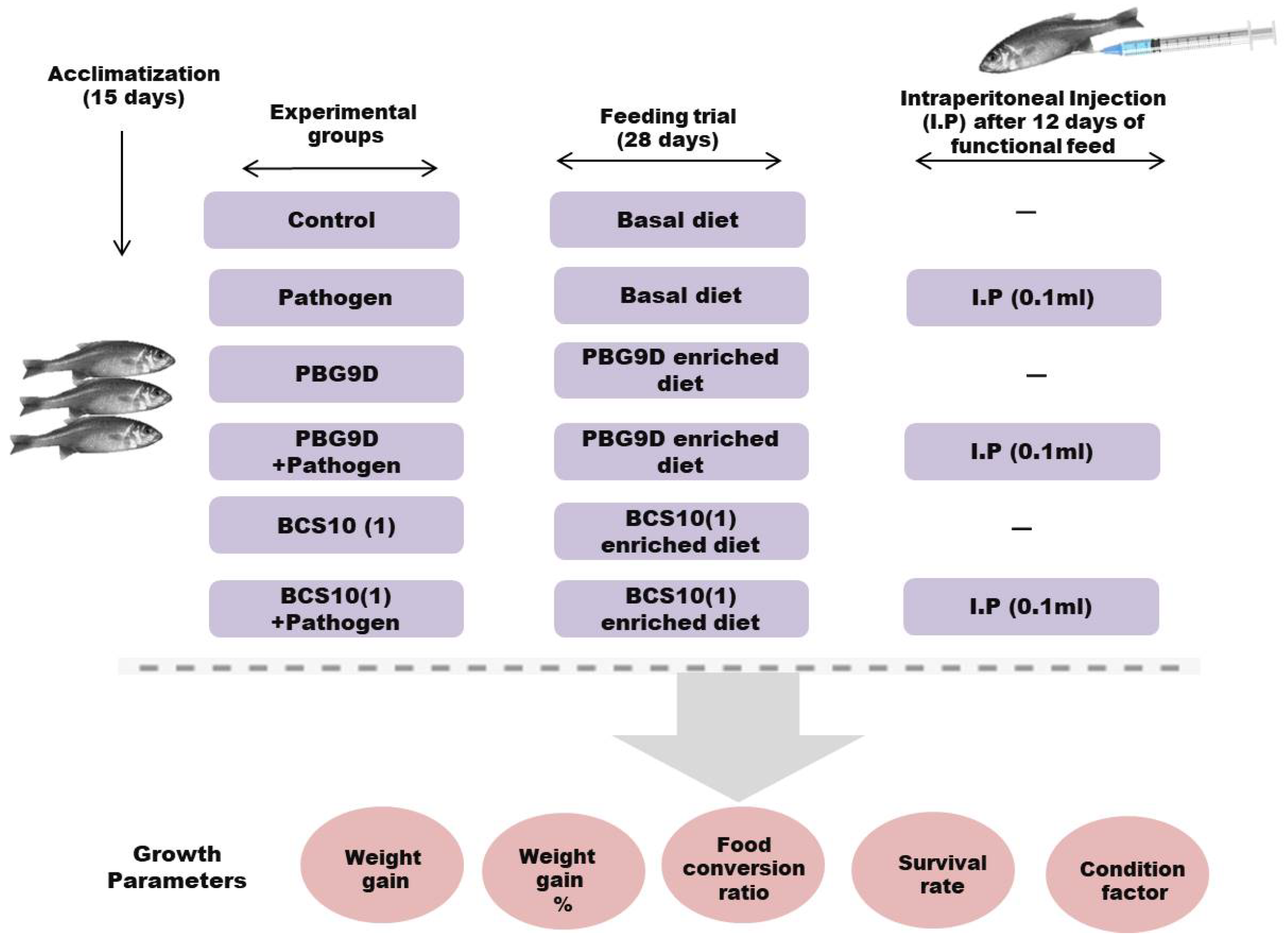
2.8. In Vivo Setup
2.8.1. Preparation of Probiotic Feed
2.8.2. Fish and Experimental Design
2.8.3. Challenge Test
2.8.4. Growth Performances
2.8.5. Statistical Analysis
3. Results
3.1. Isolation of Bacteria and Their Antimicrobial Activity
3.2. pH and Bile Tolerance
3.3. Antibiotic Susceptibility
3.4. Adhesion and Biofilm Formation
3.5. Identification of Isolates
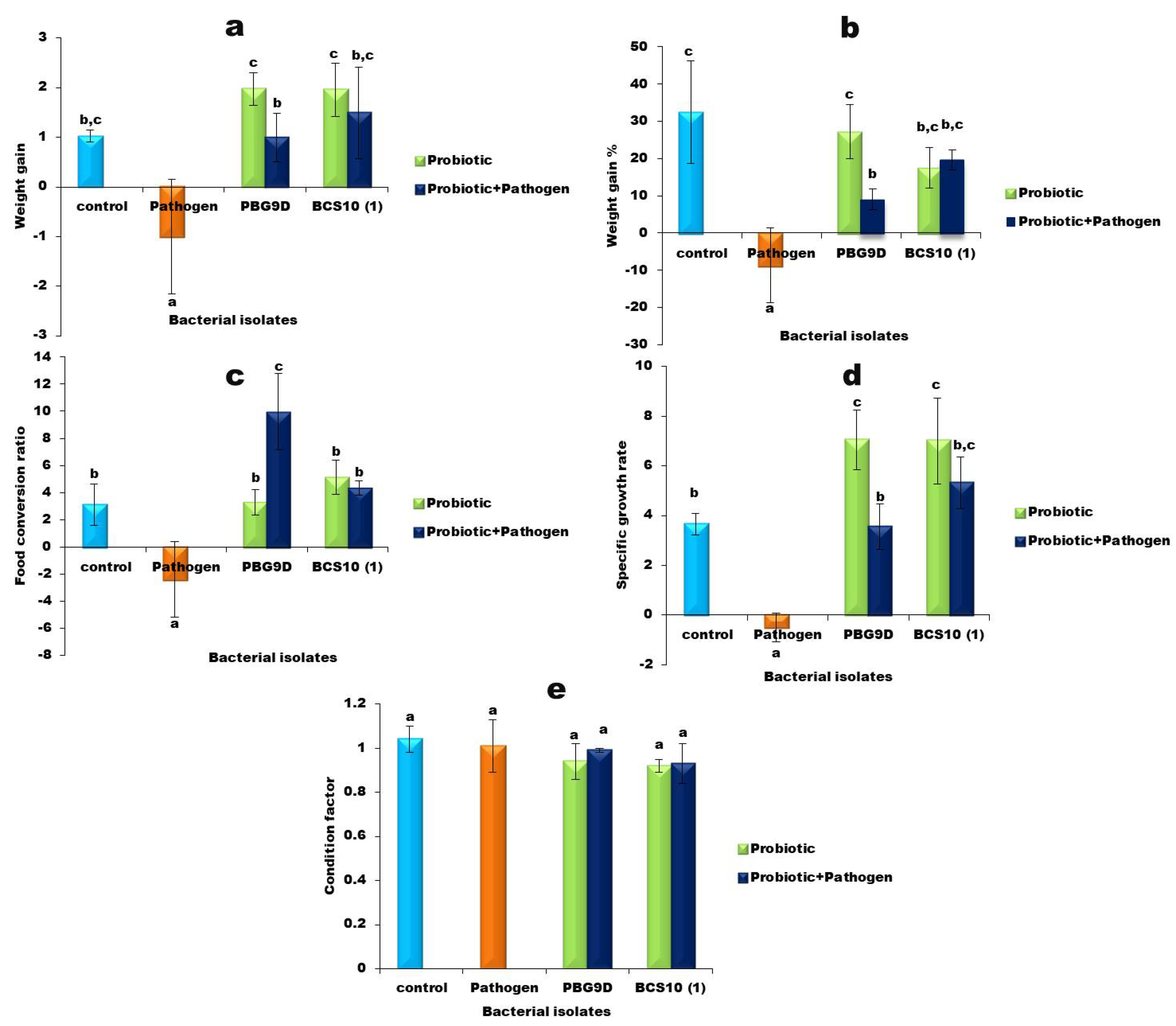
3.6. Growth Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kühlwein, H.; Merrifield, D.L.; Rawling, M.D.; Foey, A.D.; Davies, S.J. Effects of dietary β-(1,3)(1,6)-D-glucan supplementation on growth performance, intestinal morphology and haemato-immunological profile of mirror carp (Cyprinus carpio L.). J. Anim. Physiol. Anim. Nutr. 2014, 98, 279–289. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture; Sustainability in Action: Rome, Italy, 2020. [Google Scholar]
- Nandeesha, M.C.; Sentilkumar, V.; Antony Jesu Prabhu, P. Feed Management of Major Carps in India, with Special Reference to Practices Adopted in Tamil Nadu. In On-Farm Feeding and Feed Management in Aquaculture. FAO Fisheries and Aquaculture Technical Paper; FAO: Rome, Italy, 2013; pp. 433–462. [Google Scholar]
- Majumder, S.; Saikia, S.K. Ecological intensification for feeding rohu Labeo rohita (Hamilton, 1822): A review and proposed steps towards an efficient resource fishery. Aquac. Res. 2020, 51, 3072–3078. [Google Scholar] [CrossRef]
- FAO. Cultured Aquatic Species Information Programme. Labeo rohita; Jena, J.K., Ed.; FAO Fisheries and Aquaculture Department, Food and Agriculture Organization of the United Nations: Rome, Italy, 2006; pp. 1–11. [Google Scholar]
- Bondad-Reantaso, M.G.; Subasinghe, R.P.; Arthur, J.R.; Ogawa, K.; Chinabut, S.; Adlard, R.; Tan, Z.; Shariff, M. Disease and health management in Asian aquaculture. Vet. Parasitol. 2005, 30, 249–272. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, B.R.; Sahoo, P.K. Edwardsiellosis in fish: A brief review. J. Biosci. 2007, 32, 1331–1344. [Google Scholar] [CrossRef] [PubMed]
- Defoirdt, T.; Sorgeloos, P.; Bossier, P. Alternatives to antibiotics for the control of bacterial disease in aquaculture. Curr. Opin. Microbiol. 2011, 14, 251–258. [Google Scholar] [CrossRef]
- Ardura, A.; Linde, A.R.; Garcia-Vazquez, E. Genetic detection of Pseudomonas spp. in commercial amazonian fish. Int. J. Environ. Res. Public Health 2013, 10, 3954–3966. [Google Scholar] [CrossRef]
- Meidong, R.; Khotchanalekha, K.; Doolgindachbaporn, S.; Nagasawa, T.; Nakao, M.; Sakai, K.; Tongpim, S. Evaluation of probiotic Bacillus aerius B81e isolated from healthy hybrid catfish on growth, disease resistance and innate immunity of Pla-mong Pangasius bocourti. Fish Shellfish Immunol. 2018, 73, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Duewelhenke, N.; Krut, O.; Eysel, P. Influence on mitochondria and cytotoxicity of different antibiotics administered in high concentrations on primary human osteoblasts and cell lines. Antimicrob. Agents Chemother. 2007, 51, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Bacanlı, M.; Başaran, N. Importance of antibiotic residues in animal food. Food Chem. Toxicol. 2019, 125, 462–466. [Google Scholar] [CrossRef]
- Okocha, R.C.; Olatoye, I.O.; Adedeji, O.B. Food safety impacts of antimicrobial use and their residues in aquaculture. Public Health Rev. 2018, 39, 21. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, S.; Yilmaz, E.; Dawood, M.A.; Ringø, E.; Ahmadifar, E.; Abdel-Latif, H.M. Probiotics, prebiotics, and synbiotics used to control vibriosis in fish: A review. Aquaculture 2022, 547, 737514. [Google Scholar]
- Perez-Cano, F.J.; Dong, H.; Yaqoob, P. In vitro immunomodulatory activity of Lactobacillus fermentum CECT5716 and Lactobacillus salivarius CECT5713: Two probiotic strains isolated from human breast milk. Immunobiology 2010, 215, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic mechanisms of action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Hoseinifar, S.H.; Sun, Y.Z.; Wang, A.; Zhou, Z. Probiotics as means of diseases control in aquaculture, a review of current knowledge and future perspectives. Front. Microbiol. 2018, 9, 2429. [Google Scholar] [CrossRef] [Green Version]
- Cerezuela, R.; Fumanal, M.; Tapia-Paniagua, S.T.; Meseguer, J.; Moriñigo, M.Á.; Esteban, M.Á. Histological alterations and microbial ecology of the intestine in gilthead seabream (Sparus aurata L.) fed dietary probiotics and microalgae. Cell Tissue Res. 2012, 350, 477–489. [Google Scholar] [PubMed]
- Ibrahem, M.D. Evolution of probiotics in aquatic world: Potential effects, the current status in Egypt and recent prospectives. J. Adv. Res. 2015, 67, 65–91. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Yuan, W.; Wang, S.; Guo, W.; Li, A.; Wu, Y.; Chen, X.; Ren, Z.; Zhou, Y. In vitro screening of putative probiotics and their dual beneficial effects: To white shrimp (Litopenaeus vannamei) postlarvae and to the rearing water. Aquaculture 2019, 498, 61–71. [Google Scholar] [CrossRef]
- Mohammadian, T.; Monjezi, N.; Peyghan, R.; Mohammadian, B. Effects of dietary probiotic supplements on growth, digestive enzymes activity, intestinal histomorphology and innate immunity of common carp (Cyprinus carpio): A field study. Aquaculture 2022, 549, 737787. [Google Scholar] [CrossRef]
- Geng, X.; Dong, X.H.; Tan, B.P.; Yang, Q.H.; Chi, S.Y.; Liu, H.Y.; Liu, X.Q. Effects of dietary probiotic on the growth performance, non-specific immunity and disease resistance of cobia, Rachycentron canadum. Aquac. Nutr. 2012, 18, 46–55. [Google Scholar] [CrossRef]
- Lazado, C.C.; Caipang, C.M.; Estante, E.G. Prospects of host-associated microorganisms in fish and penaeids as probiotics with immunomodulatory functions. Fish Shellfish Immunol. 2015, 45, 2–12. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Ringø, E.; Ángeles Esteban, M.; Dadar, M.; Dawood, M.A.; Faggio, C. Host-associated probiotics: A key factor in sustainable aquaculture. Rev. Fish. Sci. Aquac. 2020, 28, 16–42. [Google Scholar] [CrossRef]
- Ramesh, D.; Vinothkanna, A.; Rai, A.K.; Vignesh, V.S. Isolation of potential probiotic Bacillus spp. and assessment of their subcellular components to induce immune responses in Labeo rohita against Aeromonas hydrophila. Fish Shellfish Immunol. 2015, 45, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Govindaraj, K.; Samayanpaulraj, V.; Narayanadoss, V.; Uthandakalaipandian, R. Isolation of lactic acid bacteria from intestine of freshwater fishes and elucidation of probiotic potential for aquaculture application. Probiotics Antimicrob. Proteins 2021, 13, 1598–1610. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.V.; Praveen, N.K.; Joseph, N.; Leo, A.M.; Vijayan, K.K. Isolation and characterization of a novel antimicrobial oxatetracyclo ketone from Bacillus stercoris MBTDCMFRI Ba37 isolated from the tropical estuarine habitats of Cochin. Mol. Biol. Rep. 2021, 48, 1299–1310. [Google Scholar] [CrossRef]
- Karl, J.P.; Hatch, A.M.; Arcidiacono, S.M.; Pearce, S.C.; Pantoja-Feliciano, I.G.; Doherty, L.A.; Soares, J.W. Effects of psychological, environmental and physical stressors on the gut microbiota. Front. Microbiol. 2018, 9, 2013. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.I.; Choudhury, T.G.; Kamilya, D.; Monsang, S.J.; Parhi, J. Characterization of Bacillus spp. isolated from intestine of Labeo rohita—Towards identifying novel probiotics for aquaculture. Aquac. Res. 2021, 52, 822–830. [Google Scholar] [CrossRef]
- Rao, Y.V.; Das, B.K.; Jyotyrmayee, P.; Chakrabarti, R. Effect of Achyranthes aspera on the immunity and survival of Labeo rohita infected with Aeromonas hydrophila. Fish Shellfish Immunol. 2006, 20, 263–273. [Google Scholar]
- Kavitha, M.; Raja, M.; Perumal, P. Evaluation of probiotic potential of Bacillus spp. isolated from the digestive tract of freshwater fish Labeo calbasu (Hamilton, 1822). Aquac. Rep. 2018, 11, 59–69. [Google Scholar] [CrossRef]
- Guo, X.H.; Kim, J.M.; Nam, H.M.; Park, S.Y.; Kim, J.M. Screening lactic acid bacteria from swine origins for multistrain probiotics based on in vitro functional properties. Anaerobe 2010, 16, 321–326. [Google Scholar] [CrossRef]
- Thankappan, B.; Ramesh, D.; Ramkumar, S.; Natarajaseenivasan, K.; Anbarasu, K. Characterization of Bacillus spp. from the gastrointestinal tract of Labeo rohita—Towards to identify novel probiotics against fish pathogens. Appl. Biochem. Biotechnol. 2015, 175, 340–353. [Google Scholar] [CrossRef]
- Chu, W.; Zhou, S.; Zhu, W.; Zhuang, X. Quorum quenching bacteria Bacillus sp. QSI-1 protect zebrafish (Danio rerio) from Aeromonas hydrophila infection. Sci. Rep. 2014, 4, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Claus, D. Genus Bacillus. Bergey’s Man. Syst. Bacteriol. 1986, 2, 1105–1139. [Google Scholar]
- Adorian, T.J.; Jamali, H.; Farsani, H.G.; Darvishi, P.; Hasanpour, S.; Bagheri, T.; Roozbehfar, R. Effects of probiotic bacteria Bacillus on growth performance, digestive enzyme activity, and hematological parameters of Asian sea bass, Lates calcarifer (Bloch). Probiotics Antimicrob. Proteins 2019, 11, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Srisapoome, P.; Areechon, N. Efficacy of viable Bacillus pumilus isolated from farmed fish on immune responses and increased disease resistance in Nile tilapia (Oreochromis niloticus): Laboratory and on-farm trials. Fish Shellfish Immunol. 2017, 67, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Singh, R. Isolation and evaluation of putative probiotic strains from different teleost to prevent Pseudomonas aeruginosa infection in Cyprinus carpio. Aquac. Res. 2019, 50, 3616–3627. [Google Scholar] [CrossRef]
- Asaduzzaman, M.; Sofia, E.; Shakil, A.; Haque, N.F.; Khan, M.N.; Ikeda, D.; Kinoshita, S.; Abol-Munafi, A.B. Host gut-derived probiotic bacteria promote hypertrophic muscle progression and upregulate growth-related gene expression of slow-growing Malaysian Mahseer Tor tambroides. Aquac. Rep. 2018, 9, 37–45. [Google Scholar] [CrossRef]
- Ramesh, D.; Souissi, S.; Ahamed, T.S. Effects of the potential probiotics Bacillus aerophilus KADR3 in inducing immunity and disease resistance in Labeo rohita. Fish Shellfish Immunol. 2017, 70, 408–415. [Google Scholar] [CrossRef]
- Tachibana, L.; Telli, G.S.; Dias, D.; Goncalves, G.S.; Ishikawa, C.M.; Cavalcante, R.B.; Natori, M.M.; Hamed, S.B.; Ranzani-Paiva, M.J. Effect of feeding strategy of probiotic Enterococcus faecium on growth performance, hematologic, biochemical parameters and non-specific immune response of Nile tilapia. Aquac. Rep. 2020, 16, 100277. [Google Scholar] [CrossRef]
- Sahoo, P.K.; Rauta, P.R.; Mohanty, B.R.; Mahapatra, K.D.; Saha, J.N.; Rye, M.; Eknath, A.E. Selection for improved resistance to Aeromonas hydrophila in Indian major carp Labeo rohita: Survival and innate immune responses in first generation of resistant and susceptible lines. Fish Shellfish Immunol. 2011, 31, 432–438. [Google Scholar] [CrossRef]
- Algammal, A.M.; Mabrok, M.; Sivaramasamy, E.; Youssef, F.M.; Atwa, M.H.; El-Kholy, A.W.; Hetta, H.F.; Hozzein, W.N. Emerging MDR-Pseudomonas aeruginosa in fish commonly harbor oprL and toxA virulence genes and blaTEM, blaCTX-M, and tetA antibiotic-resistance genes. Sci. Rep. 2020, 10, 1–2. [Google Scholar] [CrossRef]
- Dawood, M.A.; Abo-Al-Ela, H.G.; Hasan, M.T. Modulation of transcriptomic profile in aquatic animals: Probiotics, prebiotics and synbiotics scenarios. Fish Shellfish Immunol. 2020, 97, 268–282. [Google Scholar] [CrossRef]
- Ahire, J.J.; Kashikar, M.S.; Lakshmi, S.G.; Madempudi, R. Identification and characterization of antimicrobial peptide produced by indigenously isolated Bacillus paralicheniformis UBBLi30 strain. 3 Biotech. 2020, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Cherif, A.; Ouzari, H.; Daffonchio, D.; Cherif, H.; Ben Slama, K.; Hassen, A.; Jaoua, S.; Boudabous, A. Thuricin 7: A novel bacteriocin produced by Bacillus thuringiensis BMG1. 7, a new strain isolated from soil. Lett. Appl. Microbiol. 2001, 32, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Yousuf, S.; Tyagi, A.; Singh, R. Probiotic supplementation as an emerging alternative to chemical therapeutics in finfish aquaculture: A Review. Probiotics Antimicrob. Proteins 2022. Online ahead of print. [Google Scholar] [CrossRef]
- FAO. Aquaculture Feed and Fertilizer Resources Information System. 2020. Available online: https://www.fao.org/fishery/affris/species-profiles/roho-labeo/rohu-home/en/ (accessed on 31 October 2021).
- Panigrahi, A.; Azad, I.S. Microbial intervention for better fish health in aquaculture: The Indian scenario. Fish Physiol. Biochem. 2007, 33, 429–440. [Google Scholar] [CrossRef]
- Kos, B.V.; Šušković, J.; Vuković, S.; Šimpraga, M.; Frece, J.; Matošić, S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikoskelainen, S.; Ouwehand, A.C.; Bylund, G.; Salminen, S.; Lilius, E.M. Immune enhancement in rainbow trout (Oncorhynchus mykiss) by potential probiotic bacteria (Lactobacillus rhamnosus). Fish Shellfish Immunol. 2003, 15, 443–452. [Google Scholar] [CrossRef]
- Rojas, M.; Ascencio, F.; Conway, P.L. Purification and characterization of a surface protein from Lactobacillus fermentum 104R that binds to porcine small intestinal mucus and gastric mucin. Appl. Environ. Microbiol. 2002, 68, 2330–2336. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.H.; Rahimnejad, S.; Yang, S.Y.; Kim, K.W.; Lee, K.J. Evaluations of Bacillus spp. as dietary additives on growth performance, innate immunity and disease resistance of olive flounder (Paralichthys olivaceus) against Streptococcus iniae and as water additives. Aquaculture 2013, 402, 50–57. [Google Scholar] [CrossRef]
- Mukherjee, A.; Chandra, G.; Ghosh, K. Single or conjoint application of autochthonous Bacillus strains as potential probiotics: Effects on growth, feed utilization, immunity and disease resistance in Rohu, Labeo rohita (Hamilton). Aquaculture 2019, 512, 734302. [Google Scholar] [CrossRef]
- Pant, B.; Lohani, V.; Mishra, A.; Trakroo, M.D.; Tewari, H. Effect of probiotic supplementation on growth of carp fingerlings. Natl. Acad. Sci. Lett. 2019, 42, 215–220. [Google Scholar] [CrossRef]
- Shadrack, R.S.; Manabu, I.; Yokoyama, S. Efficacy of single and mix probiotic bacteria strain on growth indices, physiological condition and bio-chemical composition of juvenile amberjack (Seriola dumerili). Aquac. Rep. 2021, 20, 100753. [Google Scholar] [CrossRef]
- Feng, Z.; Song, X.; Zhao, L.; Zhu, W. Isolation of probiotics and their effects on growth, antioxidant and non-specific immunity of sea cucumber Apostichopus japonicus. Fish Shellfish Immunol. 2020, 106, 1087–1094. [Google Scholar] [CrossRef] [PubMed]
- Mercado, V.; Olmos, J. Bacteriocin production by Bacillus species: Isolation, characterization, and application. Probiotics Antimicrob. Proteins 2022, 14, 1151–1169. [Google Scholar] [CrossRef] [PubMed]
- Ahire, J.J.; Mokashe, N.U.; Chaudhari, B.L. Effect of dietary probiotic Lactobacillus helveticus on growth performance, antioxidant levels, and absorption of essential trace elements in goldfish (Carassius auratus). Probiotics Antimicrob. Proteins 2019, 11, 559–568. [Google Scholar] [CrossRef]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, A.; Hussain, Z.; Akram, A.M.; Alorabi, M.; Sarwar, N.; Rehman, R.A.; Khan, N.A.; Khan, M.F.; Minahal, Q.; El Enshasy, H.A.; et al. Impact of Bacillus subtilis supplemented feed on growth and biochemical constituents in Labeo rohita fingerlings. J. King Saud. Univ. Sci. 2021, 33, 101668. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Isolated Bacterial Strains | ||
|---|---|---|
| Channa punctatus | Channa striatus | Gram positive |
| PBG1, PBG2, PBG3, PBG4, PBG5, PBG6, PBG7A, PBG7B, PBG7C, PBG7D, PBG7E, PBG8, PBG9A, PBG9B, PBG9C, PBG9D, PBG9E, PBG10 | BCS1 (1), BCS1 (2), BCS3, BCS4, BCS5, BCS6, BCS7, BCS8, BCS9, BCS10 (1), BCS10 (2), BCS11, BCS12, BCS13, BCS14, BCS15, BCS16, BCS17, BCS18 (1), BSS18 (2), BCS19, BCS20 (1), BCS20 (2), MCS1, MCS2, MCS3, MCS4, MCS5, MCS6, MCS7, MCS8, MCS9, MCS10, MCS11, MCS12, MCS13, MCS14, MCS15, MCS16, MCS17, MCS18, MCS19 | PBG9D, BCS1 (1), BCS1 (2), BCS10 (1), BCS10 (2), BCS18 (1), BCS19, BCS20 (1), BCS20 (2), MCS6, MCS15, PBG5, PBG1, BCS4, BCS6 |
| S. No | Bacterial Strains | Aeromonas hydrophila | Pseudomonas aeruginosa |
|---|---|---|---|
| 01 | PBG9D | +++ | − |
| 02 | BCS10(1) | ++ | − |
| S. No. | Antibiotics | PBG9D | BCS10 (1) |
|---|---|---|---|
| 01 | Ampicillin (25 µg) | S | S |
| 02 | Neomycin (30 µg) | S | S |
| 03 | Amoxicillin (10 µg) | R | R |
| 04 | Erythromycin (15 µg) | S | S |
| 05 | Tetracycline (30 µg) | S | S |
| 06 | Kanamycin (30 µg) | S | S |
| 07 | Penicillin (10U) | R | R |
| Biochemical Tests | PBG9D | BCS10 (1) |
|---|---|---|
| Gram staining | + | + |
| Motility test | − | − |
| Catalase test | + | + |
| Urease test | − | − |
| Indole test | − | − |
| Gelatin test | − | − |
| Glucose fermentation | + | + |
| Lactose fermentation | − | − |
| Methyl red | + | + |
| Voges–Proskauer | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yousuf, S.; Jamal, M.T.; Al-Farawati, R.K.; Al-Mur, B.A.; Singh, R. Evaluation of Bacillus paramycoides Strains Isolated from Channa Fish sp. on Growth Performance of Labeo rohita Fingerlings Challenged by Fish Pathogen Aeromonas hydrophila MTCC 12301. Microorganisms 2023, 11, 842. https://doi.org/10.3390/microorganisms11040842
Yousuf S, Jamal MT, Al-Farawati RK, Al-Mur BA, Singh R. Evaluation of Bacillus paramycoides Strains Isolated from Channa Fish sp. on Growth Performance of Labeo rohita Fingerlings Challenged by Fish Pathogen Aeromonas hydrophila MTCC 12301. Microorganisms. 2023; 11(4):842. https://doi.org/10.3390/microorganisms11040842
Chicago/Turabian StyleYousuf, Sufiara, Mamdoh T. Jamal, Radwan Kahalid Al-Farawati, Bandar Ahmad Al-Mur, and Rahul Singh. 2023. "Evaluation of Bacillus paramycoides Strains Isolated from Channa Fish sp. on Growth Performance of Labeo rohita Fingerlings Challenged by Fish Pathogen Aeromonas hydrophila MTCC 12301" Microorganisms 11, no. 4: 842. https://doi.org/10.3390/microorganisms11040842