
Antibiotic-Resistant Desulfovibrio Produces H2S from Supplements for Animal Farming
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Physiological Tests
2.2. Sequencing of Desulfovibrio vulgaris L2 Genome
3. Results
3.1. Desulfovibrio vulgaris L2 Physiology and Resistance to Antibiotics
3.2. General Genome Properties
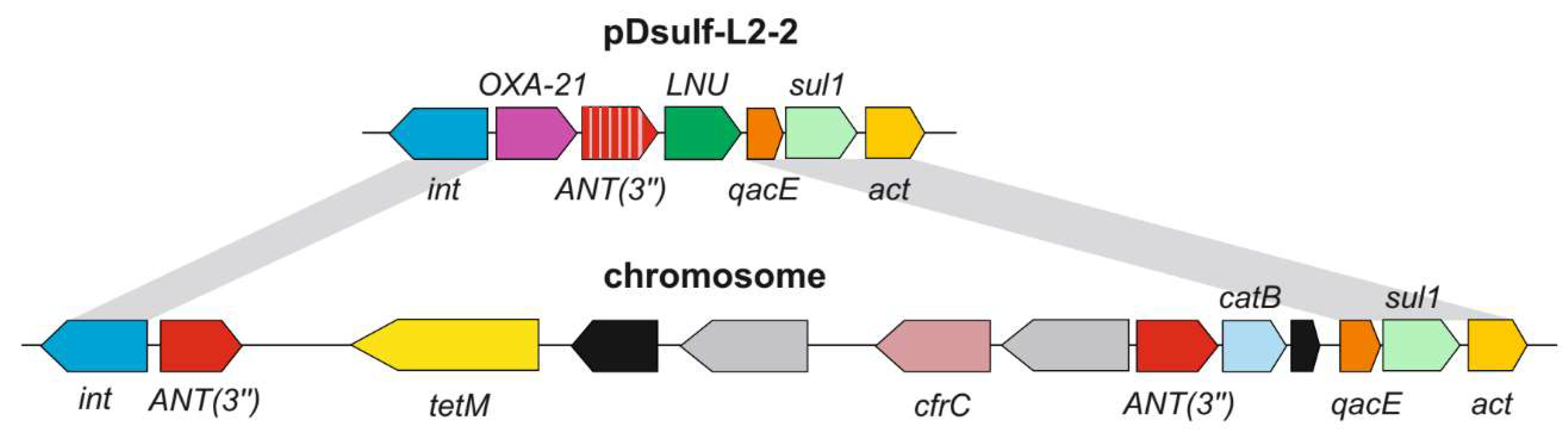
3.3. Genetic Determinants of Antibiotic Resistance
3.4. Genetic Determinants of Resistance to Mercury
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fox, J.G.; Dewhirst, F.E.; Fraser, G.J.; Paster, B.J.; Shames, B.; Murphy, J.C. Intracellular Campylobacter-like organism from ferrets and hamsters with proliferative bowel disease is a Desulfovibrio sp. J. Clin. Microbiol. 1994, 32, 1229–1237. [Google Scholar] [CrossRef] [Green Version]
- Beerens, H.; Romond, C. Sulfate-reducing anaerobic bacteria in human feces. Am. J. Clin. Nutr. 1977, 30, 1770–1776. [Google Scholar] [CrossRef]
- Gibson, G.R.; Cummings, J.H.; Macfarlane, G.T. Competition for hydrogen between sulphate-reducing bacteria and methanogenic bacteria from the human large intestine. J. Appl. Bacteriol. 1988, 65, 241–247. [Google Scholar] [CrossRef]
- Goldstein, E.J.; Citron, D.M.; Peraino, V.A.; Cross, S.A. Desulfovibrio desulfuricans bacteremia and review of human Desulfovibrio infections. J. Clin. Microbiol. 2003, 41, 2752–2754. [Google Scholar] [CrossRef] [Green Version]
- Fite, A.; Macfarlane, G.T.; Cummings, J.H.; Hopkins, M.J.; Kong, S.C.; Furrie, E.; Macfarlane, S. Identification and quantitation of mucosal and faecal desulfovibrios using real time polymerase chain reaction. Gut 2004, 53, 523–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.L.; Yin, X.C.; Long, W.M.; Fei, N.; Zhao, L.P.; Pang, X.Y. Development of a group-specific PCR-based DGGE for analyzing sulfate-reducing bacteria in human gut. Chin. J. Microbiol. 2013, 25, 706–711. [Google Scholar]
- Rey, F.E.; Gonzalez, M.D.; Cheng, J.; Wu, M.; Ahern, P.P.; Gordon, J.I. Metabolic niche of a prominent sulfate-reducing human gut bacterium. Proc. Nat. Acad. Sci. USA 2013, 110, 13582–13587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, L.L.; Ritz, N.L.; Fauque, G.D.; Lin, H.C. Sulfur Cycling and the Intestinal Microbiome. Dig. Dis. Sci. 2017, 62, 2241–2257. [Google Scholar] [CrossRef]
- Pitcher, M.C.; Beatty, E.R.; Cummings, J.H. The contribution of sulphate reducing bacteria and 5-aminosalicylic acid to faecal sulphide in patients with ulcerative colitis. Gut 2000, 46, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Fiachra, R.; Docherty, N.G.; Madeline, M.T.; Brendan, M.J.; Calvin, C.; O’Connell, P.R. Bacterial colonization of colonic crypt mucous gel and disease activity in ulcerative colitis. Ann. Surg. 2010, 252, 869–875. [Google Scholar]
- Loubinoux, J.; Bronowicki, J.P.; Pereira, I.A.; Mougenel, J.L.; Faou, A.E. Sulfate-reducing bacteria in human feces and their association with inflammatory bowel diseases. FEMS Microbiol. Ecol. 2002, 40, 107–112. [Google Scholar] [CrossRef]
- Scanlan, P.D.; Shanahan, F.; Marchesi, J.R. Culture-independent analysis of desulfovibrios in the human distal colon of healthy, colorectal cancer and polypectomized individuals. FEMS Microbiol. Ecol. 2009, 69, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Wolf, P.G.; Cowley, E.S.; Breister, A.; Matatov, S.; Lucio, L.; Polak, P.; Ridlon, J.M.; Gaskins, H.R.; Anantharaman, K. Diversity and distribution of sulfur metabolic genes in the human gut microbiome and their association with colorectal cancer. Microbiome 2022, 10, 64. [Google Scholar] [CrossRef]
- McDougall, R.; Robson, J.; Paterson, D.; Tee, W. Bacteremia caused by a recently described novel Desulfovibrio species. J. Clin. Microbiol. 1997, 35, 1805–1808. [Google Scholar] [CrossRef] [Green Version]
- Loubinoux, J.; Mory, F.; Pereira, I.A.C.; Le Faou, A.E. Bacteremia caused by a strain of Desulfovibrio related to the provisionally named Desulfovibrio fairfieldensis. J. Clin. Microbiol. 2000, 38, 931–934. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.K.; Reed, K.D. Desulfovibrio desulfuricans bacteremia in a dog. J. Clin. Microbiol. 2000, 38, 1701–1702. [Google Scholar] [CrossRef] [Green Version]
- Hagiya, H.; Kimura, K.; Nishi, I.; Yamamoto, N.; Yoshida, H.; Akeda, Y.; Tomono, K. Desulfovibrio desulfuricans bacteremia: A case report and literature review. Anaerobe 2018, 49, 112–115. [Google Scholar] [CrossRef]
- Lozniewski, A.; Maurer, P.; Schumacher, H.; Carlier, J.P.; Mory, F. First isolation of Desulfovibrio species as part of a polymicrobial infection from a brain abscess. Eur. J. Clin. Microbiol. Infect. Dis. 1999, 18, 602–603. [Google Scholar] [CrossRef]
- Okamoto, Y.; Miyabe, Y.; Seki, M.; Ushio, Y.; Sato, K.; Kasama, E.; Akiyama, K.; Karasawa, K.; Uchida, K.; Kikuchi, K.; et al. First case of a renal cyst infection caused by Desulfovibrio: A case report and literature review. BMC Nephrol. 2022, 23, 194. [Google Scholar] [CrossRef]
- Lin, Y.C.; Lin, H.F.; Wu, C.C.; Chen, C.L.; Ni, Y.H. Pathogenic effects of Desulfovibrio in the gut on fatty liver in diet-induced obese mice and children with obesity. J. Gastroenterol. 2022, 57, 913–925. [Google Scholar] [CrossRef]
- Vasoo, S.; Mason, E.L.; Gustafson, D.R.; Cunningham, S.A.; Cole, N.C.; Vetter, E.A.; Steinmann, S.P.; Wilson, W.R.; Patel, R.; Berbari, E.F.; et al. Desulfovibrio legallii prosthetic shoulder joint infection and review of antimicrobial susceptibility and clinical characteristics of Desulfovibrio infections. J. Clin. Microbiol. 2014, 52, 3105–3110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakao, K.; Tanaka, K.; Ichiishi, S.; Mikamo, H.; Shibata, T.; Watanabe, K. Susceptibilities of 23 Desulfovibrio isolates from humans. Antimicrob. Agents Chemother. 2009, 53, 5308–5311. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A. Plasmids and the spread of resistance. Int. J. Med. Microbiol. 2013, 303, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Postgate, J.R. The Sulphate-Reducing Bacteria, 2nd ed.; Cambridge University Press: London, UK, 1984; Volume 130. [Google Scholar]
- Heidelberg, J.F.; Seshadri, R.; Haveman, S.A.; Hemme, C.L.; Paulsen, I.T.; Kolonay, J.F.; Eisen, J.A.; Ward, N.; Methe, B.; Brinkac, L.M.; et al. The genome sequence of the anaerobic, sulfate-reducing bacterium Desulfovibrio vulgaris Hildenborough. Nat. Biotechnol. 2004, 22, 554–559. [Google Scholar] [CrossRef]
- Castañeda-Carrión, I.N.; Sheik, C.S.; Krumholz, L.R. Desulfovibrio africanus subsp. uniflagellum subsp. nov., a sulfate-reducing bacterium from a uranium-contaminated subsurface aquifer. Int. J. Syst. Evol. Microbiol. 2010, 60, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Castañeda-Carrión, I.N.; Whiteley, M.; Krumholz, L.R. Characterization of pNC1, a small and mobilizable plasmid for use in genetic manipulation of Desulfovibrio africanus. J. Microbiol. Methods 2009, 79, 23–31. [Google Scholar] [CrossRef]
- Rabus, R.; Venceslau, S.S.; Wöhlbrand, L.; Voordouw, G.; Wall, J.D.; Pereira, I.A. A Post-Genomic View of the Ecophysiology, Catabolism and Biotechnological Relevance of Sulphate-Reducing Prokaryotes. Adv. Microb. Physiol. 2015, 66, 55–321. [Google Scholar] [CrossRef]
- Karnachuk, O.V.; Rusanov, I.I.; Panova, I.A.; Grigoriev, M.A.; Zyusman, V.S.; Latygolets, E.A.; Kadyrbaev, M.K.; Gruzdev, E.V.; Beletsky, A.V.; Mardanov, A.V.; et al. Microbial sulfate reduction by Desulfovibrio is an important source of hydrogen sulfide from a large swine finishing facility. Sci. Rep. 2021, 11, 10720. [Google Scholar] [CrossRef]
- Karnachuk, O.; Kurochkina, S.; Tuovinen, O. Growth of sulfate-reducing bacteria with solid-phase electron acceptors. Appl. Microbiol. Biotechnol. 2002, 58, 482–486. [Google Scholar] [CrossRef]
- Li, Q.; Gould, S.A.; Htoo, J.K.K.; González-Vega, J.C.; Patience, J.F. Bioavailability of l-lysine sulfate relative to l-lysine HCl for growing-finishing pigs. Transl. Anim. Sci. 2019, 3, 1254–1262. [Google Scholar] [CrossRef] [Green Version]
- Ran, S.; Mu, C.; Zhu, W. Diversity and community pattern of sulfate-reducing bacteria in piglet gut. J. Anim. Sci. Biotechnol. 2019, 10, 40. [Google Scholar] [CrossRef]
- Holman, D.B.; Kommadath, A.; Tingley, J.P.; Abbott, D.W. Novel Insights into the Pig Gut Microbiome Using Metagenome-Assembled Genomes. Microbiol. Spectr. 2022, 10, e0238022. [Google Scholar] [CrossRef] [PubMed]
- Wylensek, D.; Hitch, T.C.A.; Riedel, T.; Afrizal, A.; Kumar, N.; Wortmann, E.; Liu, T.; Devendran, S.; Lesker, T.R.; Hernández, S.B.; et al. A collection of bacterial isolates from the pig intestine reveals functional and taxonomic diversity. Nat. Commun. 2020, 11, 6389. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.J.; Wellington, M.; Shah, R.M.; Ferreira, M.J. Antibiotic Stewardship in Food-producing Animals: Challenges, Progress, and Opportunities. Clin. Ther. 2020, 42, 1649–1658. [Google Scholar] [CrossRef] [PubMed]
- Parajuli, P.; Rajput, M.I.; Verma, N.K. Plasmids of Shigella flexneri serotype 1c strain Y394 provide advantages to bacteria in the host. BMC Microbiol. 2019, 19, 86. [Google Scholar] [CrossRef] [PubMed]
- Ikkert, O.P.; Gerasimchuk, A.L.; Bukhtiyarova, P.A.; Tuovinen, O.H.; Karnachuk, O.V. Characterization of precipitates formed by H2S-producing, Cu-resistant Firmicute isolates of Tissierella from human gut and Desulfosporosinus from mine waste. Antonie Van Leeuwenhoek 2013, 103, 1221–1234. [Google Scholar] [CrossRef]
- Widdel, F.; Bak, F. Gram negative mesophilic sulfate reducing bacteria. In The Prokaryotes: A Handbook on the Biology of Bacteria: Ecophysiology, Isolation, Identification, Applications; Balows, A., Trüper, H.G., Dworkin, M., Harder, W., Schleifer, K.H., Eds.; Springer: Berlin/Heidelberg, Germany, 1992; pp. 3352–3378. [Google Scholar]
- Karnachuk, O.V.; Pimenov, N.V.; Iusupov, S.K.; Frank, I.A.; Puhakka, J.A.; Ivanov, M.V. Distribution, diversity, and activity of sulfate-reducing bacteria in the water column in Gek-Gel Lake, Azerbaijan. Mikrobiologiia 2006, 75, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Karnachuk, O.V.; Frank, Y.A.; Lukina, A.P.; Kadnikov, V.V.; Beletsky, A.V.; Mardanov, A.V.; Ravin, N.V. Domestication of previously uncultivated ‘Candidatus Desulforudis audaxviator’ from a deep aquifer in Siberia sheds light on its physiology and evolution. ISME J. 2019, 13, 1947–1959. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [Green Version]
- Waite, D.W.; Chuvochina, M.; Pelikan, C.; Parks, D.H.; Yilmaz, P.; Wagner, M.; Loy, A.; Naganuma, T.; Nakai, R.; Whitman, W.B.; et al. Proposal to reclassify the proteobacterial classes Deltaproteobacteria and Oligoflexia, and the phylum Thermodesulfobacteria into four phyla reflecting major functional capabilities. Int. J. Syst. Evol. Microbiol. 2020, 70, 5972–6016. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.B.; Stolyar, S.; Chivian, D.; Pinel, N.; Gabster, J.A.; Dehal, P.S.; He, Z.; Yang, Z.K.; Yen, H.C.; Zhou, J.; et al. Contribution of mobile genetic elements to Desulfovibrio vulgaris genome plasticity. Environ. Microbiol. 2009, 11, 2244–2252. [Google Scholar] [CrossRef] [Green Version]
- Konstantinidis, K.T.; Tiedje, J.M. Genomic insights that advance the species definition for prokaryotes. Proc. Natl. Acad. Sci. USA 2005, 102, 2567–2572. [Google Scholar] [CrossRef] [Green Version]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.J.; Park, Y.J.; Kim, J.S.; Lee, S.; Lee, S.H.; Choi, S.; Min, J.K.; Choi, I.; Ryu, C.M. Pseudomonas syringae evades phagocytosis by animal cells via type III effector-mediated regulation of actin filament plasticity. Environ. Microbiol. 2018, 20, 3980–3991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillings, M.; Boucher, Y.; Labbate, M.; Holmes, A.; Krishnan, S.; Holley, M.; Stokes, H.W. The evolution of class 1 integrons and the rise of antibiotic resistance. J. Bacteriol. 2008, 190, 5095–5100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, A.M.; Chartone-Souza, E. Operon mer: Bacterial resistance to mercury and potential for bioremediation of contaminated environments. Genet. Mol. Res. 2003, 2, 92–101. [Google Scholar]
- Nie, Y.; Xie, X.Q.; Zhou, L.; Guan, Q.; Ren, Y.; Mao, Y.; Shi, J.S.; Xu, Z.H.; Geng, Y. Desulfovibrio fairfieldensis-Derived Outer Membrane Vesicles Damage Epithelial Barrier and Induce Inflammation and Pyroptosis in Macrophages. Cells 2022, 12, 89. [Google Scholar] [CrossRef]
- Wang, S.; Liu, X.; Xu, X.; Yang, D.; Wang, D.; Han, X.; Shi, Y.; Tian, M.; Ding, C.; Peng, D.; et al. Escherichia coli Type III Secretion System 2 ATPase EivC Is Involved in the Motility and Virulence of Avian Pathogenic Escherichia coli. Front. Microbiol. 2016, 7, 1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portaliou, A.G.; Tsolis, K.C.; Loos, M.S.; Zorzini, V.; Economou, A. Type III Secretion: Building and Operating a Remarkable Nanomachine. Trends Biochem. Sci. 2016, 41, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Petnicki-Ocwieja, T.; Schneider, D.J.; Tam, V.C.; Chancey, S.T.; Shan, L.; Jamir, Y.; Schechter, L.M.; Janes, M.D.; Buell, C.R.; Tang, X.; et al. Genome wide identification of proteins secreted by the Hrp type III protein secretion system of Pseudomonas syringae pv. tomato DC3000. Proc. Natl. Acad. Sci. USA 2002, 99, 7652–7657. [Google Scholar] [CrossRef] [Green Version]
- Osborn, A.M.; Bruce, K.D.; Strike, P.; Ritchie, D.A. Distribution, diversity and evolution of the bacterial mercury resistance (mer) operon. FEMS Microbiol. Rev. 1997, 19, 239–262. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Meagher, R.J.; Light, Y.K.; Yilmaz, S.; Chakraborty, R.; Arkin, A.P.; Hazen, T.C.; Singh, A.K. Microfluidic fluorescence in situ hybridization and flow cytometry (μFlowFISH). Lab Chip 2011, 11, 2673–2679. [Google Scholar] [CrossRef]
- Feilberg, A.; Hansen, M.J.; Liu, D.; Nyord, T. Contribution of livestock H2S to total sulfur emissions in a region with intensive animal production. Nat. Commun. 2017, 8, 1069. [Google Scholar] [CrossRef] [Green Version]
- Lei, H.; Du, Q.; Lu, N.; Jiang, X.; Li, M.; Xia, D.; Long, K. Comparison of the microbiome-metabolome response to copper sulfate and copper glycinate in growing pigs. Animals 2023, 13, 345. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karnachuk, O.V.; Beletsky, A.V.; Rakitin, A.L.; Ikkert, O.P.; Avakyan, M.R.; Zyusman, V.S.; Napilov, A.; Mardanov, A.V.; Ravin, N.V. Antibiotic-Resistant Desulfovibrio Produces H2S from Supplements for Animal Farming. Microorganisms 2023, 11, 838. https://doi.org/10.3390/microorganisms11040838
Karnachuk OV, Beletsky AV, Rakitin AL, Ikkert OP, Avakyan MR, Zyusman VS, Napilov A, Mardanov AV, Ravin NV. Antibiotic-Resistant Desulfovibrio Produces H2S from Supplements for Animal Farming. Microorganisms. 2023; 11(4):838. https://doi.org/10.3390/microorganisms11040838
Chicago/Turabian StyleKarnachuk, Olga V., Alexey V. Beletsky, Andrey L. Rakitin, Olga P. Ikkert, Marat R. Avakyan, Viacheslav S. Zyusman, Andrei Napilov, Andrey V. Mardanov, and Nikolai V. Ravin. 2023. "Antibiotic-Resistant Desulfovibrio Produces H2S from Supplements for Animal Farming" Microorganisms 11, no. 4: 838. https://doi.org/10.3390/microorganisms11040838