
Biocontrol of Soft Rot Dickeya and Pectobacterium Pathogens by Broad-Spectrum Antagonistic Bacteria within Paenibacillus polymyxa Complex

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Media
2.2. In Vitro and In Vivo Screening of Antagonistic Paenibacillus Strains
2.3. In Vivo Biocontrol of D. dadantii by P. polymyxa ShX301
2.4. Antibacterial Activities of Cell-Free Culture Supernatant (CFCS) of P. polymyxa ShX301 against D. dadantii
2.5. Detection of Lipopeptides Produced by P. polymyxa ShX301
2.6. Statistical Analysis
3. Results
3.1. Strains within the P. polymyxa Complex Inhibited D. dadantii Growth and Maceration of Sweet Potato Tuber Slices
3.2. P. polymyxa ShX301 Showed Broad-Spectrum Antagonistic Activities against Dickeya and Pectobacterium
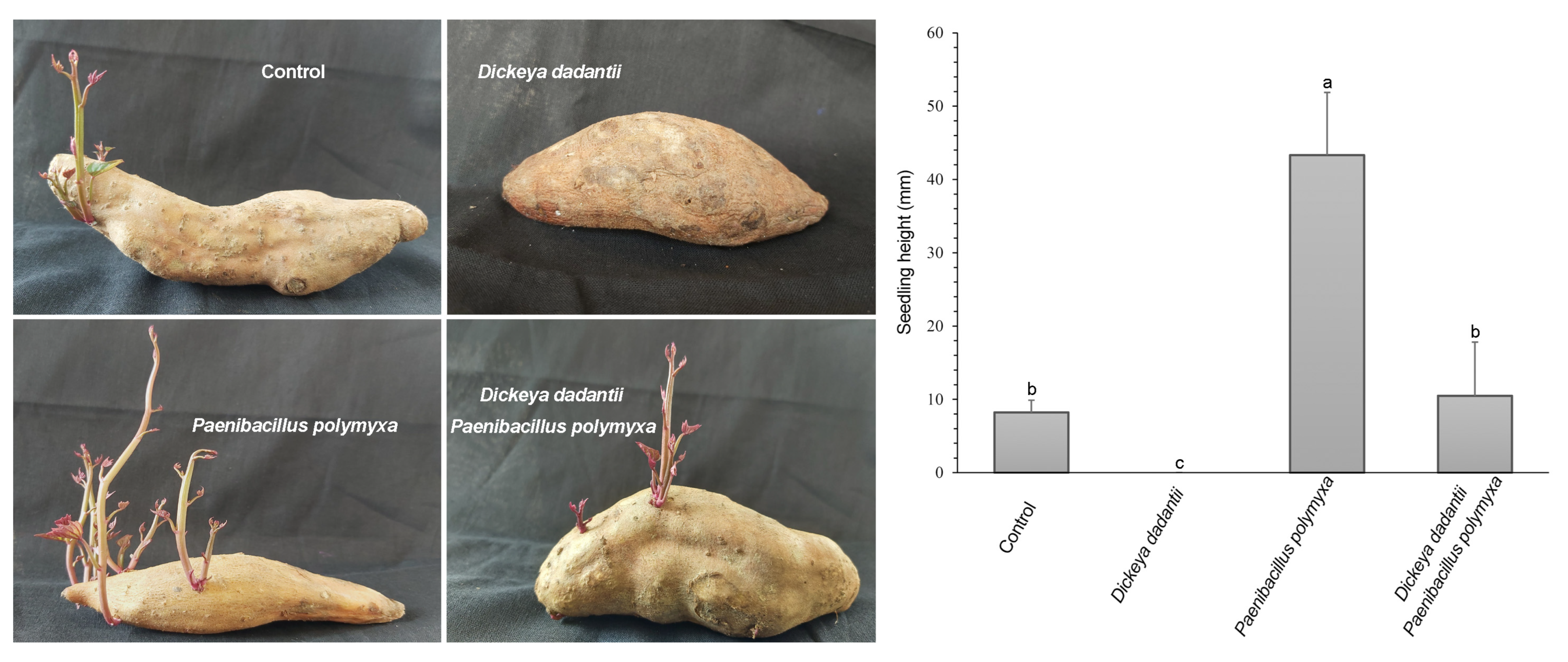
3.3. P. polymyxa ShX301 Protected Seed Tubers and Promoted Seedling Growth
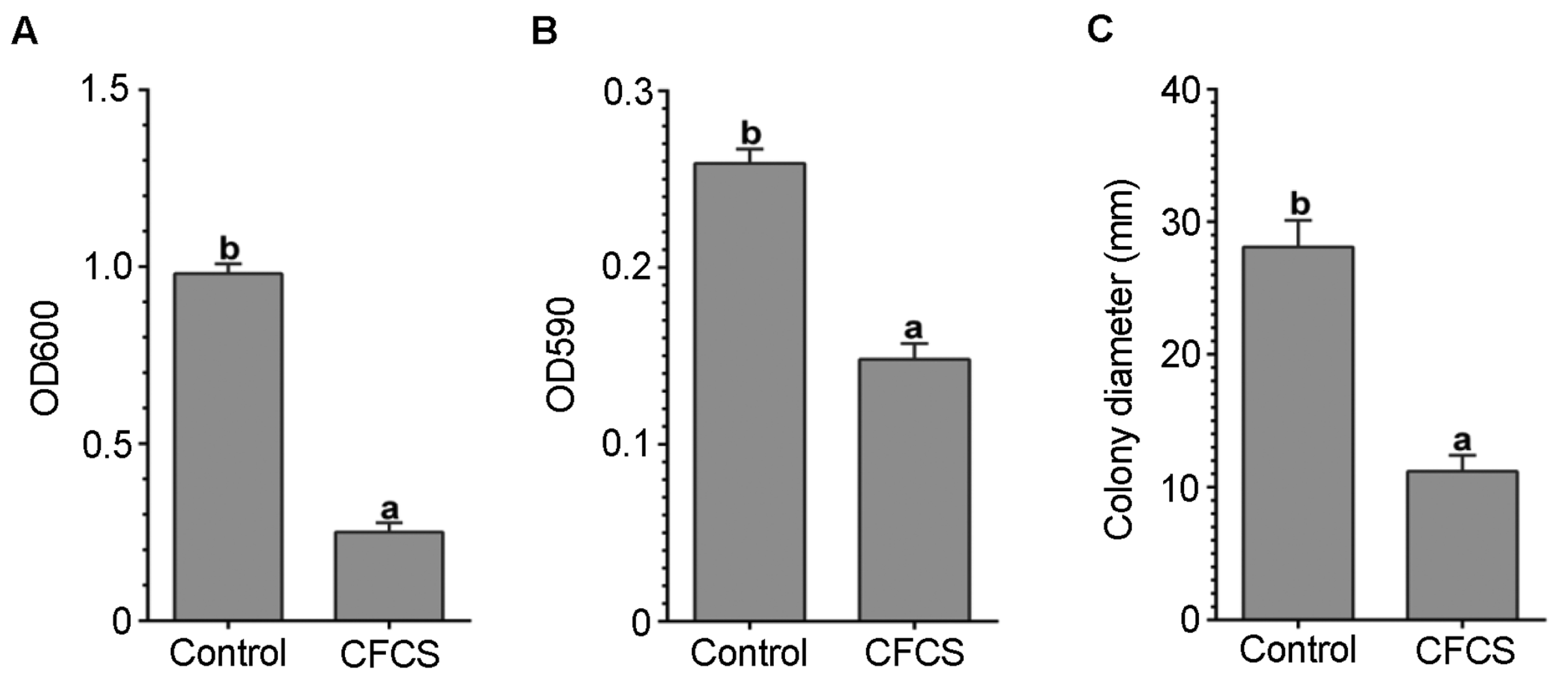
3.4. CFCS of P. polymyxa ShX301 Inhibited D. dadantii Growth, Biofilm Formation, and Swimming Motility
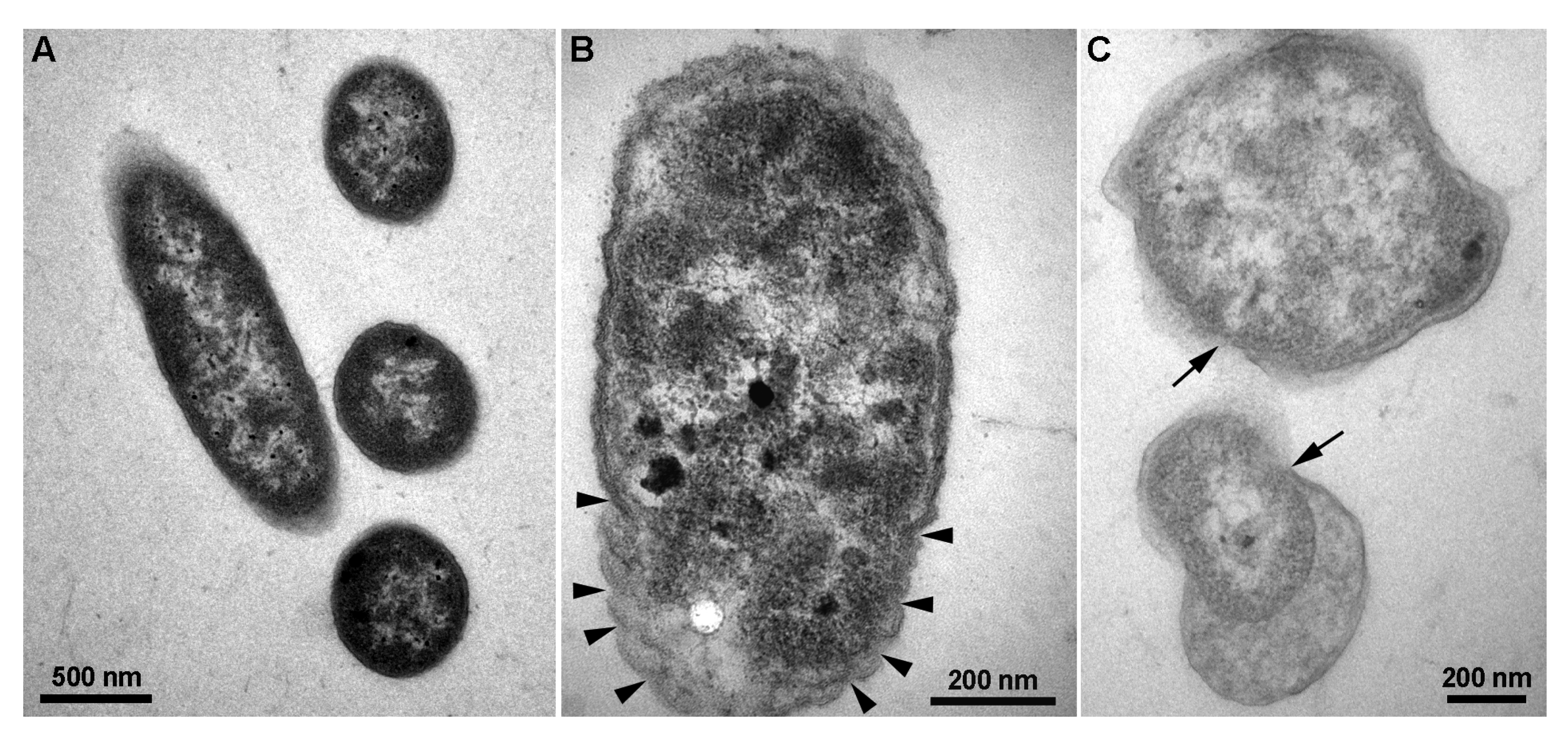
3.5. CFCS of P. polymyxa ShX301 Breached D. dadantii Cells
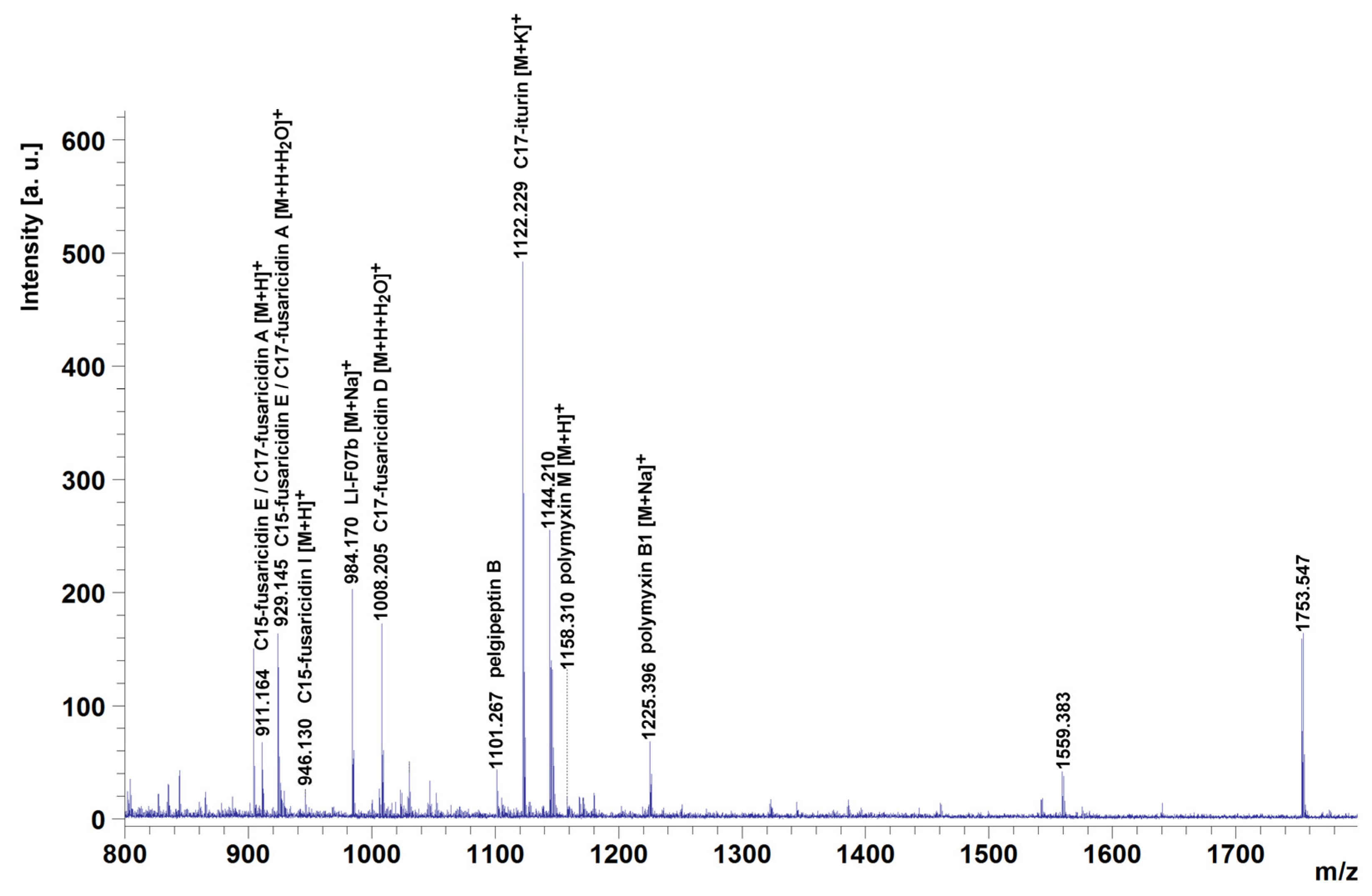
3.6. P. polymyxa ShX301 Produced Multiple Lipopeptides
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Charkowski, A.O. The changing face of bacterial soft rot diseases. Annu. Rev. Phytopathol. 2018, 56, 269–288. [Google Scholar] [CrossRef]
- Ma, B.; Hibbing, M.E.; Kim, H.S.; Reedy, R.M.; Yedidia, I.; Breuer, J.; Breuer, J.; Glasner, J.D.; Perna, N.T.; Kelman, A. Host range and molecular phylogenies of the soft rot enterobacterial genera Pectobacterium and Dickeya. Phytopathology 2007, 97, 1150–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degefu, Y.; Potrykus, M.; Golanowska, M.; Virtanen, E.; Lojkowska, E. A new clade of Dickeya spp. plays a major role in potato blackleg outbreaks in North Finland. Ann. Appl. Biol. 2013, 162, 231–241. [Google Scholar] [CrossRef]
- Van Gijsegem, F.; Toth, I.K.; van der Wolf, J.M. Outlook—Challenges and perspectives for management of diseases caused by Pectobacterium and Dickeya species. In Plant Diseases Caused by Dickeya and Pectobacterium Species; Springer: Cham, Switzerland, 2021; pp. 283–289. [Google Scholar]
- Liu, Q.G.; Zhang, Q.; Wei, C.D. Advances in research of rice bacterial foot rot. Sci. Agric. Sin. 2013, 46, 2923–2931. [Google Scholar]
- Toth, I.; Van Der Wolf, J.; Saddler, G.; Lojkowska, E.; Hélias, V.; Pirhonen, M.; Tsror, L.; Elphinstone, J. Dickeya species: An emerging problem for potato production in Europe. Plant Pathol. 2011, 60, 385–399. [Google Scholar] [CrossRef]
- Pu, X.; Zhou, J.; Lin, B.; Shen, H. First report of bacterial foot rot of rice caused by a Dickeya zeae in China. Plant Dis. 2012, 96, 1818. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Hunjan, M.S.; Kaur, H.; Rawal, R.; Kumar, A.; Singh, P. A review on bacterial stalk rot disease of maize caused by Dickeya zeae. J. Appl. Nat. Sci. 2017, 9, 1214–1225. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Fang, B.; Luo, Z.; Chen, J.; Zhang, X.; Wang, Z. First report of bacterial stem and root rot of sweet potato caused by a Dickeya sp. (Erwinia chrysanthemi) in China. Plant Dis. 2010, 94, 1503. [Google Scholar] [CrossRef]
- Stahr, M.N.; Butler, S.; Huerta, A.I.; Ritchie, D.F.; Quesada-Ocampo, L.M. First report of bacterial root rot, caused by Dickeya dadantii, on sweet potato (Ipomoea batatas) in North Carolina. Plant Dis. 2020, 104, 2723. [Google Scholar] [CrossRef]
- Liu, Q.; Xiao, W.; Wu, Z.; Li, S.; Yuan, Y.; Li, H. Identification of Dickeya dadantii as a causal agent of banana bacterial sheath rot in China. J. Plant Pathol. 2016, 98, 503–510. [Google Scholar]
- Zhang, J.; Shen, H.; Pu, X.; Lin, B.; Hu, J. Identification of Dickeya zeae as a causal agent of bacterial soft rot in banana in China. Plant Dis. 2014, 98, 436–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czajkowski, R. Bacteriophages of soft rot Enterobacteriaceae-a minireview. FEMS Microbiol. Lett. 2016, 363, fnv230. [Google Scholar] [CrossRef] [Green Version]
- Des Essarts, Y.R.; Cigna, J.; Quêtu-Laurent, A.; Caron, A.; Munier, E.; Beury-Cirou, A.; Hélias, V.; Faure, D. Biocontrol of the potato blackleg and soft rot diseases caused by Dickeya dianthicola. Appl. Environ. Microbiol. 2016, 82, 268–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, A.; Islam Masum, M.M.; Wu, X.; Abdallah, Y.; Ogunyemi, S.O.; Wang, Y.; Sun, G.; Li, B.; An, Q. Screening of Bacillus strains in biocontrol of pathogen Dickeya dadantii causing stem and root rot disease of sweet potato. Biocontrol Sci. Technol. 2020, 30, 1180–1198. [Google Scholar] [CrossRef]
- Grady, E.N.; MacDonald, J.; Liu, L.; Richman, A.; Yuan, Z.C. Current knowledge and perspectives of Paenibacillus: A review. Microb. Cell Factories 2016, 15, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rybakova, D.; Cernava, T.; Köberl, M.; Liebminger, S.; Etemadi, M.; Berg, G. Endophytes-assisted biocontrol: Novel insights in ecology and the mode of action of Paenibacillus. Plant Soil 2016, 405, 125–140. [Google Scholar] [CrossRef]
- Shafi, J.; Tian, H.; Ji, M. Bacillus species as versatile weapons for plant pathogens: A review. Biotechnol. Biotechnol. Equip. 2017, 31, 446–459. [Google Scholar] [CrossRef] [Green Version]
- Rabanal, F.; Cajal, Y. Recent advances and perspectives in the design and development of polymyxins. Nat. Prod. Rep. 2017, 34, 886–908. [Google Scholar] [CrossRef]
- Ali, M.A.; Lou, Y.; Hafeez, R.; Li, X.; Hossain, A.; Xie, T.; Lin, L.; Li, B.; Yin, Y.; Yan, J.; et al. Functional analysis and genome mining reveal high potential of biocontrol and plant growth promotion in nodule-inhabiting bacteria within Paenibacillus polymyxa complex. Front. Microbiol. 2021, 11, 618601. [Google Scholar] [CrossRef]
- Cochrane, S.A.; Vederas, J.C. Lipopeptides from Bacillus and Paenibacillus spp.: A gold mine of antibiotic candidates. Med. Res. Rev. 2016, 36, 4–31. [Google Scholar] [CrossRef]
- Needham, B.D.; Trent, M.S. Fortifying the barrier: The impact of lipid A remodelling on bacterial pathogenesis. Nat. Rev. Microbiol. 2013, 11, 467–481. [Google Scholar] [CrossRef]
- Perego, M.; Glaser, P.; Minutello, A.; Strauch, M.A.; Leopold, K.; Fischer, W. Incorporation of D-alanine into lipoteichoic acid and wall teichoic acid in Bacillus subtilis. Identification of genes and regulation. J. Biol. Chem. 1995, 270, 15598–15606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandin, C.; Caroff, M.; Condemine, G. Antimicrobial peptide resistance genes in the plant pathogen Dickeya dadantii. Appl. Environ. Microbiol. 2016, 82, 6423–6430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Li, X.L.; Zhu, S.J.; Ojaghian, M.R.; Zhang, J.Z. Biocontrol potential of Paenibacillus polymyxa against Verticillium dahliae infecting cotton plants. Biol. Control 2018, 127, 70–77. [Google Scholar] [CrossRef]
- Vater, J.; Herfort, S.; Doellinger, J.; Weydmann, M.; Dietel, K.; Faetke, S.; Lasch, P. Fusaricidins from Paenibacillus polymyxa M-1, a family of lipohexapeptides of unusual complexity—A mass spectrometric study. J. Mass Spectrom. 2017, 52, 7–15. [Google Scholar] [CrossRef]
- Vater, J.; Niu, B.; Dietel, K.; Borriss, R. Characterization of novel fusaricidins produced by Paenibacillus polymyxa M1 using MALDI-TOF mass spectrometry. J. Am. Soc. Mass Spectrom. 2015, 26, 1548–1558. [Google Scholar] [CrossRef] [Green Version]
- Huang, E.; Yang, X.; Zhang, L.; Moon, S.H.; Yousef, A.E. New Paenibacillus strain produces a family of linear and cyclic antimicrobial lipopeptides: Cyclization is not essential for their antimicrobial activity. FEMS Microbiol. Lett. 2017, 364, fnx049. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.C.; Shen, X.B.; Ding, R.; Qian, C.D.; Fang, H.H.; Li, O. Isolation and partial characterization of antibiotics produced by Paenibacillus elgii B69. FEMS Microbiol. Lett. 2010, 310, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Vater, J.; Gao, X.; Hitzeroth, G.; Wilde, C.; Franke, P. “Whole cell”-matrix-assisted laser desorption ionization-time of flight-mass spectrometry, an emerging technique for efficient screening of biocombinatorial libraries of natural compounds-present state of research. Comb. Chem. High Throughput Screen. 2003, 6, 557–567. [Google Scholar] [CrossRef]
- Bionda, N.; Pitteloud, J.P.; Cudic, P. Cyclic lipodepsipeptides: A new class of antibacterial agents in the battle against resistant bacteria. Future Med. Chem. 2013, 5, 1311–1330. [Google Scholar] [CrossRef] [Green Version]
- Niu, B.; Vater, J.; Rueckert, C.; Blom, J.; Lehmann, M.; Ru, J.J.; Chen, X.H.; Wang, Q.; Borriss, R. Polymyxin P is the active principle in suppressing phytopathogenic Erwinia spp. by the biocontrol rhizobacterium Paenibacillus polymyxa M-1. BMC Microbiol. 2013, 13, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Le, K.D.; Yu, N.H.; Kim, J.I.; Kim, J.C.; Lee, C.W. Structure and antifungal activity of pelgipeptins from Paenibacillus elgii against phytopathogenic fungi. Pestic. Biochem. Physiol. 2020, 163, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Le, K.D.; Kim, J.; Yu, N.H.; Kim, B.; Lee, C.W.; Kim, J.C. Biological control of tomato bacterial wilt, kimchi cabbage soft rot, and red pepper bacterial leaf spot using Paenibacillus elgii JCK-5075. Front. Plant Sci. 2020, 11, 775. [Google Scholar] [CrossRef] [PubMed]
- Zeriouh, H.; Romero, D.; García-Gutiérrez, L.; Cazorla, F.M.; de Vicente, A.; Pérez-García, A. The iturin-like lipopeptides are essential components in the biological control arsenal of Bacillus subtilis against bacterial diseases of cucurbits. Mol. Plant Microbe Interact. 2011, 24, 1540–1552. [Google Scholar] [CrossRef] [Green Version]
- Antúnez-Lamas, M.; Cabrera-Ordonez, E.; Lopez-Solanilla, E.; Raposo, R.; Trelles-Salazar, O.; Rodríguez-Moreno, A.; Rodríguez-Palenzuela, P. Role of motility and chemotaxis in the pathogenesis of Dickeya dadantii 3937 (ex Erwinia chrysanthemi 3937). Microbiology 2009, 155, 434–442. [Google Scholar] [CrossRef] [Green Version]
- Jahn, C.E.; Selimi, D.A.; Barak, J.D.; Charkowski, A.O. The Dickeya dadantii biofilm matrix consists of cellulose nanofibres, and is an emergent property dependent upon the type III secretion system and the cellulose synthesis operon. Microbiology 2011, 157, 2733–2744. [Google Scholar] [CrossRef] [Green Version]
- Joko, T.; Hirata, H.; Tsuyumu, S. Sugar transporter (MfsX) of the major facilitator superfamily is required for flagella-mediated pathogenesis in Dickeya dadantii 3937. J. Gen. Plant Pathol. 2007, 73, 266–273. [Google Scholar] [CrossRef]
- Prigent-Combaret, C.; Zghidi-Abouzid, O.; Effantin, G.; Lejeune, P.; Reverchon, S.; Nasser, W. The nucleoid-associated protein Fis directly modulates the synthesis of cellulose, an essential component of pellicle–biofilms in the phytopathogenic bacterium Dickeya dadantii. Mol. Microbiol. 2012, 86, 172–186. [Google Scholar] [CrossRef] [PubMed]
- Quinn, G.A.; Maloy, A.P.; McClean, S.; Carney, B.; Slater, J.W. Lipopeptide biosurfactants from Paenibacillus polymyxa inhibit single and mixed species biofilms. Biofouling 2012, 28, 1151–1166. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, Y.; Ge, K.; Du, B.; Liu, K.; Wang, C.; Ding, Y. Interactional mechanisms of Paenibacillus polymyxa SC2 and pepper (Capsicum annuum L.) suggested by transcriptomics. BMC Microbiol. 2021, 21, 70. [Google Scholar] [CrossRef]
- Li, Y.; Chen, S. Fusaricidin produced by Paenibacillus polymyxa WLY78 induces systemic resistance against Fusarium wilt of cucumber. Int. J. Mol. Sci. 2019, 20, 5240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmusk, S.; Wagner, E.G.H. The plant-growth-promoting rhizobacterium Paenibacillus polymyxa induces changes in Arabidopsis thaliana gene expression: A possible connection between biotic and abiotic stress responses. Mol. Plant Microbe Interact. 1999, 12, 951–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsror, L.; Erlich, O.; Hazanovsky, M.; Ben Daniel, B.; Zig, U.; Lebiush, S. Detection of Dickeya spp. latent infection in potato seed tubers using PCR or ELISA and correlation with disease incidence in commercial field crops under hot-climate conditions. Plant Pathol. 2012, 61, 161–168. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Isolation Source |
|---|---|
| Paenibacillus polymyxa ShX301 | Cotton rhizosphere |
| P. polymyxa ShX302 | Cotton rhizosphere |
| P. polymyxa ShX303 | Cotton rhizosphere |
| P. polymyxa RP31 | Black locust nodule |
| P. peoriae RP20 | Black locust nodule |
| P. peoriae RP51 | Black locust nodule |
| P. peoriae RP62 | Black locust nodule |
| P. peoriae CFCC 1854 | Dendrolobium triangulare nodule |
| P. kribbensis CFCC 1865 | Ormosia semicastrata nodule |
| Dickeya dadantii CZ1501 | Sweet potato |
| D. dadantii NCPPB 898T | Pelargonium capitatum |
| D. solani NCPPB 4479 | Potato |
| D. dianthicola NCPPB 3534 | Potato |
| D. fangzhongdai CGMCC 1.15464T | Pyrus pyrifolia |
| D. chrysanthemi NCPPB 402T | Chrysanthemum |
| D. oryzae ACCC 61554T | Rice |
| Pectobacterium carotovorum CFCC 10814T | Potato |
| Pectobacterium cacticida CFCC 10813T | Cactus |
| Paenibacillus Strains | In Vitro Inhibition Rate (%) | In Vivo Inhibition Rate (%) |
|---|---|---|
| P. polymyxa ShX301 | 53 ± 4 a,* | 80 ± 2 a,* |
| P. polymyxa ShX302 | 39 ± 3 bc | 57 ± 1 c |
| P. polymyxa ShX303 | 44 ± 3 ab | 70 ± 1 b |
| P. polymyxa RP 31 | 42 ± 1 ab | 67 ± 2 b |
| P. peoriae RP 51 | 37 ± 5 bc | 57 ± 2 c |
| P. peoriae RP 20 | 16 ± 3 e | 33 ± 2 f |
| P. peoriae RP 62 | 13 ± 4 e | 20 ± 1 g |
| P. peoriae CFCC 1854 | 22 ± 3 de | 38 ± 2 e |
| P. kribbensis CFCC 1865 | 30 ± 2 cd | 48 ±2 d |
| Dickeya and Pectobacterium | Inhibition Rate (%) | |
|---|---|---|
| Nutrient Agar | M9 Agar | |
| D. dadantii CZ1501 | 53 ± 3 a,* | 74 ± 2 b,* |
| D. dadantii NCPPB 898T | 50 ± 2 a | 70 ± 3 b |
| D. solani NCPPB 4479 | 45 ± 2 a | 65 ± 2 b |
| D. dianthicola NCPPB 3530 | 54 ± 3 a | 72 ± 2 b |
| D. fangzhongdai CGMCC 1.15464T | 48 ± 1 a | 69 ± 2 b |
| D. chrysanthemi NCPPB 402T | 53 ± 1 a | 72 ± 1 b |
| D. zeae NCPPB 3531 | 46 ± 3 a | 64 ± 1 b |
| P. carotovorum CFCC 10814T | 54 ± 2 a | 72 ± 3 b |
| P. cacticida CFCC 10813T | 52 ± 2 a | 70 ± 2 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, A.; Ali, M.A.; Lin, L.; Luo, J.; You, Y.; Masum, M.M.I.; Jiang, Y.; Wang, Y.; Li, B.; An, Q. Biocontrol of Soft Rot Dickeya and Pectobacterium Pathogens by Broad-Spectrum Antagonistic Bacteria within Paenibacillus polymyxa Complex. Microorganisms 2023, 11, 817. https://doi.org/10.3390/microorganisms11040817
Hossain A, Ali MA, Lin L, Luo J, You Y, Masum MMI, Jiang Y, Wang Y, Li B, An Q. Biocontrol of Soft Rot Dickeya and Pectobacterium Pathogens by Broad-Spectrum Antagonistic Bacteria within Paenibacillus polymyxa Complex. Microorganisms. 2023; 11(4):817. https://doi.org/10.3390/microorganisms11040817
Chicago/Turabian StyleHossain, Afsana, Md. Arshad Ali, Li Lin, Jinyan Luo, Yuxin You, Md. Mahidul Islam Masum, Yugen Jiang, Yanli Wang, Bin Li, and Qianli An. 2023. "Biocontrol of Soft Rot Dickeya and Pectobacterium Pathogens by Broad-Spectrum Antagonistic Bacteria within Paenibacillus polymyxa Complex" Microorganisms 11, no. 4: 817. https://doi.org/10.3390/microorganisms11040817