

Screening for Multifarious Plant Growth Promoting and Biocontrol Attributes in Bacillus Strains Isolated from Indo Gangetic Soil for Enhancing Growth of Rice Crops
 , , , ,
, , , , 
Abstract
:1. Introduction
2. Material and Methods
2.1. Collection of Soil Samples from the Paddy Rhizosphere
Isolation and Maintenance of Bacterial Isolates
2.2. Characterization of Soil Bacterial Isolates for Various PGP Attributes
2.2.1. Phosphate Solubilization
2.2.2. IAA Production and Siderophore Production
2.2.3. HCN Production
2.2.4. Ammonia Production
2.2.5. ACC Deaminase Activity
2.3. In Vitro Screening of the Bacterial Isolates for their Antagonistic Activities against Soil Borne Plant Pathogens
2.3.1. Fungal Culture
2.3.2. Qualitative Evaluation of Antagonism Due to Diffusible Compounds
2.4. Production of Cell Wall Hydrolytic and Industrially Important Enzymes by Potential Bacillus Strains
2.4.1. Chitinase and Protease Production
2.4.2. Pectinase Activity
2.4.3. Cellulase, Lipase and Amylase Activity
2.5. Morphological and Biochemical Characterization
2.6. Molecular Characterization of the Potential Isolates
2.7. Abiotic Stress Tolerance Activity of the Potential Bacillus Strains
2.7.1. Drought Resistance
2.7.2. Temperature Resistance
2.7.3. Salt Tolerance
2.8. In Vivo Experiment to Evaluate the Efficacy of Potential Isolates to Promot Plant Growth under Green House Conditions
2.8.1. Pot Culture Assay under Greenhouse Conditions
2.8.2. Seed Bacterization
2.9. Statistical Analysis
3. Results and Discussion
3.1. Plant Growth Promoting Activity of Bacillus Isolates Recovered from Soils of the IGP Region
3.1.1. IAA Production
3.1.2. Phosphate Solubilization
3.1.3. Siderophore Production
3.2. In Vitro Antifungal Activity of the Potential Bacillus Isolates
3.2.1. HCN Production
3.2.2. Ammonia Production
3.2.3. ACC Deaminase Activity
3.3. Screening of Industrially Important and Cell Wall Hydrolytic Enzymes by Potential Bacillus Isolates
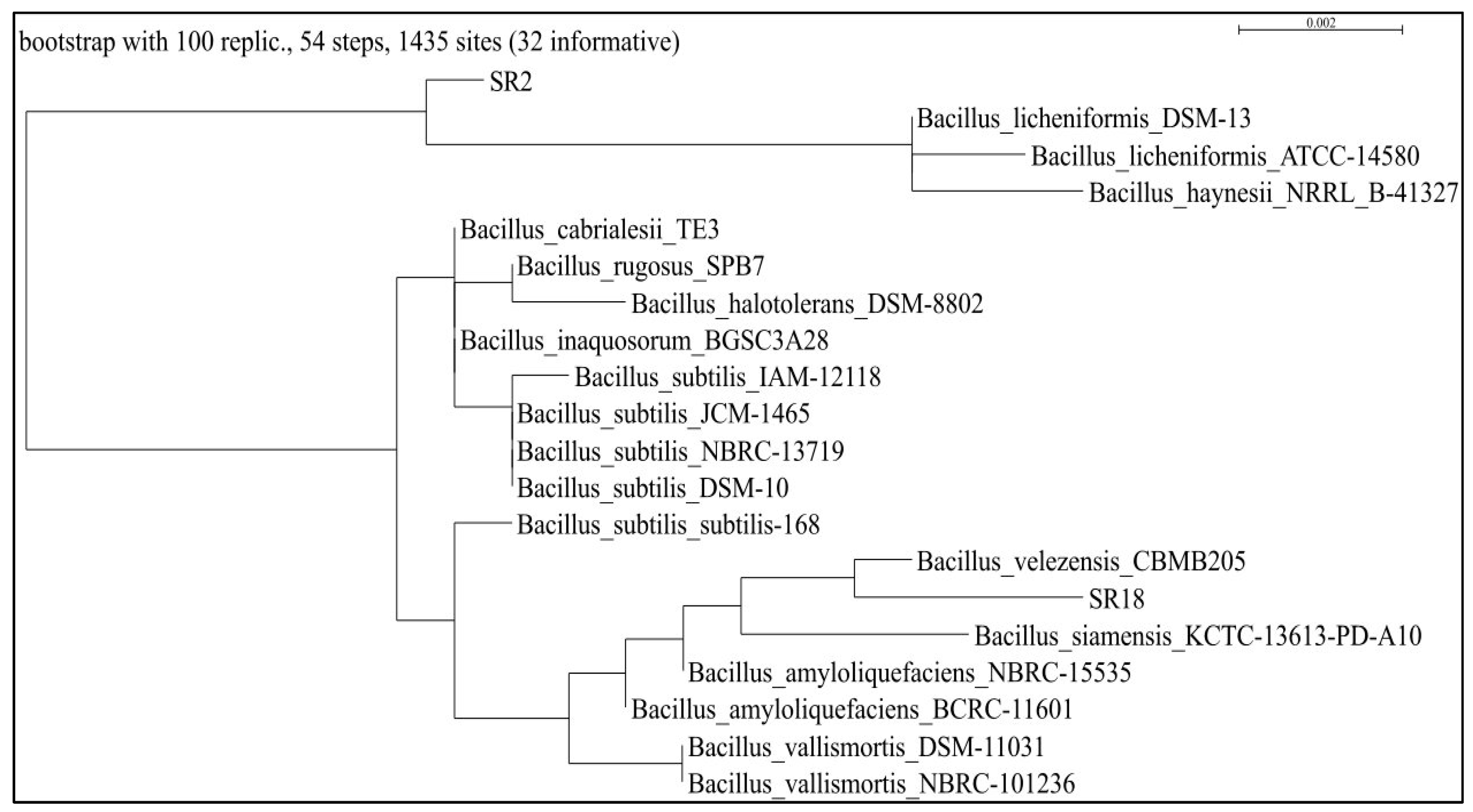
3.4. Molecular Characterization of the Potential Isolates
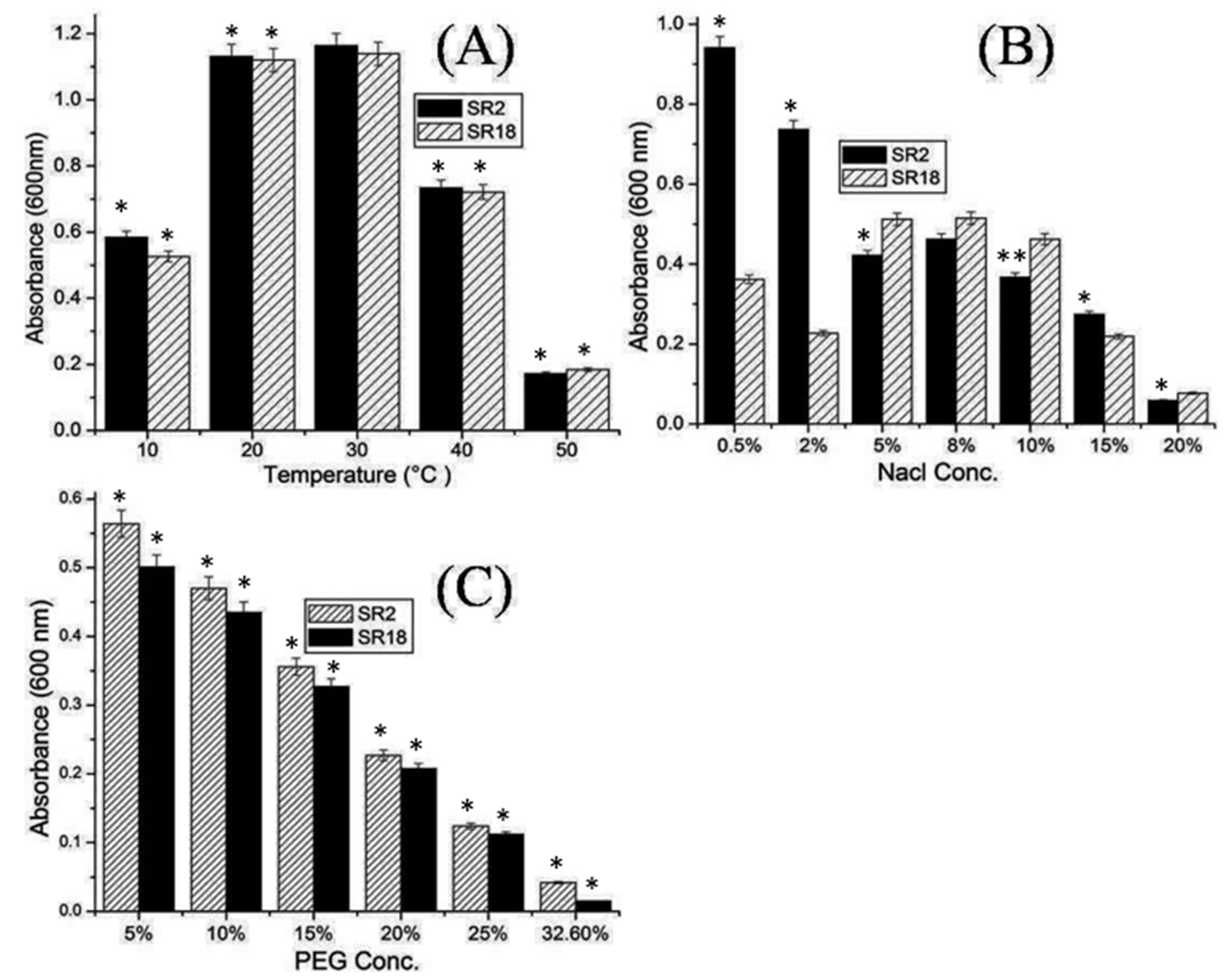
3.5. Abiotic Stress Tolerance Activity of Potential Bacillus Strains Recovered from Soils of IGP Region
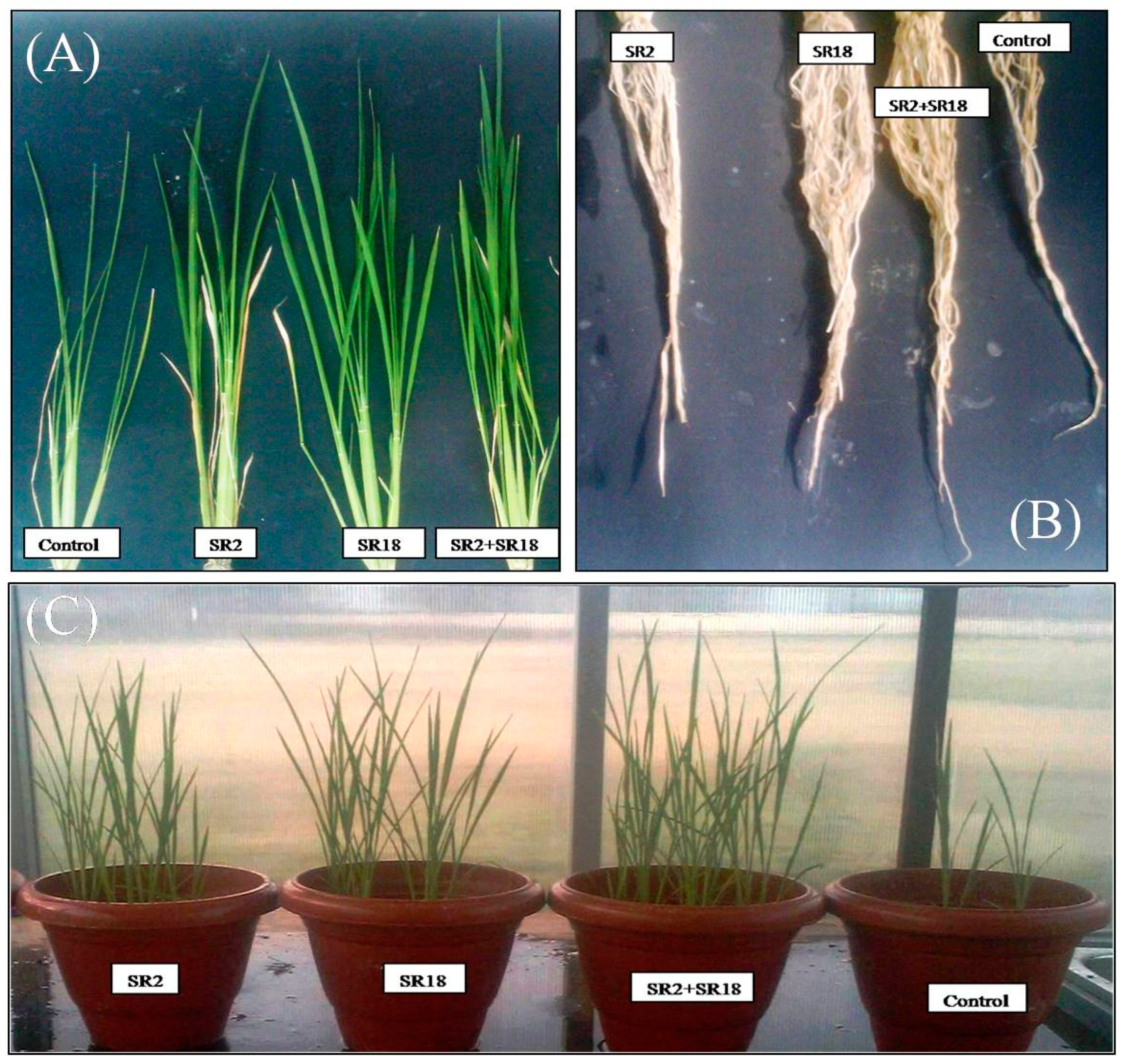
3.6. Effect of Rice Associated Bacilli on Growth of Rice Plants Growing under Green House Conditions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Goswami, D.; Dhandhukia, P.; Patela, P.; Thakkera, J.N. Screening of PGPR from saline desert of Kutch: Growth promotion in Arachis hypogea by Bacillus licheniformis A2. Microbiol. Res. 2014, 169, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Aeron, A.; Khare, E.; Jha, C.K.; Meena, V.S.; Aziz, S.M.; Islam, M.T.; Kim, K.; Meena, S.K.; Pattanayak, A.; Rajashekara, H.; et al. Revisiting the plant growth-promoting rhizobacteria: Lessons from the past and objectives for the future. Arch. Microbiol. 2020, 202, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.P.; Ahmed, S.K.M.H.; Desai, S.; Amalraj, E.L.D.; Rasul, A. In Vitro screening for abiotic stress tolerance in potent biocontrol and plant growth promoting strains of Pseudomonas and Bacillus spp. Int. J. Microbiol. 2014, 2014, 195946. [Google Scholar]
- Wang, H.; Liu, R.; You, M.P.; Barbetti, M.J.; Chen, Y. Pathogen biocontrol using plant growth-promoting bacteria (PGPR): Role of bacterial diversity. Microorganisms 2021, 9, 1988. [Google Scholar] [CrossRef]
- Jiao, X.; Takishita, Y.; Zhou, G.; Smith, D.L. Plant associated rhizobacteria for biocontrol and plant growth enhancement. Front. Plant Sci. 2021, 12, 634796. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Saad, A.M.; Soliman, S.M.; Salem, H.M.; Ahmed, A.I.; Mahmood, M.; El-Tahan, A.M.; Ebrahim, A.A.M.; Abd El-Mageed, T.A.; Negm, S.H. Plant growth-promoting microorganisms as biocontrol agents of plant diseases: Mechanisms, challenges and future perspectives. Front. Plant Sci. 2022, 13, 923880. [Google Scholar] [CrossRef]
- Pahalvi, H.N.; Rafiya, L.; Rashid, S.; Nisar, B.; Kamili, A.N. Chemical fertilizers and their impact on soil health. microbiota and biofertilizers. Ecofriendly Tools Reclam. Degrad. Soil Env. 2021, 2, 1–20. [Google Scholar]
- Khoshru, B.; Sarikhani, M.R.; Reyhanitabar, A.; Oustan, S.; Malboobi, M.A. Evaluation of the ability of rhizobacterial isolates to solubilize sparingly soluble iron under in vitro conditions. Geomicrobiol. J. 2022, 39, 804–815. [Google Scholar] [CrossRef]
- García-Salamanca, A.; Molina-Henares, M.A.; van Dillewijn, P.; Solano, J.; Pizarro-Tobías, P.; Roca, A. Bacterial diversity in the rhizosphere of maize and the surrounding carbonate-rich bulk soil. Microb. Biotechnol. 2013, 6, 36–44. [Google Scholar] [CrossRef]
- Majeed, A.; Muhammad, Z.; Ahmad, H. Plant growth promoting bacteria: Role in soil improvement, abiotic and biotic stress management of crops. Plant Cell Rep. 2018, 37, 1599–1609. [Google Scholar] [CrossRef]
- Johnson, L.F.; Curl, E.A. Isolation of Groups of Microorganisms from Soil. Methods for Research in Ecology of Soil-Borne Plant Pathogens; Burgess Publishing Co.: New York, NY, USA, 1972; p. 142. [Google Scholar]
- Pereira, S.I.A.; Abreu, D.; Moreira, H.; Vega, A.; Castro, P.M.L. Plant growth-promoting rhizobacteria (PGPR) improve the growth and nutrient use efficiency in maize (Zea mays L.) under water deficit conditions. Heliyon 2020, 6, e05106. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.H.; Gururani, M.A.; Chun, S. Isolation and characterization of plant growth promoting endophytic diazotrophic bacteria from Korean rice cultivars. Microbiol. Res. 2014, 169, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Slimani, A.; Raklami, A.; Oufdou, K.; Meddich, A. Isolation and characterization of PGPR and their potential for drought alleviation in barley Plants. Gesunde. Pflanzen. 2022, 27, 1–5. [Google Scholar]
- Sehrawat, A.; Sindhu, S.S.; Glick, B.R. Hydrogen cyanide production by soil bacteria: Biological control of pests and promotion of plant growth in sustainable agriculture. Pedosphere 2022, 32, 15–38. [Google Scholar] [CrossRef]
- Cappuccino, J.G.; Sherman, N. Biochemical activities of microorganisms. In Microbiology, A Laboratory Manual, 3rd ed.; The Benjamin/Cummings Publishing Co.: San Francisco, CA, USA, 1992; pp. 125–179. [Google Scholar]
- Dworkin, M.; Foster, J. Experiments with some microorganisms which utilize ethane and hydrogen. J. Bacteriol. 1958, 75, 592–601. [Google Scholar] [CrossRef]
- Bhattacharyya, C.; Banerjee, S.; Acharya, U.; Mitra, A.; Mallick, I.; Haldar, A.; Haldar, S.; Ghosh, A.; Ghosh, A. Evaluation of plant growth promotion properties and induction of antioxidative defense mechanism by tea rhizobacteria of Darjeeling, India. Sci. Rep. 2020, 10, 15536. [Google Scholar] [CrossRef]
- Ganesan, P.; Gnanamanickam, S.S. Biological control of Sclerotium rolfsii Sacc. in peanut by inoculation with Pseudomonas fluorescens. Soil Biol. Biochem. 1987, 19, 35–38. [Google Scholar] [CrossRef]
- Naik, P.; Raman, G.; Narayanan, K.; Sakthivel, N. Assessment of genetic and functional diversity of phosphate solubilizing fluorescent pseudomonads isolated from rhizospheric soil. BMC Microbiol. 2008, 8, 230. [Google Scholar] [CrossRef]
- Cattelan, A.P.; Hartel Furhmann, F. Screening for plant growth promoting rhizobacteria to promote early soybean growth. Soil Sci. Soc. Am. J. 1999, 63, 1670–1680. [Google Scholar] [CrossRef]
- Weisberg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F. The Clustal X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic. Acids. Res. 1997, 24, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Sandhya, V.; Ali, S.Z.; Venkatesvarlu, B.; Reddy, G.; Grover, M. Effect of osmotic stress on plant growth promoting Pseudomonas spp. Arch. Microbiol. 2020, 192, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Paungfoo-Lonhienne, C.; Redding, M.; Pratt, C.; Wang, W. Plant growth promoting rhizobacteria increase the efficiency of fertilisers while reducing nitrogen loss. J. Environ. Management. 2019, 233, 337–341. [Google Scholar] [CrossRef]
- Weller, D.M.; Cook, R.J. Suppression of take all the wheat by seed treatment with fluorescent Pseudomonads. Phytopathol 1983, 23, 23–54. [Google Scholar] [CrossRef]
- Shaffique, S.; Khan, M.A.; Wani, S.H.; Pande, A.; Imran, M.; Kang, S.M.; Rahim, W.; Khan, S.A.; Bhatta, D.; Kwon, E.H.; et al. A review on the role of endophytes and plant growth promoting rhizobacteria in mitigating heat stress in plants. Microorganisms 2022, 10, 1286. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Faraz, A.; Faizan, M. Root exudates: Composition and impact on plant–microbe interaction. Biofilms Plant Soil Health 2017, 179–193. [Google Scholar] [CrossRef]
- Walker, V.; Bertrand, C.; Bellvert, F.; Moënne-Loccoz, Y.; Bally, R.; Comte, G. Host plant secondary metabolite profiling shows complex, strain-dependent response of maize to the plant growth promoting rhizobacteria of the genus Azospirillum. New Phytol. 2011, 189, 494–506. [Google Scholar] [CrossRef]
- Mahdi, I.; Allaoui, A.; Fahsi, N.; Biskri, L. Bacillus velezensis QA2 potentially induced salt stress tolerance and enhanced phosphate uptake in quinoa plants. Microorganisms 2022, 10, 1836. [Google Scholar] [CrossRef]
- Rodríguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of phosphate-solubilizing microorganisms in sustainable agriculture—A review. Agron. Sustain. Dev. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophore. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, D.; O’Gara, F. Traits of fluorescent Pseudomonas spp. involved in suppression of plant pathogens. Microbiol. Rev. 1992, 56, 662–676. [Google Scholar] [CrossRef]
- Ferreira, M.J.; Silva, H.; Cunha, A. Siderophore-producing rhizobacteria as a promising tool for empowering plants to cope with iron limitation in saline soils: A review. Pedosphere 2019, 29, 409–420. [Google Scholar] [CrossRef]
- Lankford, C.E. Bacterial assimilation of iron. Cr. Rev. Microbiol. 1973, 2, 273–331. [Google Scholar] [CrossRef]
- Miljaković, D.; Marinković, J.; Balešević-Tubić, S. The significance of Bacillus spp. in disease suppression and growth promotion of field and vegetable crops. Microorganisms 2020, 8, 1037. [Google Scholar] [CrossRef]
- Ahmad, A.G.M.; Attia, A.Z.G.; Mohamed, M.S.; Elsayed, H.E. Fermentation, formulation and evaluation of PGPR Bacillus subtilis isolate as a bioagent for reducing occurrence of peanut soil-borne diseases. J. Integr. Agric. 2019, 18, 2080–2092. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, A.; Devi, S.; Patil, S.; Payal, C.; Negi, S. Isolation, screening and characterization of bacteria from rhizospheric soils for different plant growth promotion (PGP) activities: An in vitro study. Recent. Res. Sci. Technol. 2012, 4, 01–05. [Google Scholar]
- Bal, H.B.; Das, S.; Dangar, T.K.; Adhya, T.K. ACC deaminase and IAA producing growth promoting bacteria from the rhizosphere soil of tropical rice plants. J. Basic Microbiol. 2013, 53, 972–984. [Google Scholar] [CrossRef]
- Vanderstraeten, L.; Depaepe, T.; Bertrand, S.; Van Der Straeten, D. The ethylene precursor ACC affects early vegetative development independently of ethylene signaling. Front. Plant Sci. 2019, 10, 1591. [Google Scholar] [CrossRef]
- Jabborova, D.; Kannepalli, A.; Davranov, K.; Narimanov, A.; Enakiev, Y.; Syed, A.; Elgorban, A.M.; Bahkali, A.H.; Wirth, S.; Sayyed, R.Z.; et al. Co-inoculation of rhizobacteria promotes growth, yield, and nutrient contents in soybean and improves soil enzymes and nutrients under drought conditions. Sci. Rep. 2021, 11, 22081. [Google Scholar] [CrossRef] [PubMed]
- Kapri, A.; Tewari, L. Phosphate solubilization potential and phosphatase activity of rhizospheric Trichoderma spp. Brazilian J. Microbiol. 2010, 41, 787–795. [Google Scholar] [CrossRef]
- Chabot, R.; Antoun, H.; Cescas, P.M. Growth promotion of maize and lettuce by phosphate solubilizing Rhizobium leguminosarum biovar. Plant Soil. 1996, 184, 311–321. [Google Scholar] [CrossRef]
- Cardinale, M.; Ratering, S.; Suarez, C.; Montoya, A.M.Z.; Geissler-Plaum, R.; Schnell, S. Paradox of plant growth promotion potential of rhizobacteria and their actual promotion effect on growth of barley (Hordeum vulgare L.) under salt stress. Microbiol. Res. 2015, 181, 22–32. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Isolate | IAA (µg/mL) | PSI (mm) | Siderophore a | Biocontrol | HCN b | Ammonia b | ACC c | Cellulase | Protease | Lipase | Amylase | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R. Solani | F. Oxysporum | |||||||||||
| SR2 | 90.64 | 11 | ++ | 62% | 67% | - | + | ++ | Positive | Positive | Positive | Positive |
| SR3 | 56.32 | 2 | ++ | - | 29% | + | + | + | Positive | - | - | Positive |
| SR11 | 67.56 | 5 | ++ | - | - | - | - | - | - | - | - | - |
| SR13 | 23.67 | 2 | + | 32% | 38% | - | + | - | - | Positive | Positive | Positive |
| SR18 | 87.15 | 8.5 | ++++ | 50% | 48% | ++ | + | ++ | - | Positive | Positive | Positive |
| SR20 | 40.30 | 5 | + | - | - | - | + | + | - | - | Positive | - |
| SR56 | 17.12 | 2 | ++ | 31% | 35% | - | - | + | Positive | - | - | Positive |
| SR58 | 25.43 | - | + | - | - | - | + | - | - | positive | - | - |
| S.No. | Property | Bacillus Strains | |
|---|---|---|---|
| SR2 | SR18 | ||
| 1 | Gram nature | +, rods | +, rods |
| 2 | Endospore | Present | Present |
| 3 | Colony morphology | Colonies are irregular form, lobate margin, with bulging droplets of mucilaginous growth. | Colonies are larger, undulate margin, with circular form and flat elevation. |
| 4 | Motility | +ve | +ve |
| 5 | Oxidase | +ve | −ve |
| 6 | Catalase | +ve | +ve |
| 7 | Indole | −ve | −ve |
| 8 | Methyl red | +ve | −ve |
| 9 | Voges-Proskauer | +ve | +ve |
| 10 | Citrate | +ve | −ve |
| 11 | Starch hydrolysis | +ve | +ve |
| 12 | Gelatin hydrolysis | +ve | +ve |
| 13 | Urease | −ve | −ve |
| 14 | Nitrate | +ve | +ve |
| S.No. | Bacillus Strains | Closest Sequence | % Similarity | e-Value | Genebank Accession No. |
|---|---|---|---|---|---|
| 1. | SR2 | Bacillus licheniformis | 99% | 0.00 | KM052376 |
| 2. | SR18 | Bacillus velezensis | 99.51% | 0.00 | KM052377 |
| Treatment | Growth Parameters | |||||||
|---|---|---|---|---|---|---|---|---|
| Shoot Length (cm) | Shoot Dry Weight (g/plant) | Tiller Number | Leaf Number | Root Length (cm) | Root Number | Root Dry Weight (g/plant) | Yield (g/plant) | |
| Control (Untreated) | 45.0 ± 1.0 a | 0.288 ± 0.02 a | 3.3 ± 0.5 a | 12 ± 1.0 a | 17.8 ± 1.15 a | 51.6± 7.6 a | 0.170 ± 0.02 a | 24.40 ± 2.10 a |
| A (SR2) | 51.6 ± 0.5 b | 0.441 ± 0.01 ab | 4.6 ± 0.5 ab | 16.6 ± 2.08 b | 23.3 ± 0.76 bc | 69.6 ± 8.5 b | 0.231 ± 0.02 ab | 26.33 ± 0.76 a |
| B (SR18) | 52.7 ± 2.5 b | 0.543 ± 0.00 b | 7.0 ± 1.0 ab | 21.3 ± 1.52 c | 21.4 ± 0.55 ab | 161.3 ± 3.5 c | 0.313 ± 0.00 b | 27.53 ± 2.17 ab |
| A + B (SR2 + SR18) | 57.2 ± 1.16 c | 0.548 ± 0.01 b | 8.3 ± 0.5 b | 25.3 ± 2.08 d | 27.1 ± 1.86 c | 158.3 ± 7.6 c | 0.326 ± 0.15 b | 29.17 ± 3.27 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Devi, S.; Sharma, S.; Tiwari, A.; Bhatt, A.K.; Singh, N.K.; Singh, M.; Kaushalendra; Kumar, A. Screening for Multifarious Plant Growth Promoting and Biocontrol Attributes in Bacillus Strains Isolated from Indo Gangetic Soil for Enhancing Growth of Rice Crops. Microorganisms 2023, 11, 1085. https://doi.org/10.3390/microorganisms11041085
Devi S, Sharma S, Tiwari A, Bhatt AK, Singh NK, Singh M, Kaushalendra, Kumar A. Screening for Multifarious Plant Growth Promoting and Biocontrol Attributes in Bacillus Strains Isolated from Indo Gangetic Soil for Enhancing Growth of Rice Crops. Microorganisms. 2023; 11(4):1085. https://doi.org/10.3390/microorganisms11041085
Chicago/Turabian StyleDevi, Shikha, Shivesh Sharma, Ashish Tiwari, Arvind Kumar Bhatt, Nand Kumar Singh, Monika Singh, Kaushalendra, and Ajay Kumar. 2023. "Screening for Multifarious Plant Growth Promoting and Biocontrol Attributes in Bacillus Strains Isolated from Indo Gangetic Soil for Enhancing Growth of Rice Crops" Microorganisms 11, no. 4: 1085. https://doi.org/10.3390/microorganisms11041085