
Probiotic Bacteria from Human Milk Can Alleviate Oral Bovine Casein Sensitization in Juvenile Wistar Rats
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Used in This Study
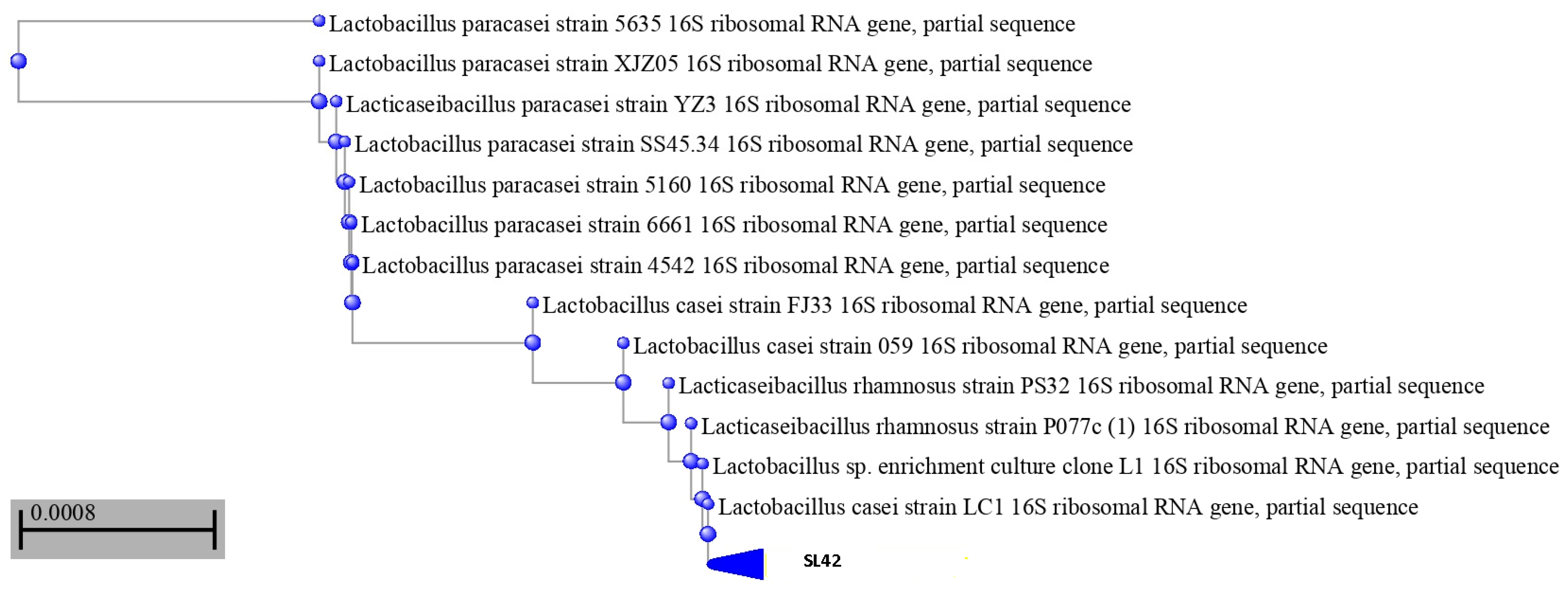
2.2. Using16S rRNA Gene Sequencing to Identify SL42 Isolate
2.3. Characterization of the Probiotic Potential
2.3.1. pH and Bile Tolerance Assays
2.3.2. Detection of Antimicrobial Activity
2.3.3. Hydrophobicity
2.3.4. Hemolytic Activity
2.3.5. Cholesterol Uptake
2.3.6. Antibiotic Susceptibility
2.4. In Vivo Study
2.4.1. Animal Housing
2.4.2. Experimental Design
2.4.3. Assessment of Macroscopic Casein Allergy Symptoms
2.4.4. Determination of Specific Casein IgE, Histamine, S100A8/A9, Inflammation-Associated Cytokines, and Eosinophil Number
2.4.5. Cultivation of Bacteria from Feces
2.4.6. Determination of Spleen/Body Weight Index and Thymus/Body Weight Index
2.4.7. Histological Analysis
2.4.8. Bacterial Translocation Test
2.5. Statistical Analysis
2.6. Ethics Approval
3. Results
3.1. SL42 Is a Lacticaseibacillus rhamnosus as Confirmed by 16S rRNA Analysis
3.2. L. rhamnosus SL42 Expresses a Satisfying Probiotic Potential
3.3. Macroscopic Symptoms Disappear after One-Week Casein Gavage
3.4. Calprotectin, Eosinophils, and Cytokines Associated with CAS- Induced Allergy Were Successfully Decreased in Plasma of Rats Gavaged with the SL42 Strain
3.5. Probiotic Administration Modifies LAB and Clostridia Populations in Rats
3.6. Probiotic Administration Does Not Change Spleen and Thymus Weights
3.7. Inflammation of Jejunal Tissue and Eosinophil Infiltration were Significantly Reduced by Probiotic Treatment
3.8. Probiotic Bacteria SL42 and DSM17938 Prevent Bacterial Translocation to Mesenteric Lymph Nodes in Wistar Rats Sensitized with Casein
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gray, C.L. Food Allergy in South Africa. Curr. Allergy Asthma Rep. 2017, 17, 668. [Google Scholar] [CrossRef] [PubMed]
- Yakhlef, M.; Souiki, L. Contribution to the estimation of the prevalence of food allergy in schoolchildren in the city of Guelma (Algeria). Rev. Fr. D’allergol. 2021, 61, 573–578. [Google Scholar] [CrossRef]
- Luyt, D.; Ball, H.; Makwana, N. BSACI guideline for the diagnosis and 463 management of cow’s milk allergy. Clin. Exp. Allergy 2014, 44, 642–672. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.A.; Fiocchi, A.; Baars, T.; Jordakieva, G.; Nowak-Wegrzyn, A.; Pali-Schöll, I.; Passanisi, S.; Pranger, C.L.; Roth-Walter, F.; Takkinen, F.; et al. Diagnosis and Rationale for Action against Cow’s Milk Allergy (DRACMA) Guidelines update-III-Cow’s milk allergens and mechanisms triggering immune activation. World Allergy Organ. J. 2022, 15, 100668. [Google Scholar] [CrossRef]
- Abdel-Gadir, A.; Stephen-Victor, E.; Gerber, G.; Rivas, M.N.; Wang, S.; Harb, H.; Wang, H.; Li, N.; Crestani, E.; Spielman, S.; et al. Microbiota therapy acts via a regulatory T cell MyD88/RORγt pathway to suppress food allergy. Nat. Med. 2019, 25, 1164–1174. [Google Scholar] [CrossRef] [PubMed]
- Iweala, O.I.; Nagler, C.R. The microbiome and food allergy. Annu. Rev. Immunol. 2019, 37, 377–403. [Google Scholar] [CrossRef]
- Zamanillo-Campos, R.; Alonso, L.C.; Martin, M.J.F.; Escusa, P.N.; Fernandez, M.T. Nutritional counseling for cow’s milk protein allergy in infants from birth to 2 y of ages: Scoping review. Nutrition 2022, 98, 111633. [Google Scholar] [CrossRef]
- Feehley, T.; Plunkett, H.; Bao, R.; Hong, S.M.; Culleen, G.; Belda-Ferre, P.; Campbell, E.; Aitoro, R.; Nocerino, R.; Paparo, L.; et al. Healthy infants harbor intestinal bacteria that protect against food allergy. Nat. Med. 2019, 25, 448–453. [Google Scholar] [CrossRef]
- Wesemann, R.; Nagler, C. The Microbiome, Timing, and Barrier Function in the Context of Allergic Disease. Immunity 2016, 44, 728–738. [Google Scholar] [CrossRef] [Green Version]
- Łubiech, K.; Twarużek, M. Lactobacillus Bacteria in Breast Milk. Nutrients 2020, 12, 3783. [Google Scholar] [CrossRef]
- Anjum, J.; Nazir, S.; Tariq, M.; Barrett, K.; Zaidi, A. Lactobacillus commensals autochthonous to human milk have the hallmarks of potent probiotics. Microbiology 2020, 166, 966–980. [Google Scholar] [CrossRef] [PubMed]
- Torii, A.; Torii, S.; Fujiwara, S.; Tanaka, H.; Inagaki, N.; Nagai, H. Lactobacillus acidophilus strain L-92 regulates the production of Th1 cytokine as well as Th2 cytokines. Allergol. Int. 2007, 56, 293–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martorell-Aragonés, A.; Echeverría-Zudaire, L.; Alonso-Lebrero, E.; Boné-Calvo, J.; Martin Muñoz, M.F.; Nevot-Falcó, S.; Piquer-Gibert, M.; Valdesoiro-Navarrete, L. Position document: IgE-mediated cow’s milk allergy. Allergol. Immunopathol. 2015, 43, 507–526. [Google Scholar] [CrossRef] [PubMed]
- An, S.B.; Yang, B.G.; Jang, G.; Kim, D.Y.; Kim, J.; Oh, S.M.; Oh, N.; Lee, S.; Moon, J.Y.; Kim, J.A.; et al. Combined IgE neutralization and Bifidobacterium longum supplementation reduces the allergic response in models of food allergy. Nat. Commun. 2022, 13, 5669. [Google Scholar] [CrossRef] [PubMed]
- Shida, K.; Takahashi, R.; Iwadate, E.; Takamizawa, K.; Yasui, H.; Sato, T.; Habu, S.; Hachimura, S.; Kaminogawa, S. Lactobacillus casei strain shirota suppresses serum immunoglobulin E and immunoglobulin G1 responses and systemic anaphylaxis in a food allergy model. Clin. Exp. Allergy 2002, 32, 563–570. [Google Scholar] [CrossRef]
- Murosaki, S.; Yamamoto, Y.; Ito, K.; Ito, K.; Inokuchi, T.; Kusaka, H.; Ikeda, H.; Yoshikai, Y. Heat-killed Lactobacillus plantarum L-137 suppresses naturally fed antigen-specific IgE production by stimulation of IL-12 production in mice. J. Allergy Clin. Immunol. 1998, 102, 57–64. [Google Scholar] [CrossRef]
- Ziar, H.; Riazi, A. Polysorbate 80 improves the adhesion and survival of yogurt starters with cholesterol uptake abilities. Saudi J. Biol. Sci. 2022, 29, 103367. [Google Scholar] [CrossRef]
- Ziar, H.; Gérard, P.; Riazi, A. Effect of prebiotic carbohydrates on growth, Bile survival and Cholesterol-lowering activities of dairy-related bacteria. Int. J. Food Sci. Technol. 2014, 47, 1421–1429. [Google Scholar] [CrossRef]
- Voulgari, K.; Hatzikamari, M.; Delepoglou, A.; Georgakopoulos, P.; Litopoulou-Tzanetaki, E.; Tzanetakis, N. Antifungal activity of non-starter lactic acid bacteria isolates from dairy products. Food Control 2010, 21, 136–142. [Google Scholar] [CrossRef]
- Pérez, P.F.; Minnaard, Y.; Disalvo, E.A.; De Antoni, G.L. Surface properties of bifidobacterial strains of human origin. Appl. Environ. Microbiol. 1998, 64, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Buxton, R. Blood Agar Plates and Hemolysis Protocols. 2005. Available online: https://www.asm.org/getattachment/7ec0de2b-bb16-4f6e-ba072aea25a43e76/protocol2885.pdf (accessed on 19 November 2019).
- Ziar, H.; Yahla, I.; Riazi, A.; Gérard, P. Ca-Alginate-Carob Galactomannans Beads to Preserve Viability During Digestive Hostility Transit and Cholesterol Uptake Ability of Probiotic Bacteria. In Proceedings of the 4th International Symposium on Materials and Sustainable Development, ISMSD 2019; Benmounah, A., Abadlia, M.T., Saidi, M., Zerizer, A., Eds.; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Pereira, D.I.; McCartney, A.L.; Gibson, G.R. An in vitro study of the probioticpotential of a bile-salt-hydrolyzing Lactobacillus fermentum strain, and determination of its cholesterol-lowering properties, Appl. Environ. Microbiol. 2003, 69, 4743–4752. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Zhang, R.; Tian, X.; Zhou, X.; Pan, X.; Wong, A. Assessing the risk of probiotic dietary supplements in the context of antibiotic resistance. Front. Microbiol. 2017, 8, 908. [Google Scholar] [CrossRef] [PubMed]
- Plessas, S.; Nouska, C.; Karapetsas, A.; Kazakos, S.; Alexopoulos, A.; Mantzourani, I.; Chondrou, P.; Fournomiti, M.; Galanis, A.; Bezirtzoglou, E. Isolation, characterization and evaluation of the probiotic potential of a novel Lactobacillus strain isolated from Feta-type cheese. Food Chem. 2017, 226, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Cheng, J.; Deng, M.C.; Chou, C.H.; Jan, T.R. Prebiotic effect of diosgenin, an immunoactive steroidal sapogenin of the Chinese yam. Food Chem. 2012, 132, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Hink, T.; Burnham, C.A.; Dubberke, E.R. A systematic evaluation of methods to optimize culture-based recovery of Clostridium difficile from stool specimens. Anaerobe 2013, 19, 39–43. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Zhang, S.; Zhang, L.; Li, B. Effect of orally administered collagen peptides from bovine bone on skin aging in chronologically aged mice. Nutrients 2017, 9, 1209. [Google Scholar] [CrossRef] [Green Version]
- Stanojević, S.; Blagojević, V.; Ćuruvija, I.; Veljović, K.; Bajić, S.S.; Kotur-Stevuljević, J.; Bogdanović, A.; Petrović, R.; Vujnović, I.; Kovačević-Jovanović, V. Oral treatment with Lactobacillus rhamnosus 64 during the early postnatal period improves the health of adult rats with TNBS-induced colitis. J. Funct. Foodsn 2018, 48, 92–105. [Google Scholar] [CrossRef]
- Muraro, A.; Halken, S.; Arshad, S.H.; Beyer, K.; Dubois, A.E.; Du Toit, G.; Eigenmann, P.A.; Grimshaw, K.E.; Hoest, A.; Lack, G.; et al. EAACI food allergy and anaphylaxis guidelines. Primary prevention of food allergy. Allergy 2014, 69, 590–601. [Google Scholar] [CrossRef]
- Tulyeu, J.; Kumagai, H.; Jimbo, E.; Watanabe, S.; Yokoyama, K.; Cui, L.; Osaka, H.; Mieno, M.; Yamagata, T. Probiotics Prevents Sensitization to Oral Antigen and Subsequent Increases in Intestinal Tight Junction Permeability in Juvenile-Young Adult Rats. Microorganisms 2019, 7, 463. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.; Pan, L.; Peng, C.; Dong, L.; Cao, S.; Cheng, H.; Wang, Y.; Zhang, C.; Gu, R.; Wang, J.; et al. Isolation and characterization of lactic acid bacteria from human milk. J. Dairy Sci. 2020, 103, 9980–9991. [Google Scholar] [CrossRef]
- Moossavi, S.; Azad, M.B. Origins of human milk microbiota: New evidence and arising questions. Gut Microbes 2019, 12, 1667722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campedelli, I.; Mathur, H.; Salvetti, E.; Rea, M.C.; Torriani, S.; Ross, R.P.; Hill, C.; O’Toole, P.W. Genus-wide assessment of antibiotic resistance in Lactobacillus spp. Appl. Environ. Microbiol. 2019, 85, e01738-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Oh, J.-H.; Alexander, L.M.; Ozçam, M.; Van Pijkeren, J.P. D-alanyl-dalanine ligase as a broad-host-range counter selection marker in vancomycin resistant lactic Acid Bacteria. J. Bacteriol. 2018, 200, e00607–e00617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saviano, A.; Mattia Brigida, O.; Migneco, A.; Gunawardena, G.; Zanza, C.; Candelli, M.; Franceschi, F.; Ojetti, V. Lactobacillus Reuteri DSM 17938 (Limosilactobacillus reuteri) in Diarrhea and Constipation: Two Sides of the Same Coin? Medicina 2021, 57, 643. [Google Scholar] [CrossRef]
- Pilegaard, K.; Madsen, C. An oral Brown Norway rat model for food allergy: Comparison of age, sex, dosing volume, and allergen preparation. Toxicology 2004, 196, 247–257. [Google Scholar] [CrossRef]
- Yan, F.; Liu, L.; Cao, H.; Moore, D.J.; Washington, M.K.; Wang, B.; Peek, R.; Acra, S.; Polk, D. Neonatal colonization of mice with LGG promotes intestinal development and decreases susceptibility to colitis in adulthood. Mucosal Immunol. 2017, 10, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Neau, E.; Delannoy, J.; Marion, C.; Cottart, C.H.; Labellie, C.; Holowacz, S.; Butel, M.-J.; Kapel, N.; Waligora-Dupriet, A.-J. Three novel candidate probiotic strains with prophylactic properties in a murine model of cow’s milk allergy. Appl. Environ. Microbiol. 2016, 82, 1722–1733. [Google Scholar] [CrossRef] [Green Version]
- Esber, N.; Mauras, A.; Delannoy, J.; Labellie, C.; Mayeur, C.; Caillaud, M.A.; Kashima, T.; Souchaud, L.; Nicolis, I.; Kapel, N.; et al. Three candidate probiotic strains impact gut microbiota and induce anergy in mice with cow’s milk allergy. Appl. Environ. Microbiol. 2020, 86, e01203–e01220. [Google Scholar] [CrossRef]
- Okada, K.; Itoh, H.; Ikemoto, M. Circulating S100A8/A9 is potentially a biomarker that could reflect the severity of experimental colitis in rats. Heliyon 2020, 6, e03470. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Li, F.; Wang, J.; Ma, J.; Sheng, X. Upregulation of calprotectin in mild IgE-mediated ovalbumin hypersensitivity. Oncotarget 2017, 8, 37342–37354. [Google Scholar] [CrossRef] [Green Version]
- Pham, D.L.; Yoon, M.G.; Ban, G.Y.; Kim, S.H.; Kim, M.A.; Ye, Y.M.; Shin, Y.S.; Park, H.S. Serum S100A8 and S100A9 Enhance Innate Immune Responses in the Pathogenesis of Baker’s Asthma. Int. Arch. Allergy Immunol. 2015, 168, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Takahashi, N.; Kato, T.; Matsuda, Y.; Yokoji, M.; Yamada, M.; Nakajima, T.; Kondo, N.; Endo, N.; Yamamoto, R.; et al. Aggravation of collagen-induced arthritis by orally administered Porphyromonas gingivalis through modulation of the gut microbiota and gut immune system. Sci. Rep. 2017, 7, 6955. [Google Scholar] [CrossRef] [PubMed] [Green Version]





 , MRS-cys
, MRS-cys  , TSA II
, TSA II  , Hektoen
, Hektoen  ) are presented as the mean ± SD. Statistical analysis was conducted by using one-way ANOVA with Tukey’s multiple comparisons test. * p < 0.05. Control group receiving only PBS (control group); nonsensitized group treated with SL42 strain (SL42 group); nonsensitized group treated with DSM 17938 strain (DSM 17938 group); casein-sensitized group (casein group); casein-sensitized group treated with SL42 (casein + SL42 group); and casein-sensitized group treated with DSM 17938 strain (casein- + DSM 17938 group).
, MRS-cys , TSA II , Hektoen ) are presented as the mean ± SD. Statistical analysis was conducted by using one-way ANOVA with Tukey’s multiple comparisons test. * p < 0.05. Control group receiving only PBS (control group); nonsensitized group treated with SL42 strain (SL42 group); nonsensitized group treated with DSM 17938 strain (DSM 17938 group); casein-sensitized group (casein group); casein-sensitized group treated with SL42 (casein + SL42 group); and casein-sensitized group treated with DSM 17938 strain (casein- + DSM 17938 group).
) are presented as the mean ± SD. Statistical analysis was conducted by using one-way ANOVA with Tukey’s multiple comparisons test. * p < 0.05. Control group receiving only PBS (control group); nonsensitized group treated with SL42 strain (SL42 group); nonsensitized group treated with DSM 17938 strain (DSM 17938 group); casein-sensitized group (casein group); casein-sensitized group treated with SL42 (casein + SL42 group); and casein-sensitized group treated with DSM 17938 strain (casein- + DSM 17938 group).
, MRS-cys , TSA II , Hektoen ) are presented as the mean ± SD. Statistical analysis was conducted by using one-way ANOVA with Tukey’s multiple comparisons test. * p < 0.05. Control group receiving only PBS (control group); nonsensitized group treated with SL42 strain (SL42 group); nonsensitized group treated with DSM 17938 strain (DSM 17938 group); casein-sensitized group (casein group); casein-sensitized group treated with SL42 (casein + SL42 group); and casein-sensitized group treated with DSM 17938 strain (casein- + DSM 17938 group).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| L. rhamnosus SL42 | L. reuteri DSM 17938 | |
|---|---|---|
| Parameters | Cell Viability/Activity | |
| Acid resistance (pH = 2) | 2 × 107 CFU/mL (0 min) | 5 × 107 CFU/mL (0 min) |
| 7.5 × 106 CFU/mL (30 min) | 2.3 × 107 CFU/mL (30 min) | |
| 6.9 × 106 CFU/ mL (60 min) | 1.9 × 106 CFU/mL (60 min) | |
| 6.6 × 106 CFU/mL (120 min) | 1.2 × 106 CFU/mL (120 min) | |
| Bile survival (0.3%) | 5 × 107 CFU/mL (0 min) | 2.3 × 107 CFU/mL (0 min) |
| 1.4 × 107 CFU/mL (4 h) | 2.1 × 106 CFU/mL (4 h) | |
| 9.3 × 106 CFU/mL (24 h) | 1.2 × 106 CFU/mL (24 h) | |
| Cholesterol uptake | 6.01 mg/g | 6.09 mg/g |
| Hydrophobicity (%) | 51% | 76% |
| Hemolytic activity | γ-hemolytic (no hemolysis) | γ-hemolytic (no hemolysis) |
| Pathogens | Inhibition Zone (mm) | |
|---|---|---|
| L. rhamnosus SL42 | L. reuteri DSM 17938 | |
| Candida albicans ATCC 10231 | 14 ± 0.18 | 13 ± 0.36 |
| Escherichia coli ATCC 25922 | 18 ± 0.02 | 14 ± 0.10 |
| Bacillus cereus ATCC 10876 | 9 ± 0.11 | 6± 0.10 |
| Staphylococcus aureus ATCC 33862 | 15 ± 0.30 | 8 ± 0.20 |
| Pseudomonas aeruginosa ATCC 27853 | 17 ± 0.22 | 15 ± 0.20 |
| Salmonella enterica subsp. enterica serotype Enteritidis ATCC 4931 | 9 ± 0.08 | 16 ± 0.04 |
| Klebsiella pneumoniae ATCC 13883 | 11 ± 0.02 | 14 ± 0.20 |
| Shigella sp. (isolate from our collection) | 5 ± 0.02 | 6 ± 0.10 |
| Probiotic Bacteria | Antibiotic Susceptibility (mm) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Amoxicillin (30 µg) | Ciprofloxacin (5 µg) | Gentamicin (10 µg) | Penicillin (10 µg) | Sulfonamide (25 µg) | Streptomycin (10 µg) | Clindamycin (2 µg) | Chloramphenicol (30 µg) | Vancomycin (30 µg) | Norfloxacin (10 µg) | Novobiocin (4 µg) | |
| L. rhamnosus SL42 | 15 ± 0.1 I | 0 ± 0.01 R | 10 ± 0.02 R | 20 ± 0.01 S | 0 ± 0.1 R | 0 ± 0.1 R | 0 ± 0.0 R | 25 ± 0.1 S | 8 ± 0.03 R | 16 ± 0.1 I | 22 ± 0.02 S |
| L. reuteri DSM 17938 | 0 ± 0.01 R | 16 ± 0.02 I | 8 ± 0.01 R | 15 ± 0.01 I | 0 ± 0.1 R | 16 ± 0.1 I | 18 ± 0.0 I | 16 ± 0.1 I | 12 ± 0.02 R | 16 ± 0.1 I | 22 ± 0.0 S |
| Group | Number of Affected Rats | CFU/ MLN of Rat |
|---|---|---|
| Control | 0/8 | 0 |
| SL42 | 0/8 | 0 |
| DSM17938 | 0/8 | 0 |
| Casein | 6/8 | 288 * |
| Casein + SL42 | 0/8 | 0 |
| Casein + DSM17938 | 0/8 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keddar, K.; Ziar, H.; Belmadani, N.; Monnoye, M.; Gérard, P.; Riazi, A. Probiotic Bacteria from Human Milk Can Alleviate Oral Bovine Casein Sensitization in Juvenile Wistar Rats. Microorganisms 2023, 11, 1030. https://doi.org/10.3390/microorganisms11041030
Keddar K, Ziar H, Belmadani N, Monnoye M, Gérard P, Riazi A. Probiotic Bacteria from Human Milk Can Alleviate Oral Bovine Casein Sensitization in Juvenile Wistar Rats. Microorganisms. 2023; 11(4):1030. https://doi.org/10.3390/microorganisms11041030
Chicago/Turabian StyleKeddar, Kawtar, Hasnia Ziar, Noussaiba Belmadani, Magali Monnoye, Philippe Gérard, and Ali Riazi. 2023. "Probiotic Bacteria from Human Milk Can Alleviate Oral Bovine Casein Sensitization in Juvenile Wistar Rats" Microorganisms 11, no. 4: 1030. https://doi.org/10.3390/microorganisms11041030