
A Morphometric Approach to Understand Prokaryoplankton: A Study in the Sicily Channel (Central Mediterranean Sea)
 ,
,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Sampling
2.2. Environmental Variables
2.3. Microbial Variables
2.4. Statistical Analyses
3. Results
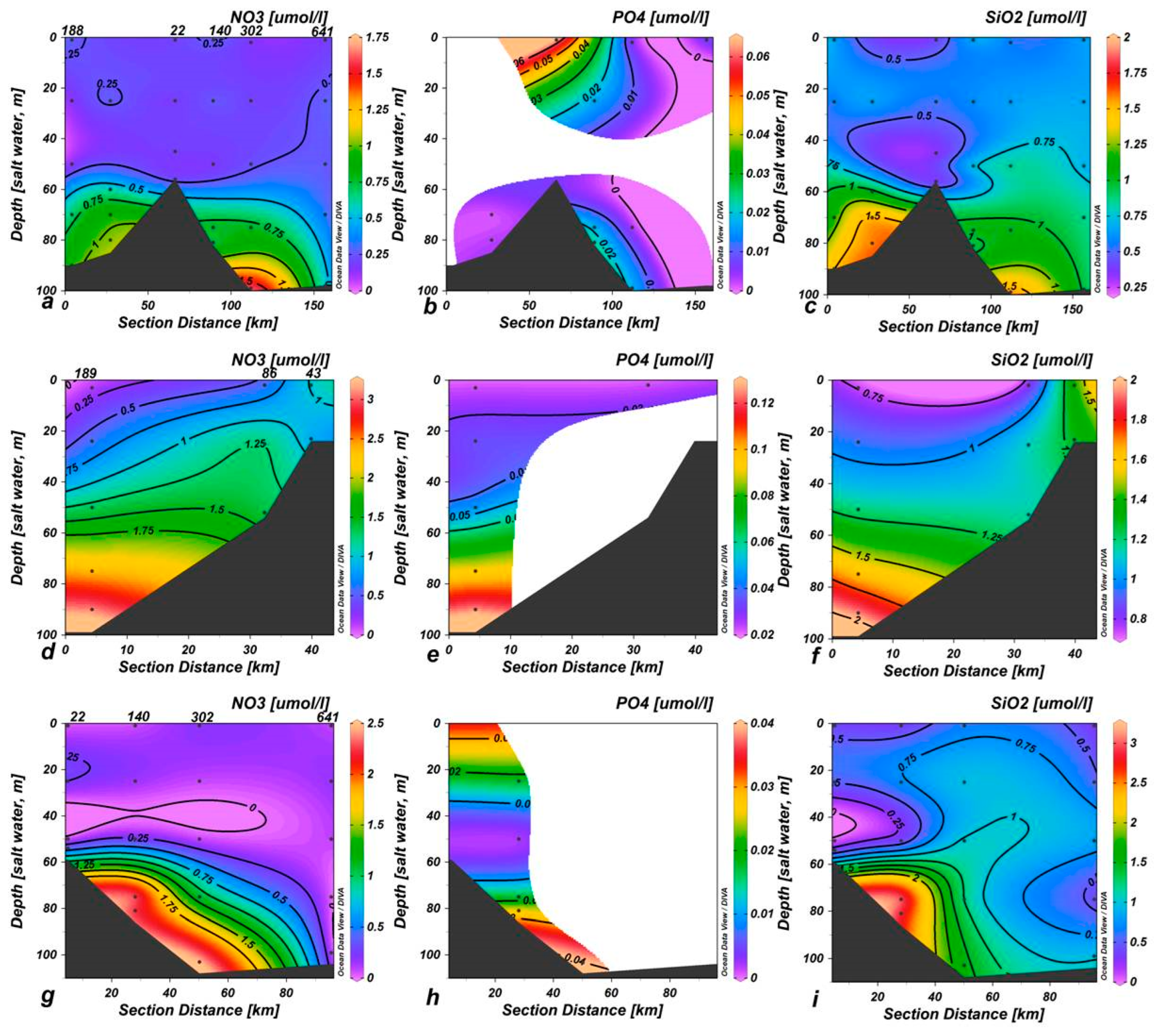
3.1. Hydrological and Chemical Features
3.2. Microbial Variables
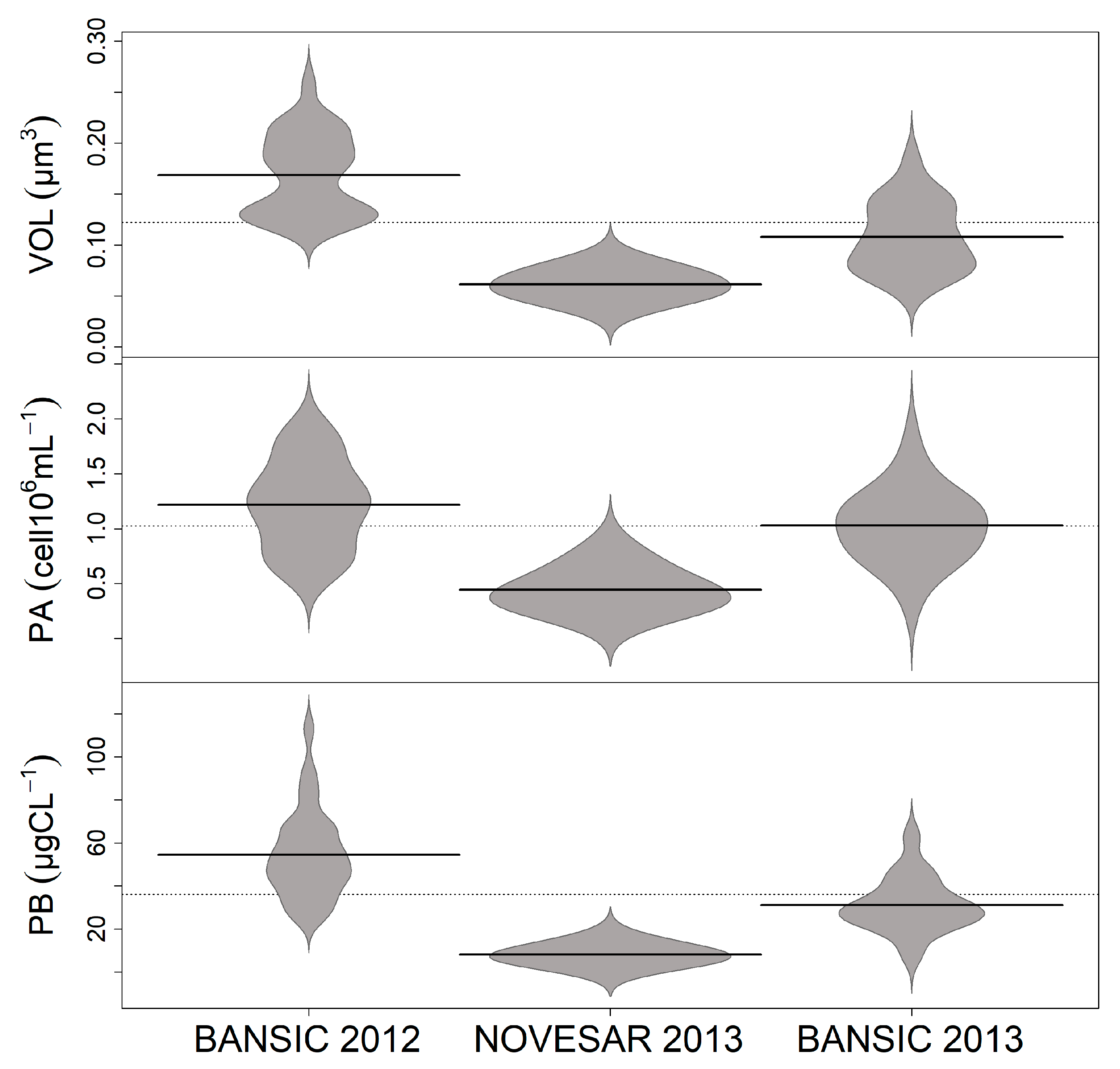
3.3. Cell Morphotypes and Their Morphometry
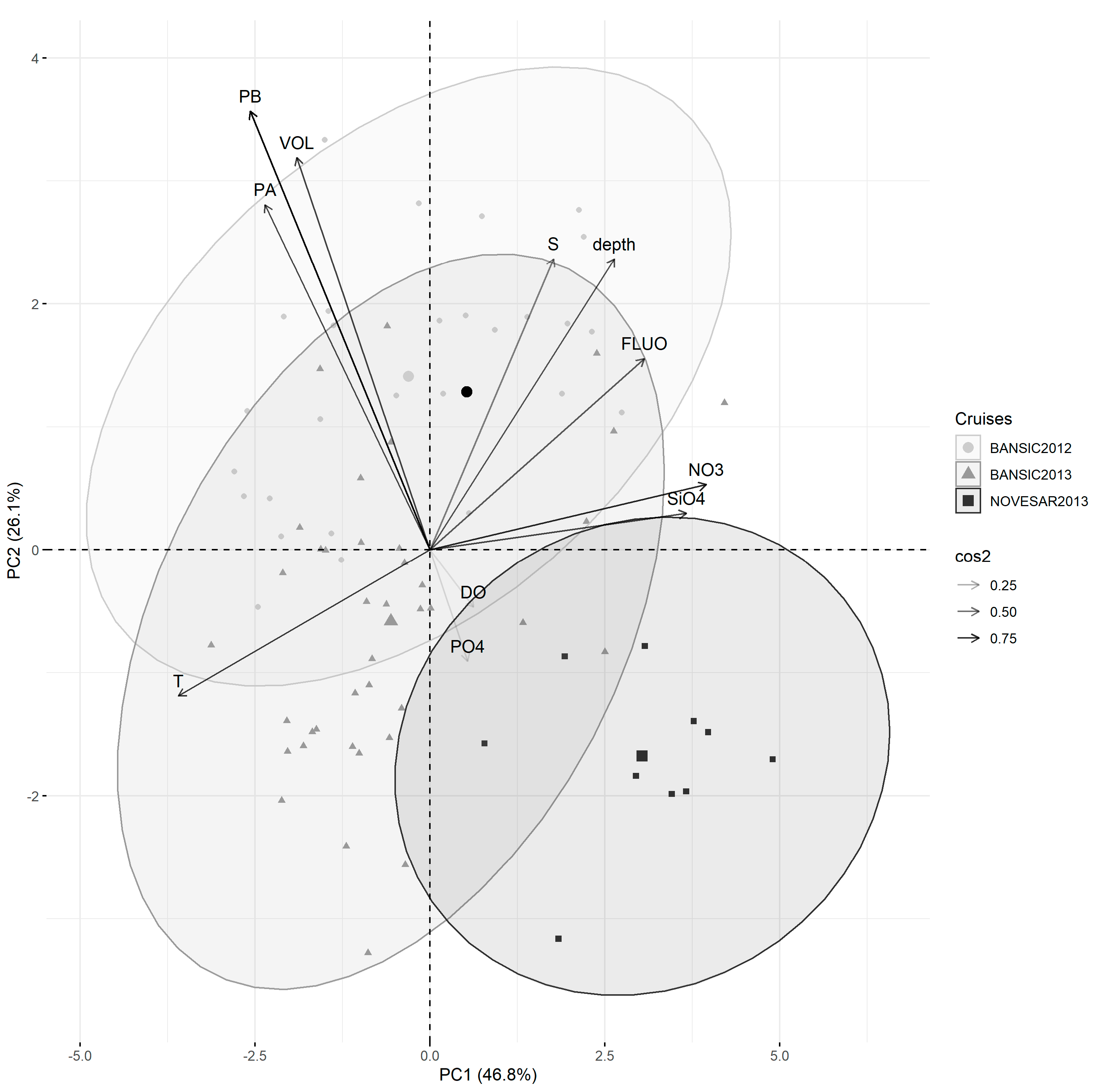
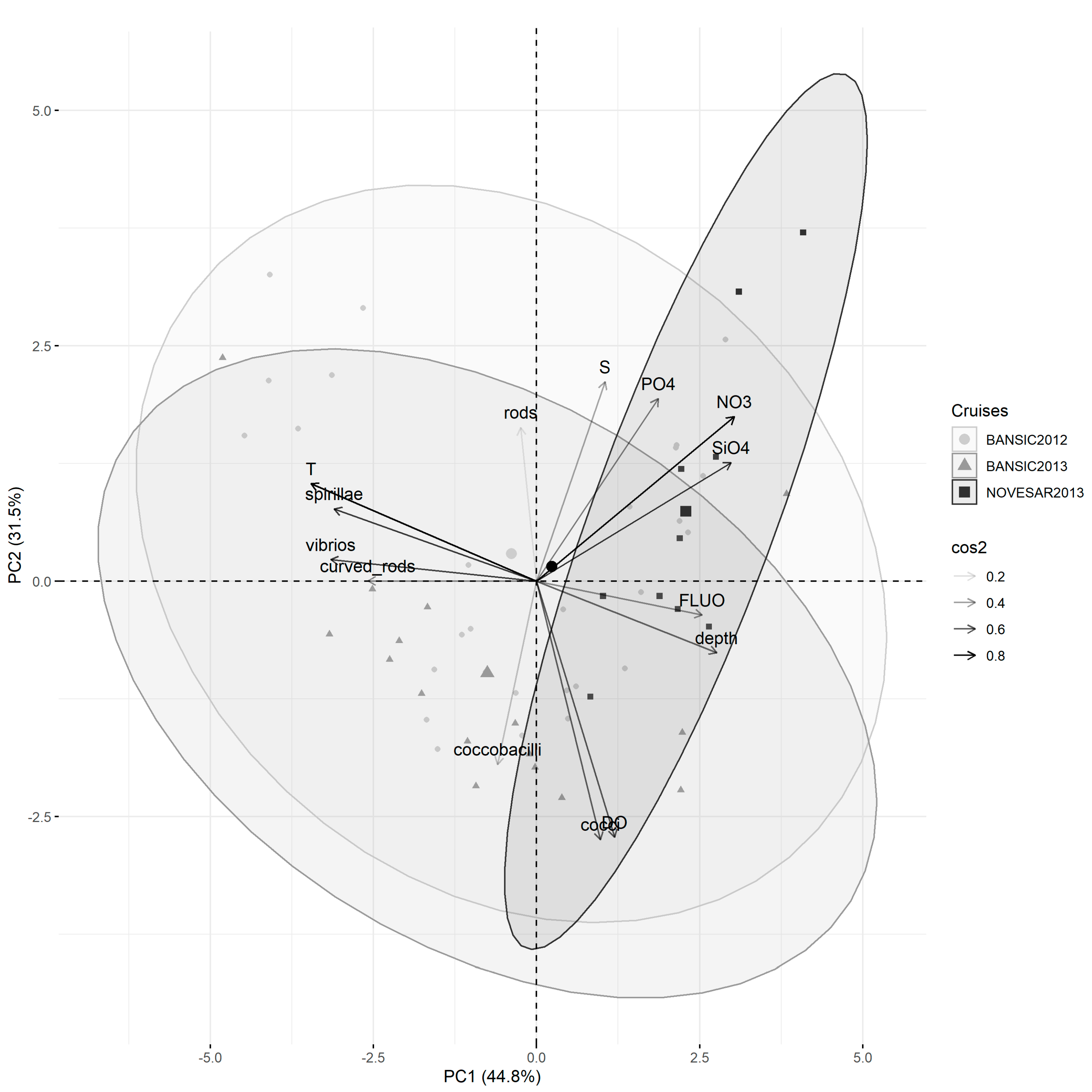
3.4. Relations between Prokaryotic Features and Environmental Variables
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kleparski, L.; Beaugrand, G.; Edwards, M.; Schmitt, F.G.; Kirby, R.R.; Breton, E.; Gevaert, F.; Maniez, E. Morphological traits, niche-environment interaction and temporal changes in diatoms. Progr. Ocean. 2022, 201, 102747. [Google Scholar] [CrossRef]
- Mahadevaswamy, M.; Yamakanamardi, S.M.; Harsha, T.S. Bacterial (free-living and particle bound) cell-size in the surface waters of river Cauvery and its upstream tributaries in Karnataka State, India. Appl. Ecol. Environ. Res. 2008, 6, 29–47. [Google Scholar] [CrossRef]
- Pernthaler, J.; Amann, R. Fate of heterotrophic microbes in pelagic habitats: Focus on populations. Microbiol. Mol. Biol. Rev. 2005, 6, 440–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalcheva, H.; Beshkova, M.; Pehlivanov, L.; Kalchev, R. Bacterioplankton Dynamics and the Influence of Environmental Factors on It in the Srebarna Lake. In Proceedings of the Third International Scientific Conference BALWOIS, Ohrid, North Macedonia, 27–31 May 2008; pp. 1–9. [Google Scholar]
- Hessen, D.O.; Daufresne, M.; Leinaas, H.P. Temperature-size relations from the cellular-genomic perspective. Biol. Rev. 2012, 88, 476–489. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.S.; Huszar, V.; Kruk, C.; Naselli-Flores, L.; Melo, S. Towards a functional classification of the freshwater phytoplankton. J. Plank. Res. 2002, 24, 417–428. [Google Scholar] [CrossRef]
- Acevedo-Trejos, E.; Brandt, G.; Bruggeman, J.; Merico, A. Mechanisms shaping size structure and functional diversity of phytoplankton communities in the ocean. Sci. Rep. 2015, 5, 8918. [Google Scholar] [CrossRef] [Green Version]
- Sourisseau, M.; Le Guennec, V.; Le Gland, G.; Plus, M.; Chapelle, A. Resource Competition Affects Plankton Community Structure Evidence from Trait-Based Modeling. Front. Mar. Sci. 2017, 4, 52. [Google Scholar] [CrossRef] [Green Version]
- Kruk, C.; Devercelli, M.; Vera, L. Reynolds Functional Groups: A trait-based pathway from patterns to predictions. Hydrobiologia 2021, 848, 113–129. [Google Scholar] [CrossRef]
- Tamburini, C.; Garel, M.; Badr, A.A.; Mérigot, B.; Kriwy, P.; Charrière, B.; Budillon, G. Distribution and activity of prokaryotes in the water column of the Tyrrhenian Sea. Deep-Sea Res. II 2009, 56, 700–712. [Google Scholar] [CrossRef]
- Sousa, A.M.; Machado, I.; Pereira, M.O. Phenotypic switching: An opportunity to bacteria thrive. In Science Against Microbial Pathogens: Communicating Current Research and Technological Advances; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2011; pp. 252–262. ISBN 978-84-939843-1-1. [Google Scholar]
- Andersen, K.H.; Berge, T.; Gonçalves, R.J.; Hartvig, M.; Heuschele, J.; Hylander, S.; Jacobsen, N.S.; Lindermann, C.; Martens, E.A.; Neuheimer, A.B.; et al. Characteristic sizes of life in the oceans, from bacteria to whales. Annu. Rev. Mar. Sci. 2016, 8, 217–241. [Google Scholar] [CrossRef] [Green Version]
- Young, K.D. The selective value of bacterial shape. Microbiol. Mol. Biol. Rev. 2006, 70, 660–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straza, T.R.A.; Cottrell, M.T.; Ducklow, H.W.; Kirchman, D.L. Geographic and phylogenetic variation in bacterial biovolume as revealed by protein and nucleic acid staining. Appl. Environ. Microbiol. 2009, 75, 4028–4034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentile, G.; Maimone, G.; La Ferla, R.; Azzaro, M.; Catalfamo, M.; Genovese, M.; Santisi, S.; Maldani, M.; Macrì, A.; Cappello, S. Phenotypic variations of Oleispira antarctica RB 8(T) in different growth conditions. Curr. Microbiol. 2020, 77, 3414–3421. [Google Scholar] [CrossRef]
- Kundu, K.; Weber, N.; Grieblera, C.; Elsnera, M. Phenotypic heterogeneity as key factor for growth and survival under oligotrophic conditions. Environ. Microbiol. 2020, 22, 3339–3356. [Google Scholar] [CrossRef] [PubMed]
- La Ferla, R.; Maimone, G.; Caruso, G.; Azzaro, F.; Azzaro, M.; Decembrini, F.; Cosenza, A.; Leonardi, M.; Paranhos, R. Are prokaryotic cell shape and size suitable to ecosystem characterization? Hydrobiologia 2014, 726, 65–80. [Google Scholar] [CrossRef]
- Zaccone, R.; Azzaro, M.; Caruso, G.; Crisafi, E.; Decembrini, F.; Leonardi, M.; Maimone, G.; Monticelli, L.; La Ferla, R. Effects of climate changes on the microbial activities and prokaryotic abundances in the euphotic layer of the Central Mediterranean Sea. Hydrobiologia 2019, 842, 5–30. [Google Scholar] [CrossRef]
- Azzaro, M.; Maimone, G.; La Ferla, R.; Cosenza, A.; Rappazzo, A.C.; Caruso, G.; Paranhos, R.; Cabral, A.S.; Forte, E.; Guglielmin, M. The prokaryotic community in an extreme Antarctic environment: The brines of Boulder Clay lakes (Northern Victoria Land). Hydrobiologia 2021, 848, 1837–1857. [Google Scholar] [CrossRef]
- La Ferla, R.; Azzaro, M.; Budillon, G.; Caroppo, C.; Decembrini, F.; Maimone, G. Distribution of the prokaryotic biomass and community respiration in the main water masses of the Southern Tyrrhenian Sea (June and December 2005). Adv. Oceanogr. Limnol. 2010, 1, 235–257. [Google Scholar] [CrossRef]
- Azzaro, M.; La Ferla, R.; Maimone, G.; Monticelli, L.S.; Zaccone, R.; Civitarese, G. Prokaryotic dynamics and heterotrophic metabolism in a deep convection site of Eastern Mediterranean Sea (the Southern Adriatic Pit). Cont. Shelf. Res. 2012, 44, 106–118. [Google Scholar] [CrossRef]
- Placenti, F.; Azzaro, M.; Artale, V.; La Ferla, R.; Caruso, G.; Santinelli, C.; Maimone, G.; Monticelli, L.S.; Quinci, M.; Sprovieri, M. Biogeochemical patterns and microbial processes in the Eastern Mediterranean Deep Water of Ionian Sea. Hydrobiologia 2018, 815, 97–112. [Google Scholar] [CrossRef]
- Patti, B.; Guisande, C.; Bonanno, A.; Basilone, G.; Cuttitta, A.; Mazzola, S. Role of physical forcing and nutrient availability on the control of satellite-based chlorophyll a concentration in the coastal upwelling area of the Sicilian channel. Sci. Mar. 2010, 74, 577–588. [Google Scholar] [CrossRef] [Green Version]
- Placenti, F.; Schroeder, K.; Bonanno, A.; Zgozi, S.; Sprovieri, M.; Borghini, M.; Rumolo, P.; Cerati, G.; Bonomo, S.; Genovese, S.; et al. Water masses and nutrient distribution in the Gulf of Syrte and between Sicily and Libya. J. Mar. Syst. 2013, 121–122, 36–46. [Google Scholar] [CrossRef]
- Placenti, F.; Torri, M.; Pessini, F.; Patti, B.; Tancredi, V.; Cuttitta, A.; Giaramita, L.; Tranchida, G.; Sorgente, R. Hydrological and biogeochemical patterns in the Sicily Channel: New insights from the last decade (2010–2020). Front. Mar. Sci. 2022, 9, 33540. [Google Scholar] [CrossRef]
- Robinson, A.R.; Sellschopp, J.; Warn-Varnas, A.; Leslie, W.G.; Lozano, C.J.; Haley, P.J., Jr.; Anderson, L.A.; Lermusiaux, P.F.J. The Atlantic Ionian Stream. J. Mar. Syst. 1999, 20, 129–156. [Google Scholar] [CrossRef]
- Basilone, G.; Bonanno, A.; Patti, B.; Mazzola, S.; Barra, M.; Cuttitta, A.; McBride, R. Spawning site selection by European anchovy (Engraulis encrasicolus) in relation to oceanographic conditions in the Strait of Sicily. Fish. Oceanog. 2013, 22, 309–323. [Google Scholar] [CrossRef]
- Patti, B.; Zarrad, R.; Jarboui, O.; Cuttitta, A.; Basilone, G.; Aronica, S.; Placenti, F.; Tranchide, G.; Armeri, G.M.; Buffa, G.; et al. Anchovy (Engraulis encrasicolus) early life stages in the Central Mediterranean Sea: Connectivity issues emerging among adjacent sub-areas across the Strait of Sicily. Hydrobiologia 2018, 821, 25–40. [Google Scholar] [CrossRef]
- Decembrini, F.; Caroppo, C.; Azzaro, M. Size structure and production of phytoplankton community and carbon pathways channelling in the Southern Tyrrhenian Sea (Western Mediterranean). Deep Sea Res. Part II Top Stud. Oceanogr. 2009, 56, 687–699. [Google Scholar] [CrossRef]
- Grasshoff, K.; Kremling, K.; Ehrhardt, M. Methods of Seawater Analysis; Wiley-VCH Verlag GmbH: Weinheim, Germany, 1999; ISBN 9783527295890. [Google Scholar]
- La Ferla, R.; Maimone, G.; Azzaro, M.; Conversano, F.; Brunet, C.; Cabral, A.S.; Paranhos, R. Vertical distribution of the prokaryotic cell size in the Mediterranean Sea. Helgol. Mar. Res. 2012, 66, 635–650. [Google Scholar] [CrossRef] [Green Version]
- Turley, C.M.; Hughes, D.J. Effects of storage on direct estimates of bacterial numbers of preserved seawater samples. Deep-Sea Res. 1992, 39, 375–394. [Google Scholar] [CrossRef]
- Porter, K.G.; Feig, Y.S. The use of DAPI for identifying and counting aquatic microflora. Limnol. Oceanogr. 1980, 25, 943–948. [Google Scholar] [CrossRef]
- Posch, T.; Franzoi, J.; Prader, M.; Salcher, M.M. New image analysis tool to study biomass and morphotypes of three major bacterioplankton groups in an alpine lake. Aquat. Microb. Ecol. 2009, 54, 113–126. [Google Scholar] [CrossRef] [Green Version]
- Loferer-Krößbacher, M.; Klima, J.; Psenner, R. Determination of bacterial cell dry mass by 680 transmission electron microscopy and densitometric image analysis. App. Environ. Microbiol. 1998, 64, 688–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeder, M.; Kohler, E.; Zeder, L.; Pernthaler, J. A novel algorithm for the determination of bacterial cell 775 volumes that is unbiased by cell morphology. Microsc. Microanal. 2011, 17, 799–809. [Google Scholar] [CrossRef] [Green Version]
- Brussaard, C.P.D. Optimization of procedures for counting viruses by flow cytometry. Appl. Environ. Microbiol. 2004, 70, 1506–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampstra, P. A boxplot alternative for visual comparison of distributions. J. Stat. Softw. 2008, 28, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; Plymouth Marine Laboratory: Plymouth, MA, USA, 2006. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Jolliffe, I. Principal component analysis. In Encyclopedia of Statistics in Behavioral Science; John Wiley & Sons, Ltd.: London, UK, 2005. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin, Germany, 2016. [Google Scholar]
- Kassambara, A.; Mundt, F. Package ‘Factoextra’. Extr. Vis. Results Multivar Data Anal 76. 2017. Available online: https://cran.microsoft.com/snapshot/2016-11-30/web/packages/factoextra/factoextra.pdf (accessed on 13 October 2022).
- Wentzky, V.C.; Tittel, J.; Jäger, C.G.; Bruggeman, J.; Rinke, K. Seasonal succession of functional traits in phytoplankton communities and their interaction with trophic state. J. Ecol. 2020, 101, 1649–1663. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.-Q.; Jian, S.-L.; Li, K.-M.; Wu, Z.-B.; Guan, H.-T.; Hao, J.-W.; Wang, S.-Y.; Lin, Y.-Y.; Wang, G.-J.; Li, A.-H. Community structure of bacterioplankton and its relationship with environmental factors in the upper reaches of the Heihe River in Qinghai Plateau. Envirn. Microbiol. 2020, 23, 1210–1221. [Google Scholar]
- Bonanno, A.; Placenti, F.; Basilone, G.; Mifsud, R.; Genovese, S.; Patti, B.; Di Bitetto, M.; Aronica, S.; Barra, M.; Giacalone, G.; et al. Variability of water mass properties in the Strait of Sicily in summer period of 1998–2013. Ocean. Sci. 2014, 10, 759–770. [Google Scholar] [CrossRef] [Green Version]
- Cuttitta, A.; Quinci, E.M.; Patti, B.; Bonomo, S.; Bonanno, A.; Musco, M.; Torri, M.; Placenti, F.; Basilone, G.; Genovese, S.; et al. Different key roles of mesoscale oceanographic structures and ocean bathymetry in shaping larval fish distribution pattern: A case study in Sicilian waters in summer 2009. J. Sea. Res. 2016, 115, 6–17. [Google Scholar] [CrossRef]
- Gocke, K.; Hernández, C.; Giesenhagen, H.; Hoppe, H.G. Seasonal variations of bacterial abundance and biomass and their relation to phytoplankton in the hypertrophic tropical lagoon Ciénaga Grande de Santa Marta, Colombia. J. Plank. Res. 2004, 26, 1429–1439. [Google Scholar] [CrossRef] [Green Version]
- Sjöstedt, J.; Hagström, Å.; Li Zweifel, U. Variation in cell volume and community composition of bacteria in response to temperature. Aquat. Microb. Ecol. 2012, 66, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Araújo, M.F.F.; Godinho, M.J.L. Seasonal and spatial distribution of bacterioplankton in a fluvial-lagunar system of a tropical region: Density, biomass, cellular volume and morphologic variation. Braz. Arch. Biol. Technol. 2008, 51, 205–214. [Google Scholar] [CrossRef]
- Morán, X.A.G.; Alonso-Sáez, L.; Nogueira, E.; Ducklow, H.W.; Gonzalez, N.; Lòpez-Urrutia, A.; Diaz-Perez, L.; Calvo-Diaz, A.; Arandia-Gorostidi, N.; Huete-Stauffer, T.M. More, smaller bacteria in response to ocean’s warming? Proc. R. Soc. B 2015, 282, 20150371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeder, K.; Gasparini, G.P.; Borghini, M.; Cerrati, G.; Delfanti, R. Biogeochemical tracers and fluxes in the Western Mediterranean Sea, spring 2005. J. Mar. Syst. 2010, 80, 8–24. [Google Scholar] [CrossRef]
- Zaccone, R.; Azzaro, M.; Azzaro, F.; Caruso, G.; Caroppo, C.; Decembrini, F.; Diociaiuti, T.; Fonda Umani, S.; Leonardi, M.; Maimone, G.; et al. Trophic structure and microbial activity in a spawning area of Engraulis encrasicolus. Estuar. Coast. Shelf. Sci. 2018, 207, 215–222. [Google Scholar] [CrossRef]
- Quinones, R.A.; Platt, T.; Rodrìguez, J. Patterns of biomass-size spectra from oligotrophic waters of the Northwest Atlantic. Prog. Oceanogr. 2003, 570, 405–427. [Google Scholar] [CrossRef]
- La Ferla, R.; Azzaro, M.; Caruso, G.; Monticelli, L.S.; Maimone, G.; Zaccone, R.; Packard, T.T. Prokaryotic abundance and heterotrophic metabolism in the deep Mediterranean Sea. Adv. Oceanogr. Limnol. 2010, 1, 143–166. [Google Scholar] [CrossRef]
- Zaccone, R.; Azzaro, M.; Caruso, G.; Leonardi, M.; Maimone, G.; Monticelli, L.S.; Azzaro, M.; Cuttitta, A.; Patti, B.; La Ferla, R. Seasonal changes on microbial metabolism and biomass in the euphotic layer of Sicilian Channel. Mar. Environ. Res. 2015, 112, 20–32. [Google Scholar] [CrossRef]
- Teixeira, M.C.; Santana, N.F.; Rodrigues de Azevedo, J.C.; Pagioro, T.A. Bacterioplankton features and its relations with doc characteristics and other limnological variables in Paraná river floodplain environments (PR/MS-Brazil). Braz. J. Microbiol. 2011, 42, 897–908. [Google Scholar] [CrossRef] [Green Version]
- Sigee, D.C. Freshwater Microbiology: Biodiversity and Dynamic Interactions of Microorganisms in the Freshwater Environment; John Wiley and Sons Ltd.: London, UK, 2005; ISBN 978-0-470-01125-6. [Google Scholar]
- Zubkov, M.V.; Sleigh, M.A.; Burkill, P.H.; Leakey, R.J.G. Bacterial growth and grazing in contrasting areas of north and south Atlantic. J. Plankton. Res. 2000, 22, 685–711. [Google Scholar] [CrossRef]
- Quiroga, M.V.; Mataloni, G.; Wanderley, B.M.S.; Amado, A.M.; Unrein, F. Bacterioplankton morphotypes structure and cytometric fingerprint rely on environmental conditions in a sub-Antarctic peatland. Hydrobiologia 2017, 787, 255–268. [Google Scholar] [CrossRef]
- Thingstad, T.F.; Øvreås, L.; Egge, J.K.; Løvdal, T.; Heldal, M. Use of non-limiting substrates to increase size; a generic strategy to simultaneously optimize uptake and minimize predation in pelagic osmotrophs? Ecol. Lett. 2005, 8, 675–682. [Google Scholar] [CrossRef]
- Racy, F.; Godinho, M.J.L.; Regali-Seleghim, M.H.; Bossolan, N.R.S.; Ferrari, A.C.; Lucca, J.V. Assessment of the applicability of morphological and size diversity indices to bacterial populations of reservoir in different trophic states. Acta. Limnol. Bras. 2005, 17, 395–408. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cruise | Station | Longitude | Latitude | CTD | Microbiological | Dissolved |
|---|---|---|---|---|---|---|
| [East] | [North] | Parameters | Nutrients | |||
| BANSIC-2012 | 22 | 15.235 | 36.638 | x | x | x |
| 143 | 15.315 | 36.364 | x | x | x | |
| 137 | 14.886 | 36.565 | x | x | x | |
| 188 | 14.624 | 36.607 | x | x | x | |
| 302 | 14.967 | 36.292 | x | x | x | |
| 641 | 14.700 | 35.945 | x | x | x | |
| NOVESAR-2013 | 22 | 15.046 | 36.647 | x | x | x |
| 137 | 15.001 | 36.589 | x | x | x | |
| 188 | 14.696 | 36.574 | x | x | x | |
| BANSIC-2013 | 2 | 15.259 | 36.787 | x | x | |
| 22 | 15.232 | 36.638 | x | x | ||
| 137 | 14.884 | 36.564 | x | x | ||
| 140 | 15.099 | 36.464 | x | |||
| 296 | 14.535 | 36.491 | x | x | ||
| 641 | 14.699 | 35.945 | x | |||
| 303 | 15.038 | 36.257 | x | x | ||
| 302 | 14.965 | 36.290 | x | |||
| 461 | 14.188 | 36.418 | x | x | x |
| BANSIC-12 | NOVESAR-13 | BANSIC-13 | ||
|---|---|---|---|---|
| T °C | range | 14.80–26.90 | 14.80–15.10 | 14.43–24.58 |
| mean ± sd | 18.20 ± 4.40 | 14.90 ± 0.08 | 19.65 ± 3.75 | |
| S | range | 37.3–38.70 | 38.0–38.80 | 37.56–38.71 |
| mean ± sd | 38.3 ± 0.30 | 38.4 ± 0.25 | 38.10 ± 0.42 | |
| DO (mg L−1) | range | 6.30–8.10 | 6.90–8.20 | 7.04–8.25 |
| mean ± sd | 7.30 ± 0.60 | 7.70 ± 0.38 | 7.64 ± 0.46 | |
| DEN (kg/m3) | range | 25.2–28.80 | 28.4–28.90 | 25.50–28.67 |
| mean ± sd | 27.70 ± 1.29 | 28.6 ± 0.18 | 27.08 ± 0.99 | |
| FLUO (µg L−1) | range | 0.02–0.39 | 0.04–0.38 | 0.01–8.25 |
| mean ± sd | 0.13 ± 0.11 | 0.24 ± 0.12 | 0.07 ± 0.09 | |
| NO3 (µmol L−1) | range | 0.13–1.62 | 0.15–3.12 | 0.03–2.43 |
| mean ± sd | 0.45 ± 0.37 | 1.26 ± 0.90 | 0.38 ± 0.62 | |
| PO4 (µmol L−1) | range | 0.001–0.070 | 0.023–0.127 | 0.02–0.02 |
| mean ± sd | 0.018 ± 0.02 | 0.057 ± 0.042 | 0.02 ± 0.00 | |
| SiO4 (µmol L−1) | range | 0.2–1.58 | 0.74–1.952 | 0.32–3.18 |
| mean ± sd | 0.85 ± 0.36 | 1.22 ± 0.37 | 0.81 ± 0.69 | |
| Total Chl a (mg m−3) | range | 0.024–0.317 | n.d. | 0.009–0.224 |
| mean ± sd | 0.099 ± 0.077 | n.d. | 0.055 ± 0.056 | |
| micro-sized (>10 μm) | range | 0.001–0.062 | n.d. | 0.001–0.056 |
| mean ± sd | 0.012 ± 0.015 | n.d. | 0.011 ± 0.014 | |
| nano-sized (10–2.0 μm) | range | 0.006–0.143 | n.d. | 0.003–0.071 |
| mean ± sd | 0.026 ± 0.028 | n.d. | 0.016 ± 0.019 | |
| pico-sized (2–0.2 μm) | range | 0.016–0.148 | n.d. | 0.004–0.097 |
| mean ± sd | 0.065 ± 0.047 | n.d. | 0.029 ± 0.296 |
| L (µm) | W (µm) | ||||||
|---|---|---|---|---|---|---|---|
| Mean ± Sd | Min | Max | Mean ± Sd | Min | Max | ||
| BANSIC-2012 | Total cells | 1.16 ± 0.79 | 0.32 | 6.34 | 0.47 ± 0.14 | 0.21 | 1.38 |
| n = 4573 | vibrios | 2.52 ± 0.57 | 1.46 | 4.06 | 0.40 ± 0.11 | 0.21 | 0.64 |
| spirillae | 2.90 ± 0.69 | 1.39 | 4.54 | 0.35 ± 0.09 | 0.21 | 0.64 | |
| coccobacilli | 0.85 ± 0.19 | 0.53 | 1.68 | 0.58 ± 0.12 | 0.32 | 1.05 | |
| Cocci | 0.51 ± 0.15 | 0.32 | 1.38 | 0.51 ± 0.15 | 0.32 | 1.38 | |
| rods | 1.68 ± 0.66 | 0.64 | 6.34 | 0.41 ± 0.10 | 0.21 | 0.76 | |
| curved rods | 1.99 ± 0.55 | 1.07 | 5.08 | 0.38 ± 0.12 | 0.21 | 0.81 | |
| filamentous forms | n.d | n.d | n.d | n.d | n.d | n.d | |
| NOVESAR-2013 | Total cells | 0.69 ± 0.42 | 0.32 | 3.88 | 0.37 ± 0.09 | 0.15 | 0.95 |
| n = 1344 | vibrios | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| spirillae | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |
| coccobacilli | 0.62 ± 0.17 | 0.42 | 1.7 | 0.42 ± 0.08 | 0.32 | 0.95 | |
| Cocci | 0.40 ± 0.09 | 0.32 | 0.95 | 0.40 ± 0.09 | 0.32 | 0.95 | |
| rods | 1.01 ± 0.36 | 0.42 | 3.23 | 0.32 ± 0.08 | 0.15 | 0.65 | |
| curved rods | 1.38 ± 0.33 | 0.74 | 2.24 | 0.26 ± 0.07 | 0.15 | 0.44 | |
| filamentous forms | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |
| BANSIC-2013 | Total cells | 0.85 ± 0.59 | 0.28 | 6.37 | 0.43 ± 0.14 | 0.11 | 1.83 |
| n = 2852 | vibrios | 2.23 ± 0.75 | 1.14 | 6.37 | 0.36 ± 0.11 | 0.15 | 0.65 |
| spirillae | 2.35 ± 0.73 | 0.71 | 3.94 | 0.34 ± 0.09 | 0.15 | 0.53 | |
| coccobacilli | 0.69 ± 0.18 | 0.28 | 2.19 | 0.46 ± 0.14 | 0.21 | 1.83 | |
| cocci | 0.48 ± 0.14 | 0.30 | 1.27 | 0.48 ± 0.14 | 0.30 | 1.27 | |
| rods | 1.17 ± 0.43 | 0.55 | 2.86 | 0.37 ± 0.11 | 0.11 | 0.65 | |
| curved rods | 1.56 ± 0.50 | 0.53 | 3.59 | 0.35 ± 0.10 | 0.53 | 3.59 | |
| filamentous forms | 3.37 ± 1.54 | 2.04 | 6.41 | 0.24 ± 0.012 | 0.21 | 0.24 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maimone, G.; Azzaro, M.; Placenti, F.; Paranhos, R.; Cabral, A.S.; Decembrini, F.; Zaccone, R.; Cosenza, A.; Rappazzo, A.C.; Patti, B.; et al. A Morphometric Approach to Understand Prokaryoplankton: A Study in the Sicily Channel (Central Mediterranean Sea). Microorganisms 2023, 11, 1019. https://doi.org/10.3390/microorganisms11041019
Maimone G, Azzaro M, Placenti F, Paranhos R, Cabral AS, Decembrini F, Zaccone R, Cosenza A, Rappazzo AC, Patti B, et al. A Morphometric Approach to Understand Prokaryoplankton: A Study in the Sicily Channel (Central Mediterranean Sea). Microorganisms. 2023; 11(4):1019. https://doi.org/10.3390/microorganisms11041019
Chicago/Turabian StyleMaimone, Giovanna, Maurizio Azzaro, Francesco Placenti, Rodolfo Paranhos, Anderson Sousa Cabral, Franco Decembrini, Renata Zaccone, Alessandro Cosenza, Alessandro Ciro Rappazzo, Bernardo Patti, and et al. 2023. "A Morphometric Approach to Understand Prokaryoplankton: A Study in the Sicily Channel (Central Mediterranean Sea)" Microorganisms 11, no. 4: 1019. https://doi.org/10.3390/microorganisms11041019