
Impact of Gut Microbiota on Host Aggression: Potential Applications for Therapeutic Interventions Early in Development
 ,
,
Abstract
:1. Introduction
2. Animal Models of Aggression
3. Intestinal Microbiota
3.1. Characteristics of the Intestinal Microbiota
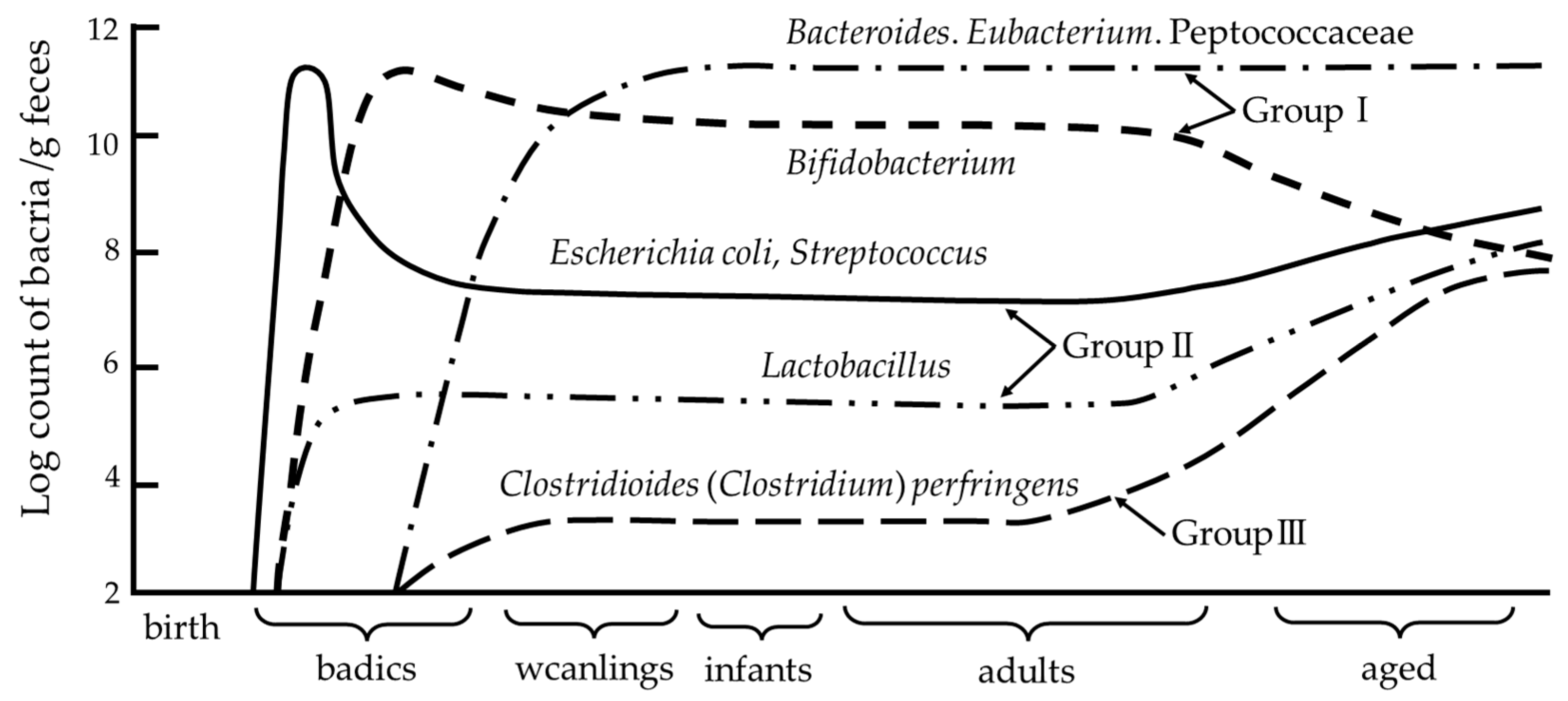
3.2. Developmental Processes of the Intestinal Microbiota
3.3. Role of Bifidobacteria
3.4. Effects of the Intestinal Environment on Behavior and Mental Activity
4. Intestinal Environment and Aggression in Animals
4.1. Mice
4.2. Dogs
4.3. Hamsters
4.4. Drosophila
5. Therapeutic Intervention for Aggression Targeting Gut Microbiota
5.1. Normalized Gut Microbiota in GF Mice
5.2. Probiotics and Prebiotics
5.3. Fecal Microbiota Transplant
5.4. Potential Applications for Therapeutic Interventions Early in Development
5.5. Maternal Gut Microbiota
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Anderson, D.J. Optogenetics, sex, and violence in the brain: Implications for psychiatry. Biol. Psychiatry 2012, 71, 1081–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Boer, S.F. Animal models of excessive aggression: Implications for human aggression and violence. Curr. Opin. Psychol. 2018, 19, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Flanigan, M.E.; Russo, S.J. Recent advances in the study of aggression. Neuropsychopharmacology 2019, 44, 241–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siever, L.J. Neurobiology of aggression and violence. Am. J. Psychiatry 2008, 165, 429–442. [Google Scholar] [CrossRef]
- Hsu, Y.; Earley, R.L.; Wolf, L.L. Modulation of aggressive behaviour by fighting experience: Mechanisms and contest outcomes. Biol. Rev. Camb. Philos. Soc. 2006, 81, 33–74. [Google Scholar] [CrossRef]
- Aleyasin, H.; Flanigan, M.E.; Russo, S.J. Neurocircuitry of aggression and aggression seeking behavior: Nose poking into brain circuitry controlling aggression. Curr. Opin. Neurobiol. 2018, 49, 184–191. [Google Scholar] [CrossRef]
- Mahadevia, D.; Saha, R.; Manganaro, A.; Chuhma, N.; Ziolkowski-Blake, A.; Morgan, A.A.; Dumitriu, D.; Rayport, S.; Ansorge, M.S. Dopamine promotes aggression in mice via ventral tegmental area to lateral septum projections. Nat. Commun. 2021, 12, 6796. [Google Scholar] [CrossRef]
- Caramaschi, D.; de Boer, S.F.; de Vries, H.; Koolhaas, J.M. Development of violence in mice through repeated victory along with changes in prefrontal cortex neurochemistry. Behav. Brain Res. 2008, 189, 263–272. [Google Scholar] [CrossRef] [Green Version]
- De Boer, S.F.; Caramaschi, D.; Natarajan, D.; Koolhaas, J.M. The vicious cycle towards violence: Focus on the negative feedback mechanisms of brain serotonin neurotransmission. Front. Behav. Neurosci. 2009, 3, 52. [Google Scholar] [CrossRef] [Green Version]
- Calcagnoli, F.; de Boer, S.F.; Beiderbeck, D.I.; Althaus, M.; Koolhaas, J.M.; Neumann, I.D. Local oxytocin expression and oxytocin receptor binding in the male rat brain is associated with aggressiveness. Behav. Brain Res. 2014, 261, 315–322. [Google Scholar] [CrossRef]
- Koolhaas, J.M.; de Boer, S.F.; Coppens, C.M.; Buwalda, B. Neuroendocrinology of coping styles: Towards understanding the biology of individual variation. Front. Neuroendocrinol. 2010, 31, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Haller, J.; Mikics, E.; Halász, J.; Tóth, M. Mechanisms differentiating normal from abnormal aggression: Glucocorticoids and serotonin. Eur. J. Pharmacol. 2005, 526, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Heinz, A.J.; Beck, A.; Meyer-Lindenberg, A.; Sterzer, P.; Heinz, A. Cognitive and neurobiological mechanisms of alcohol-related aggression. Nat. Rev. Neurosci. 2011, 12, 400–413. [Google Scholar] [CrossRef] [PubMed]
- Miczek, K.A.; Weerts, E.M.; Tornatzky, W.; DeBold, J.F.; Vatne, T.M. Alcohol and “bursts” of aggressive behavior: Ethological analysis of individual differences in rats. Psychopharmacology 1992, 107, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Chiavegatto, S.; Quadros, I.M.; Ambar, G.; Miczek, K.A. Individual vulnerability to escalated aggressive behavior by a low dose of alcohol: Decreased serotonin receptor mRNA in the prefrontal cortex of male mice. Genes Brain Behav. 2010, 9, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Veenema, A.H. Early life stress, the development of aggression and neuroendocrine and neurobiological correlates: What can we learn from animal models? Front. Neuroendocrinol. 2009, 30, 497–518. [Google Scholar] [CrossRef]
- Haller, J.; Harold, G.; Sandi, C.; Neumann, I.D. Effects of adverse early-life events on aggression and anti-social behaviours in animals and humans. J. Neuroendocrinol. 2014, 26, 724–738. [Google Scholar] [CrossRef]
- Tóth, M.; Halász, J.; Mikics, E.; Barsy, B.; Haller, J. Early social deprivation induces disturbed social communication and violent aggression in adulthood. Behav. Neurosci. 2008, 122, 849–854. [Google Scholar] [CrossRef]
- Mikami, K.; Tochio, T.; Watanabe, N. Modeling aggression in animals. In Handbook of Anger, Aggression, and Violence; Martin, C., Preedy, V.R., Patel, V.B., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 1–20. [Google Scholar]
- Wallace, T.C.; Guarner, F.; Madsen, K.; Cabana, M.D.; Gibson, G.; Hentges, E.; Sanders, M.E. Human gut microbiota and its relationship to health and disease. Nutr. Rev. 2011, 69, 392–403. [Google Scholar] [CrossRef]
- Guarner, F.; Malagelada, J.R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Mikami, K.; Kimura, M.; Takahashi, H. Influence of maternal bifidobacteria on the development of gut bifidobacteria in infants. Pharmaceuticals 2012, 5, 629–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faith, J.J.; Guruge, J.L.; Charbonneau, M.; Subramanian, S.; Seedorf, H.; Goodman, A.L.; Clemente, J.C.; Knight, R.; Heath, A.C.; Leibel, R.L.; et al. The long-term stability of the human gut microbiota. Science 2013, 341, 1237439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favier, C.F.; de Vos, W.M.; Akkermans, A.D. Development of bacterial and bifidobacterial communities in feces of newborn babies. Anaerobe 2003, 9, 219–229. [Google Scholar] [CrossRef]
- Bezirtzoglou, E. The intestinal microflora during the first weeks of life. Anaerobe 1997, 3, 173–177. [Google Scholar] [CrossRef]
- Mitsuoka, T. Intestinal flora and aging. Nutr. Rev. 1992, 50, 438–446. [Google Scholar] [CrossRef]
- Mackie, R.I.; Sghir, A.; Gaskins, H.R. Developmental microbial ecology of the neonatal gastrointestinal tract. Am. J. Clin. Nutr. 1999, 69, 1035S–1045S. [Google Scholar] [CrossRef] [Green Version]
- Caicedo, R.A.; Schanler, R.J.; Li, N.; Neu, J. The developing intestinal ecosystem: Implications for the neonate. Pediatr. Res. 2005, 58, 625–628. [Google Scholar] [CrossRef] [Green Version]
- Benno, Y.; Sawada, K.; Mitsuoka, T. The intestinal microflora of infants: Composition of fecal flora in breast-fed and bottle-fed infants. Microbiol. Immunol. 1984, 28, 975–986. [Google Scholar] [CrossRef]
- Lundell, A.C.; Björnsson, V.; Ljung, A.; Ceder, M.; Johansen, S.; Lindhagen, G.; Törnhage, C.J.; Adlerberth, I.; Wold, A.E.; Rudin, A. Infant B cell memory differentiation and early gut bacterial colonization. J. Immunol. 2012, 188, 4315–4322. [Google Scholar] [CrossRef] [Green Version]
- Olszak, T.; An, D.; Zeissig, S.; Vera, M.P.; Richter, J.; Franke, A.; Glickman, J.N.; Siebert, R.; Baron, R.M.; Kasper, D.L.; et al. Microbial exposure during early life has persistent effects on natural killer T cell function. Science 2012, 336, 489–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and butyrate-producing colon bacteria: Importance and strategies for their stimulation in the human gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudo, N.; Chida, Y.; Aiba, Y.; Sonoda, J.; Oyama, N.; Yu, X.N.; Kubo, C.; Koga, Y. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J. Physiol. 2004, 558, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Nishino, R.; Mikami, K.; Takahashi, H.; Tomonaga, S.; Furuse, M.; Hiramoto, T.; Aiba, Y.; Koga, Y.; Sudo, N. Commensal microbiota modulate murine behaviors in a strictly contamination-free environment confirmed by culture-based methods. Neurogastroenterol. Motil. 2013, 25, 521–528. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Leroy, F. Cross-feeding between bifidobacteria and butyrate-producing colon bacteria explains bifdobacterial competitiveness, butyrate production, and gas production. Int. J. Food Microbiol. 2011, 149, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.M.; Bercik, P. The relationship between intestinal microbiota and the central nervous system in normal gastrointestinal function and disease. Gastroenterology 2009, 136, 2003–2014. [Google Scholar] [CrossRef] [Green Version]
- Bercik, P.; Denou, E.; Collins, J.; Jackson, W.; Lu, J.; Jury, J.; Deng, Y.; Blennerhassett, P.; Macri, J.; McCoy, K.D.; et al. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology 2011, 141, 599–609.e3. [Google Scholar] [CrossRef] [Green Version]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The microbiota-gut-brain axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Neufeld, K.M.; Kang, N.; Bienenstock, J.; Foster, J.A. Reduced anxiety-like behavior and central neurochemical change in germ-free mice. Neurogastroenterol. Motil. 2011, 23, 255–264.e119. [Google Scholar] [CrossRef]
- Vuong, H.E.; Yano, J.M.; Fung, T.C.; Hsiao, E.Y. The microbiome and host behavior. Annu. Rev. Neurosci. 2017, 40, 21–49. [Google Scholar] [CrossRef]
- Leclercq, S.; Mian, F.M.; Stanisz, A.M.; Bindels, L.B.; Cambier, E.; Ben-Amram, H.; Koren, O.; Forsythe, P.; Bienenstock, J. Low-dose penicillin in early life induces long-term changes in murine gut microbiota, brain cytokines and behavior. Nat. Commun. 2017, 8, 15062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, V.; Han, M.H.; Graham, D.L.; Berton, O.; Renthal, W.; Russo, S.J.; Laplant, Q.; Graham, A.; Lutter, M.; Lagace, D.C.; et al. Molecular adaptations underlying susceptibility and resistance to social defeat in brain reward regions. Cell 2007, 131, 391–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golden, S.A.; Christoffel, D.J.; Heshmati, M.; Hodes, G.E.; Magida, J.; Davis, K.; Cahill, M.E.; Dias, C.; Ribeiro, E.; Ables, J.L.; et al. Epigenetic regulation of RAC1 induces synaptic remodeling in stress disorders and depression. Nat. Med. 2013, 19, 337–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, N.; Mikami, K.; Hata, T.; Kimoto, K.; Nishino, R.; Akama, F.; Yamamoto, K.; Sudo, N.; Koga, Y.; Matsumoto, H. Effect of gut microbiota early in life on aggressive behavior in mice. Neurosci. Res. 2021, 168, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Schneider, R.; Hoffmann, H.J.; Schicknick, H.; Moutier, R. Genetic analysis of isolation-induced aggression. I. Comparison between closely related inbred mouse strains. Behav. Neural Biol. 1992, 57, 198–204. [Google Scholar] [CrossRef]
- Kirchoff, N.S.; Udell, M.A.R.; Sharpton, T.J. The gut microbiome correlates with conspecific aggression in a small population of rescued dogs (Canis familiaris). PeerJ 2019, 7, e6103. [Google Scholar] [CrossRef] [Green Version]
- Mondo, E.; Barone, M.; Soverini, M.; D’Amico, F.; Cocchi, M.; Petrulli, C.; Mattioli, M.; Marliani, G.; Candela, M.; Accorsi, P.A. Gut microbiome structure and adrenocortical activity in dogs with aggressive and phobic behavioral disorders. Heliyon 2020, 6, e03311. [Google Scholar] [CrossRef]
- Gaiani, R.; Chiesa, F.; Mattioli, M.; Nannetti, G.; Galeati, G. Androstenedione and testosterone concentrations in plasma and milk of the cow throughout pregnancy. J. Reprod. Fertil. 1984, 70, 55–59. [Google Scholar] [CrossRef] [Green Version]
- Sylvia, K.E.; Jewell, C.P.; Rendon, N.M.; St John, E.A.; Demas, G.E. Sex-specific modulation of the gut microbiome and behavior in Siberian hamsters. Brain Behav. Immun. 2017, 60, 51–62. [Google Scholar] [CrossRef]
- Jasnow, A.M.; Huhman, K.L.; Bartness, T.J.; Demas, G.E. Short-day increases in aggression are inversely related to circulating testosterone concentrations in male Siberian hamsters (Phodopus sungorus). Horm. Behav. 2000, 38, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Rendon, N.M.; Rudolph, L.M.; Sengelaub, D.R.; Demas, G.E. The agonistic adrenal: Melatonin elicits female aggression via regulation of adrenal androgens. Proc. Biol. Sci. 2015, 282, 20152080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rendon, N.M.; Soini, H.A.; Scotti, M.A.; Weigel, E.R.; Novotny, M.V.; Demas, G.E. Photoperiod and aggression induce changes in ventral gland compounds exclusively in male Siberian hamsters. Horm. Behav. 2016, 81, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ren, C.C.; Sylvia, K.E.; Munley, K.M.; Deyoe, J.E.; Henderson, S.G.; Vu, M.P.; Demas, G.E. Photoperiod modulates the gut microbiome and aggressive behavior in Siberian hamsters. J. Exp. Biol. 2020, 223, jeb212548. [Google Scholar] [CrossRef]
- Cusick, J.A.; Wellman, C.L.; Demas, G.E. Maternal stress and the maternal microbiome have sex-specific effects on offspring development and aggressive behavior in Siberian hamsters (Phodopus sungorus). Horm. Behav. 2022, 141, 105146. [Google Scholar] [CrossRef]
- Shor, E.K.; Brown, S.P.; Freeman, D.A. Bacteria and bellicosity: Photoperiodic shifts in gut microbiota drive seasonal aggressive behavior in male Siberian hamsters. J. Biol. Rhythm. 2022, 37, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Albers, H.E.; Huhman, K.L.; Meisel, R.L. Hormonal basis of social conflict and communication. In Hormones, Brain and Behavior; Pfaff, D.W., Arnold, A.P., Fahrbach, S.E., Etgen, A.M., Rubin, R.T., Eds.; Academic Press: San Diego, CA, USA, 2002. [Google Scholar]
- Jia, Y.; Jin, S.; Hu, K.; Geng, L.; Han, C.; Kang, R.; Pang, Y.; Ling, E.; Tan, E.K.; Pan, Y.; et al. Gut microbiome modulates Drosophila aggression through octopamine signaling. Nat. Commun. 2021, 12, 2698. [Google Scholar] [CrossRef] [PubMed]
- Grinberg, M.; Levin, R.; Neuman, H.; Ziv, O.; Turjeman, S.; Gamliel, G.; Nosenko, R.; Koren, O. Antibiotics increase aggression behavior and aggression-related pheromones and receptors in Drosophila melanogaster. iScience 2022, 25, 104371. [Google Scholar] [CrossRef]
- Wang, L.; Anderson, D.J. Identification of an aggression-promoting pheromone and its receptor neurons in Drosophila. Nature 2010, 463, 227–231. [Google Scholar] [CrossRef] [Green Version]
- Schlüter, T.; Winz, O.; Henkel, K.; Prinz, S.; Rademacher, L.; Schmaljohann, J.; Dautzenberg, K.; Cumming, P.; Kumakura, Y.; Rex, S.; et al. The impact of dopamine on aggression: An [18F]-FDOPA PET Study in healthy males. J. Neurosci. 2013, 33, 16889–16896. [Google Scholar] [CrossRef] [Green Version]
- Langmajerová, M.; Roubalová, R.; Šebela, A.; Vevera, J. The effect of microbiome composition on impulsive and violent behavior: A systematic review. Behav. Brain Res. 2023, 440, 114266. [Google Scholar] [CrossRef]
- Tcherni-Buzzeo, M. Dietary interventions, the gut microbiome, and aggressive behavior: Review of research evidence and potential next steps. Aggress. Behav. 2023, 49, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104 (Suppl. S2), S1–S63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluznick, J.L. Gut microbiota in renal physiology: Focus on short-chain fatty acids and their receptors. Kidney Int. 2016, 90, 1191–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canani, R.B.; Costanzo, M.D.; Leone, L.; Pedata, M.; Meli, R.; Calignano, A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J. Gastroenterol. 2011, 17, 1519–1528. [Google Scholar] [CrossRef]
- Rivière, A.; Gagnon, M.; Weckx, S.; Roy, D.; De Vuyst, L. Mutual cross-feeding interactions between Bifidobacterium longum subsp. longum NCC2705 and Eubacterium rectale ATCC 33656 explain the bifidogenic and butyrogenic effects of arabinoxylan oligosaccharides. Appl. Environ. Microbiol. 2015, 81, 7767–7781. [Google Scholar] [CrossRef] [Green Version]
- Boets, E.; Gomand, S.V.; Deroover, L.; Preston, T.; Vermeulen, K.; De Preter, V.; Hamer, H.M.; Van den Mooter, G.; De Vuyst, L.; Courtin, C.M.; et al. Systemic availability and metabolism of colonic-derived short-chain fatty acids in healthy subjects: A stable isotope study. J. Physiol. 2017, 595, 541–555. [Google Scholar] [CrossRef] [Green Version]
- Burokas, A.; Arboleya, S.; Moloney, R.D.; Peterson, V.L.; Murphy, K.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Targeting the Microbiota-Gut-Brain Axis: Prebiotics Have Anxiolytic and Antidepressant-like Effects and Reverse the Impact of Chronic Stress in Mice. Biol. Psychiatry 2017, 82, 472–487. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Publishing: Washington, DC, USA, 2013. [Google Scholar]
- Fung, L.K.; Mahajan, R.; Nozzolillo, A.; Bernal, P.; Krasner, A.; Jo, B.; Coury, D.; Whitaker, A.; Veenstra-Vanderweele, J.; Hardan, A.Y. Pharmacologic treatment of severe irritability and problem behaviors in autism: A systematic review and meta-analysis. Pediatrics 2016, 137 (Suppl. S2), S124–S135. [Google Scholar] [CrossRef] [Green Version]
- Lecavalier, L. Behavioral and emotional problems in young people with pervasive developmental disorders: Relative prevalence, effects of subject characteristics, and empirical classification. J. Autism Dev. Disord. 2006, 36, 1101–1114. [Google Scholar] [CrossRef] [PubMed]
- Nikolov, R.N.; Bearss, K.E.; Lettinga, J.; Erickson, C.; Rodowski, M.; Aman, M.G.; McCracken, J.T.; McDougle, C.J.; Tierney, E.; Vitiello, B.; et al. Gastrointestinal symptoms in a sample of children with pervasive developmental disorders. J. Autism Dev. Disord. 2009, 39, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Orsso, C.E.; Deehan, E.C.; Kung, J.Y.; Tun, H.M.; Wine, E.; Madsen, K.L.; Zwaigenbaum, L.; Haqq, A.M. Probiotics, prebiotics, Synbiotics, and fecal microbiota transplantation in the treatment of behavioral symptoms of autism spectrum disorder: A systematic review. Autism Res. 2021, 14, 1820–1836. [Google Scholar] [CrossRef] [PubMed]
- Aman, M.G.; Singh, N.N.; Stewart, A.W.; Field, C.J. The aberrant behavior checklist: A behavior rating scale for the assessment of treatment effects. Am. J. Ment. Defic. 1985, 89, 485–491. [Google Scholar]
- Shaaban, S.Y.; El Gendy, Y.G.; Mehanna, N.S.; El-Senousy, W.M.; El-Feki, H.S.A.; Saad, K.; El-Asheer, O.M. The role of probiotics in children with autism spectrum disorder: A prospective, open-label study. Nutr. Neurosci. 2018, 21, 676–681. [Google Scholar] [CrossRef]
- Arnold, L.E.; Luna, R.A.; Williams, K.; Chan, J.; Parker, R.A.; Wu, Q.; Hollway, J.A.; Jeffs, A.; Lu, F.; Coury, D.L.; et al. Probiotics for gastrointestinal symptoms and quality of life in autism: A placebo-controlled pilot trial. J. Child Adolesc. Psychopharmacol. 2019, 29, 659–669. [Google Scholar] [CrossRef]
- Liu, Y.W.; Liong, M.T.; Chung, Y.E.; Huang, H.Y.; Peng, W.S.; Cheng, Y.F.; Lin, Y.S.; Wu, Y.Y.; Tsai, Y.C. Effects of Lactobacillus plantarum PS128 on children with autism spectrum disorder in Taiwan: A randomized, double-blind, placebo-controlled trial. Nutrients 2019, 11, 820. [Google Scholar] [CrossRef] [Green Version]
- Niu, M.; Li, Q.; Zhang, J.; Wen, F.; Dang, W.; Duan, G.; Li, H.; Ruan, W.; Yang, P.; Guan, C.; et al. Characterization of intestinal microbiota and probiotics treatment in children with autism spectrum disorders in China. Front. Neurol. 2019, 10, 1084. [Google Scholar] [CrossRef] [Green Version]
- Santocchi, E.; Guiducci, L.; Prosperi, M.; Calderoni, S.; Gaggini, M.; Apicella, F.; Tancredi, R.; Billeci, L.; Mastromarino, P.; Grossi, E.; et al. Effects of probiotic supplementation on gastrointestinal, sensory and core symptoms in autism spectrum disorders: A randomized controlled trial. Front. Psychiatry 2020, 11, 550593. [Google Scholar] [CrossRef]
- Sanctuary, M.R.; Kain, J.N.; Chen, S.Y.; Kalanetra, K.; Lemay, D.G.; Rose, D.R.; Yang, H.T.; Tancredi, D.J.; German, J.B.; Slupsky, C.M.; et al. Pilot study of probiotic/colostrum supplementation on gut function in children with autism and gastrointestinal symptoms. PLoS ONE 2019, 14, e0210064. [Google Scholar] [CrossRef] [Green Version]
- Van Nood, E.; Vrieze, A.; Nieuwdorp, M.; Fuentes, S.; Zoetendal, E.G.; de Vos, W.M.; Visser, C.E.; Kuijper, E.J.; Bartelsman, J.F.; Tijssen, J.G.; et al. Duodenal infusion of donor feces for recurrent Clostridium difficile. N. Engl. J. Med. 2013, 368, 407–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, D.W.; Adams, J.B.; Gregory, A.C.; Borody, T.; Chittick, L.; Fasano, A.; Khoruts, A.; Geis, E.; Maldonado, J.; McDonough-Means, S.; et al. Microbiota Transfer Therapy alters gut ecosystem and improves gastrointestinal and autism symptoms: An open-label study. Microbiome 2017, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.W.; Adams, J.B.; Coleman, D.M.; Pollard, E.L.; Maldonado, J.; McDonough-Means, S.; Caporaso, J.G.; Krajmalnik-Brown, R. Long-term benefit of microbiota Transfer Therapy on autism symptoms and gut microbiota. Sci. Rep. 2019, 9, 5821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuong, H.E.; Pronovost, G.N.; Williams, D.W.; Coley, E.J.L.; Siegler, E.L.; Qiu, A.; Kazantsev, M.; Wilson, C.J.; Rendon, T.; Hsiao, E.Y. The maternal microbiome modulates fetal neurodevelopment in mice. Nature 2020, 586, 281–286. [Google Scholar] [CrossRef]
- Mikami, K.; Takahashi, H.; Kimura, M.; Isozaki, M.; Izuchi, K.; Shibata, R.; Sudo, N.; Matsumoto, H.; Koga, Y. Influence of maternal bifidobacteria on the establishment of bifidobacteria colonizing the gut in infants. Pediatr. Res. 2009, 65, 669–674. [Google Scholar] [CrossRef] [Green Version]
- Sirilun, S.; Takahashi, H.; Boonyaritichaikij, S.; Chaiyasut, C.; Lertruangpanya, P.; Koga, Y.; Mikami, K. Impact of maternal bifidobacteria and the mode of delivery on Bifidobacterium microbiota in infants. Benef. Microbes 2015, 6, 767–774. [Google Scholar] [CrossRef]
- Takahashi, H.; Mikami, K.; Nishino, R.; Matsuoka, T.; Kimura, M.; Koga, Y. Comparative analysis of the properties of bifidobacterial isolates from fecal samples of mother–infant pairs. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 653–660. [Google Scholar] [CrossRef]
- Makino, H.; Kushiro, A.; Ishikawa, E.; Muylaert, D.; Kubota, H.; Sakai, T.; Oishi, K.; Martin, R.; Ben Amor, K.; Oozeer, R.; et al. Transmission of intestinal Bifidobacterium longum subsp. longum strains from mother to infant, determined by multilocus sequencing typing and amplified fragment length polymorphism. Appl. Environ. Microbiol. 2011, 77, 6788–6793. [Google Scholar] [CrossRef] [Green Version]
- Makino, H.; Kushiro, A.; Ishikawa, E.; Kubota, H.; Gawad, A.; Sakai, T.; Oishi, K.; Martin, R.; Ben-Amor, K.; Knol, J.; et al. Mother-to-infant transmission of intestinal bifidobacterial strains has an impact on the early development of vaginally delivered infant’s microbiota. PLoS ONE 2013, 8, e78331. [Google Scholar] [CrossRef] [Green Version]
- Makino, H. Bifidobacterial strains in the intestines of newborns originate from their mothers. Biosci. Microbiota Food Health 2018, 37, 79–85. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Reference | Animal Model | Main Comparable or Exposure | Assessment of Aggression | Effect of Gut Microbiota on Aggression |
|---|---|---|---|---|
| Leclercq et al. [42] | Mouse | Pregnant females were treated with either drinking water (control group), antibiotics, or antibiotics and Lactobacillus until weaning of pups (postnatal day 21). | Short (acute) version adapted from the chronic social defeat described by Krishnan et al. [43] and Golden et al. [44] was assessed. | Mice pups, whose gut microbiotas were disturbed with antibiotics from gestation through weaning, showed increased anxiety-like and impaired social behaviors, as well as a tendency to exhibit aggression. |
| Watanabe et al. [45] | Mouse | Germ-free (GF) and Ex-germ-free (EX-GF) mice | Aggressive behaviors including biting, wrestling, tail-rattling, aggressive grooming, or chasing were assessed based on the study by Schneider et al. [46] | Ex-GF mice with commensal gut microbiota showed significantly lower levels of aggression-related behavior than GF mice. When GF mice were conventionalized by administering feces from Ex-GF mice, the groups administered feces at 0 and 6 weeks of age showed aggression behaviors less frequently than normal GF mice. Among these mice conventionalized with Ex-GF feces, both the aggressive behavior rates of mice at 0 and 6 weeks were significantly lower than at 10 weeks. |
| Kirchoff et al. [47] | Dog | Aggressive and nonaggressive dogs | Aggressive dogs displayed aggression during one of three scenarios: an introduction to a life-size dog plush, introduction to a dog of the same-sex behind a barrier, and introduction to a dog of the same-sex without a barrier. Aggressive displays toward the life-size dog plush included growling, snarling, biting, holding, and shaking combined with tense behavior inconsistent with object play, as well as aggressive displays toward the same-sex dogs including growling and lunging, lunging and snarling, climbing on withers and growling, attempting to bite, and biting. | Differences in the β diversity of the gut microbiota between aggressive and nonaggressive dogs supported a link between canine aggression and the composition of the gut microbiota. In addition, several bacteria (Lactobacillus, Dorea, Blautia, Turicibacter, and Bacteroides) had increased and decreased relative abundances in aggressive dogs compared to nonaggressive dogs. |
| Mondo et al. [48] | Dog | Dogs with aggressive, phobic, or normal behavior | Aggressive behaviors were evaluated based on the study by Giussani et al. [49]. | The relative abundance of commonly classified subdominant bacteria, such as Catenibacterium and Megamonas, was increased in the gut microbiota of dogs exhibiting aggressive behavior. Levels of testosterone and cortisol, hormones involved in aggressive behavior, were not closely associated with gut microbiota. |
| Sylvia et al. [50] | Hamster | Hamsters that received antibiotics or sterilized water | Using the resident–intruder paradigm with same-sex social partners per previously outlined methods for this species, aggressive behaviors were assessed based on the studies by Jasnow et al. [51], Rendon et al. [52], and Rendon et al. [53]. | The effects of single versus repeated antibiotic treatments (including a recovery phase) on behavior were tested. Two treatments caused marked decreases in aggressive behavior in males; aggression returned to normal levels following recovery. Antibiotic-treated females, in contrast, showed decreased aggression after a single treatment. Unlike males, female aggression did not return to normal during either recovery period. |
| Ren et al. [54] | Hamster | Hamsters housed in either long-day or short-day (SD) photoperiods for 9 weeks. SD conditions were divided into short-day responders (SD-R) and short-day nonresponders (SD-NR) according to physiological response to changes in the photoperiod. | Aggressive behaviors including latency to first attack, frequency and duration of attacks, and chases were assessed based on the studies by Jasnow et al. [51], Rendon et al. [52], Rendon et al. [53], and Sylvia et al. [50]. | SD-R females displayed increased aggression. The relative abundance of anaeroplasmataceae in females was associated with aggression in SD-R hamsters. |
| Cusick et al. [55] | Hamster | Pregnant females were assigned to one of four treatments: antibiotics only, stress only, antibiotics and stress, or control. | The resident–intruder trial, the frequency and duration of actions performed by the experimental individuals (i.e., residents), was examined and scored, following an established protocol in the authors’ lab. Aggressive (e.g., attack, chase) and nonaggressive (e.g., intruder investigation) behaviors were assessed. | Considering males and females individually, female offspring produced by stress only mothers were more aggressive than both female offspring produced by control mothers and female offspring produced by antibiotic and stress mothers. Maternal exposure to antibiotics affected the aggressive behaviors of male offspring. |
| Shor et al. [56] | Hamster | Hamsters were randomly assigned into four treatment groups: LDfs, hamsters housed in long-day (LD) conditions, which received fecal microbiota transplants (FMTs) from a short-day (SD) (fs) donor; SDfl, hamsters housed in SD conditions that received FMT from an LD (fl) donor; LDfl, hamsters housed in LD conditions that received FMT from an LD (fl) donor; SDfs, hamsters housed in SD conditions that received FMT from an SD (fs) donor. | The resident–intruder (R–I) paradigm was conducted to quantify the effects of FMT on aggressive behavior. The R–I procedure involved placing an “intruder” animal into the home cage of a “resident” animal and observing the subsequent displays of aggressive behaviors [57]. | Implanting short-day Siberian hamsters with fecal microbiota from LD hamsters resulted in a reversal of seasonal aggression, whereby SD hamsters displayed aggression levels typical of LD hamsters. |
| Jia et al. [58] | Drosophila | Conventionally reared and germ-free (GF) flies, and GF embryos with mixed bacteria | Number of lunges in males and head butting in females was manually counted. For intermale aggression, lunging frequency and latency of fighting (the time from the beginning to the first lunging) were used to compare aggression levels. | The microbiota promoted aggressive behaviors in both Drosophila males and females. GF males showed a substantial decrease in inter-male aggression, which was rescued using microbial recolonization. These germ-free males were not as competitive as wildtype males for mating with females, although they displayed regular levels of courtship behaviors. |
| Grinberg et al. [59] | Drosophila | Flies grown on media supplemented with a mixture of antibiotics (Abx) to eliminate gut bacteria, Lactobacillus brevis-monocolonized flies, Lactobacillus plantarum-monocolonized flies, or untreated flies (control). Each group consisted of eight males. | Under ideal conditions for aggression [60], the visible movements of lunging, boxing, chasing, or wing threats were counted. | The Abx treatment increased the number of aggressive encounters among male flies compared to the control group, whereas supplementation with L. plantarum or L. brevis reduced aggression compared to both the Abx-treated flies. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikami, K.; Watanabe, N.; Tochio, T.; Kimoto, K.; Akama, F.; Yamamoto, K. Impact of Gut Microbiota on Host Aggression: Potential Applications for Therapeutic Interventions Early in Development. Microorganisms 2023, 11, 1008. https://doi.org/10.3390/microorganisms11041008
Mikami K, Watanabe N, Tochio T, Kimoto K, Akama F, Yamamoto K. Impact of Gut Microbiota on Host Aggression: Potential Applications for Therapeutic Interventions Early in Development. Microorganisms. 2023; 11(4):1008. https://doi.org/10.3390/microorganisms11041008
Chicago/Turabian StyleMikami, Katsunaka, Natsuru Watanabe, Takumi Tochio, Keitaro Kimoto, Fumiaki Akama, and Kenji Yamamoto. 2023. "Impact of Gut Microbiota on Host Aggression: Potential Applications for Therapeutic Interventions Early in Development" Microorganisms 11, no. 4: 1008. https://doi.org/10.3390/microorganisms11041008