
Continuous Production of Ethanol, 1-Butanol and 1-Hexanol from CO with a Synthetic Co-Culture of Clostridia Applying a Cascade of Stirred-Tank Bioreactors

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Medium
2.2. Preculture Preparations
2.3. Stirred-Tank Bioreactor Set-Up for Gas-Fermentation Processes
2.4. Analytical Methods
3. Results and Discussion
3.1. CO-Inhibition Studies with C. kluyveri in Stirred-Tank Reactors
3.2. Continuous Sulfide Supply in Autotrophic Batch Processes with C. carboxidivorans
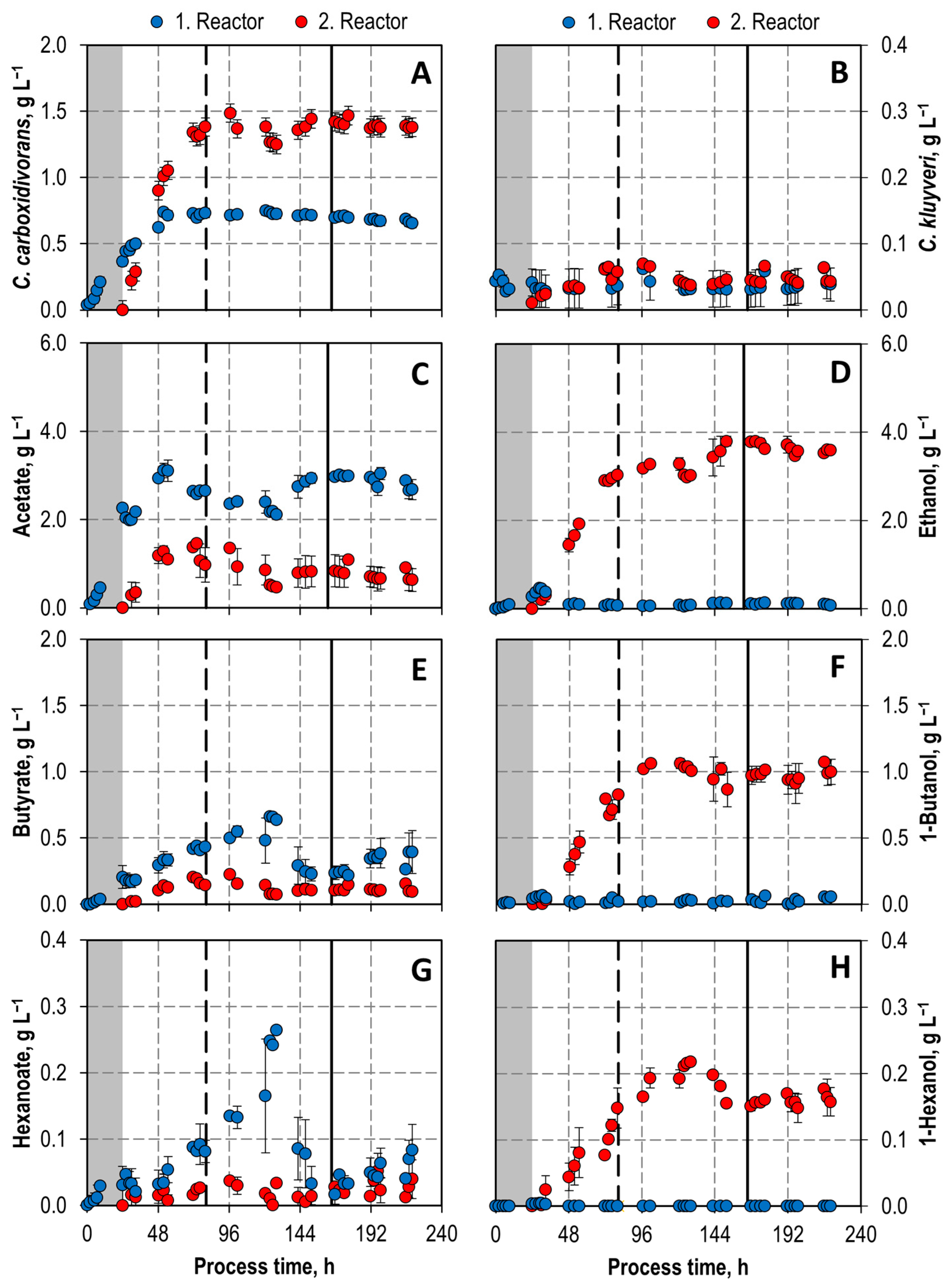
3.3. Co-Cultivation Studies in Two Bioreactors in Series with Varying pH and pCO
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kottenhahn, P.; Philipps, G.; Jennewein, S. Hexanol biosynthesis from syngas by Clostridium carboxidivorans P7–product toxicity, temperature dependence and in situ extraction. Heliyon 2021, 7, e07732. [Google Scholar] [CrossRef]
- Weuster-Botz, D. Process engineering aspects for the microbial conversion of C1 gases. Adv. Biochem. Eng. Biotechnol. 2022, 180, 33–56. [Google Scholar] [CrossRef]
- Daniell, J.; Köpke, M.; Simpson, S. Commercial biomass syngas fermentation. Energies 2012, 5, 5372–5417. [Google Scholar] [CrossRef] [Green Version]
- Köpke, M.; Mihalcea, C.; Liew, F.; Tizard, J.H.; Ali, M.S.; Conolly, J.J.; Al-Sinawi, B.; Simpson, S.D. 2,3-butanediol production by acetogenic bacteria, an alternative route to chemical synthesis, using industrial waste gas. Appl. Environ. Microbiol. 2011, 77, 5467–5475. [Google Scholar] [CrossRef] [Green Version]
- Molitor, B.; Richter, H.; Martin, M.E.; Jensen, R.O.; Juminaga, A.; Mihalcea, C.; Angenent, L.T. Carbon recovery by fermentation of CO-rich off gases–turning steel mills into biorefineries. Bioresour. Technol. 2016, 215, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.V.; Mohan, S.V.; Chang, Y.-C. Medium-Chain Fatty Acids (MCFA) production through anaerobic fermentation using Clostridium kluyveri: Effect of ethanol and acetate. Appl. Biochem. Biotechnol. 2018, 185, 594–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbusch, K.J.J.; Hamelers, H.V.M.; Plugge, C.M.; Buisman, C.J.N. Biological formation of caproate and caprylate from acetate: Fuel and chemical production from low grade biomass. Energy Environ. Sci. 2011, 4, 216–224. [Google Scholar] [CrossRef]
- Spirito, C.M.; Richter, H.; Rabaey, K.; Stams, A.J.M.; Angenent, L.T. Chain elongation in anaerobic reactor microbiomes to recover resources from waste. Curr. Opin. Biotechnol. 2014, 27, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Hinderberger, J.; Seedorf, H.; Zhang, J.; Buckel, W.; Thauer, R.K. Coupled ferredoxin and crotonyl coenzyme A (CoA) reduction with NADH catalyzed by the butyryl-CoA dehydrogenase/Etf complex from Clostridium kluyveri. J. Bacteriol. 2008, 190, 843–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalcante, W.d.A.; Leitão, R.C.; Gehring, T.A.; Angenent, L.T.; Santaella, S.T. Anaerobic fermentation for n-caproic acid production: A review. Process Biochem. 2017, 54, 106–119. [Google Scholar] [CrossRef]
- Gildemyn, S.; Molitor, B.; Usack, J.G.; Nguyen, M.; Rabaey, K.; Angenent, L.T. Upgrading syngas fermentation effluent using Clostridium kluyveri in a continuous fermentation. Biotechnol. Biofuels 2017, 10, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucek, L.A.; Spirito, C.M.; Angenent, L.T. High n-caprylate productivities and specificities from dilute ethanol and acetate: Chain elongation with microbiomes to upgrade products from syngas fermentation. Energy Environ. Sci. 2016, 9, 3482–3494. [Google Scholar] [CrossRef]
- Yin, Y.; Zhang, Y.; Karakashev, D.B.; Wang, J.; Angelidaki, I. Biological caproate production by Clostridium kluyveri from ethanol and acetate as carbon sources. Bioresour. Technol. 2017, 241, 638–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seedorf, H.; Fricke, W.F.; Veith, B.; Brüggemann, H.; Liesegang, H.; Strittmatter, A.; Miethke, M.; Buckel, W.; Hinderberger, J.; Li, F.; et al. The genome of Clostridium kluyveri, a strict anaerobe with unique metabolic features. Proc. Natl. Acad. Sci. USA 2008, 105, 2128–2133. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Yang, L.; Huo, S.; Su, Y.; Zhang, Y. Optimization of the cell immobilization-based chain-elongation process for efficient n -caproate production. ACS Sustain. Chem. Eng. 2021, 9, 4014–4023. [Google Scholar] [CrossRef]
- Diender, M.; Stams, A.J.M.; Sousa, D.Z. Production of medium-chain fatty acids and higher alcohols by a synthetic co-culture grown on carbon monoxide or syngas. Biotechnol. Biofuels 2016, 9, 82. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Blanco, C.; Veiga, M.C.; Kennes, C. Efficient production of n-caproate from syngas by a co-culture of Clostridium aceticum and Clostridium kluyveri. J. Environ. Manag. 2022, 302, 113992. [Google Scholar] [CrossRef]
- Bäumler, M.; Schneider, M.; Ehrenreich, A.; Liebl, W.; Weuster-Botz, D. Synthetic co-culture of autotrophic Clostridium carboxidivorans and chain elongating Clostridium kluyveri monitored by flow cytometry. Microb. Biotechnol. 2022, 15, 1471–1485. [Google Scholar] [CrossRef]
- Schneider, M.; Bäumler, M.; Lee, N.M.; Weuster-Botz, D.; Ehrenreich, A.; Liebl, W. Monitoring co-cultures of Clostridium carboxidivorans and Clostridium kluyveri by fluorescence in situ hybridization with specific 23S rRNA oligonucleotide probes. submitted. Syst. Appl. Microbiol. 2021, 44, 126271. [Google Scholar] [CrossRef]
- Doll, K.; Rückel, A.; Kämpf, P.; Wende, M.; Weuster-Botz, D. Two stirred-tank bioreactors in series enable continuous production of alcohols from carbon monoxide with Clostridium carboxidivorans. Bioprocess Biosyst. Eng. 2018, 41, 1403–1416. [Google Scholar] [CrossRef]
- Oliveira, L.; Röhrenbach, S.; Holzmüller, V.; Weuster-Botz, D. Continuous sulfide supply enhanced autotrophic production of alcohols with Clostridium ragsdalei. Bioresour. Bioprocess. 2022, 9, 15. [Google Scholar] [CrossRef]
- Tomlinson, N.; Barker, H.A. Carbon dioxide and acetate utilization by Clostridium kluyveri: I. Influence of nutritional conditions on utilization patterns. J. Biol. Chem. 1954, 585–595. [Google Scholar] [CrossRef]
- San-Valero, P.; Fernández-Naveira, Á.; Veiga, M.C.; Kennes, C. Influence of electron acceptors on hexanoic acid production by Clostridium kluyveri. J. Environ. Manag. 2019, 242, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Lemon, B.J.; Peters, J.W. Binding of exogenously added carbon monoxide at the active site of the iron-only hydrogenase (CpI) from Clostridium pasteurianum. Biochemistry 1999, 38, 12969–12973. [Google Scholar] [CrossRef]
- Do, Y.S.; Smeenk, J.; Broer, K.M.; Kisting, C.J.; Brown, R.; Heindel, T.J.; Bobik, T.A.; DiSpirito, A.A. Growth of Rhodospirillum rubrum on synthesis gas: Conversion of CO to H2 and poly-beta-hydroxyalkanoate. Biotechnol. Bioeng. 2007, 97, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Najafpour, G.D.; Younesi, H.; Lahijani, P.; Uzir, M.H.; Mohamed, A.R. Bioconversion of synthesis gas to second generation biofuels: A review. Renew. Sustain. Energy Rev. 2011, 15, 4255–4273. [Google Scholar] [CrossRef]
- Abubackar, H.N.; Veiga, M.C.; Kennes, C. Biological conversion of carbon monoxide: Rich syngas or waste gases to bioethanol. Biofuels Bioprod. Bioref. 2011, 5, 93–114. [Google Scholar] [CrossRef] [Green Version]
- Barker, H.A.; Kamen, M.D.; Bornstein, B.T. The synthesis of butyric and caproic acids from ethanol and acetic acid by Clostridium Kluyveri. Proc. Natl. Acad. Sci. USA 1945, 31, 373–381. [Google Scholar] [CrossRef] [Green Version]
- Angenent, L.T.; Richter, H.; Buckel, W.; Spirito, C.M.; Steinbusch, K.J.J.; Plugge, C.M.; Strik, D.P.B.T.B.; Grootscholten, T.I.M.; Buisman, C.J.N.; Hamelers, H.V.M. Chain elongation with reactor microbiomes: Open-culture biotechnology to produce biochemicals. Environ. Sci. Technol. 2016, 50, 2796–2810. [Google Scholar] [CrossRef]
- González-Cabaleiro, R.; Lema, J.M.; Rodríguez, J.; Kleerebezem, R. Linking thermodynamics and kinetics to assess pathway reversibility in anaerobic bioprocesses. Energy Environ. Sci. 2013, 6, 3780. [Google Scholar] [CrossRef]
- Richter, H.; Molitor, B.; Diender, M.; Sousa, D.Z.; Angenent, L.T. A narrow pH range supports butanol, hexanol, and octanol production from syngas in a continuous co-culture of Clostridium ljungdahlii and Clostridium kluyveri with in-line product extraction. Front. Microbiol. 2016, 7, 1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| css, g L−1 | cfinal, g L−1 | STY, g L−1 d−1 | |||||
|---|---|---|---|---|---|---|---|
| 1. Reactor | 2. Reactor | Batch [18] | 1. Reactor | 2. Reactor | Cascade | Batch [18] | |
| C. kluyveri | 0.04 ± 0.02 | 0.04 ± 0.01 | 0.06 ± 0.01 | 0.15 ± 0.05 | 0.03 ± 0.01 | 0.04 ± 0.02 | 0.03 ± 0.01 |
| C. carboxidivorans | 0.68 ± 0.01 | 1.42 ± 0.04 | 0.47 ± 0.03 | 1.78 ± 0.02 | 1.62 ± 0.16 | 1.24 ± 0.05 | 0.73 ± 0.02 |
| Acetate | 2.89 ± 0.09 | 0.84 ± 0.36 | 1.20 ± 0.02 | 6.62 ± 0.15 | −4.56 ± 0.55 | 0.75 ± 0.31 | 4.15 ± 0.03 |
| Ethanol | 0.11 ± 0.01 | 3.78 ± 0.00 | 2.09 ± 0.20 | 0.28 ± 0.02 | 8.18 ± 0.14 | 3.12 ± 0.05 | 0.59 ± 0.00 |
| Butyrate | 0.31 ± 0.07 | 0.11 ± 0.05 | 0.31 ± 0.05 | 1.08 ± 0.14 | −0.72 ± 0.25 | 0.11 ± 0.04 | 0.51 ± 0.03 |
| 1-Butanol | 0.03 ± 0.01 | 0.97 ± 0.07 | 0.84 ± 0.06 | 0.04 ± 0.01 | 2.38 ± 0.12 | 0.89 ± 0.06 | 0.33 ± 0.03 |
| Hexanoate | 0.05 ± 0.01 | 0.03 ± 0.01 | 0.36 ± 0.03 | 0.33 ± 0.05 | −0.24 ± 0.04 | 0.02 ± 0.01 | 0.12 ± 0.02 |
| 1-Hexanol | − | 0.16 ± 0.01 | 0.24 ± 0.01 | − | 0.44 ± 0.01 | 0.15 ± 0.01 | 0.08 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bäumler, M.; Burgmaier, V.; Herrmann, F.; Mentges, J.; Schneider, M.; Ehrenreich, A.; Liebl, W.; Weuster-Botz, D. Continuous Production of Ethanol, 1-Butanol and 1-Hexanol from CO with a Synthetic Co-Culture of Clostridia Applying a Cascade of Stirred-Tank Bioreactors. Microorganisms 2023, 11, 1003. https://doi.org/10.3390/microorganisms11041003
Bäumler M, Burgmaier V, Herrmann F, Mentges J, Schneider M, Ehrenreich A, Liebl W, Weuster-Botz D. Continuous Production of Ethanol, 1-Butanol and 1-Hexanol from CO with a Synthetic Co-Culture of Clostridia Applying a Cascade of Stirred-Tank Bioreactors. Microorganisms. 2023; 11(4):1003. https://doi.org/10.3390/microorganisms11041003
Chicago/Turabian StyleBäumler, Miriam, Veronika Burgmaier, Fabian Herrmann, Julian Mentges, Martina Schneider, Armin Ehrenreich, Wolfgang Liebl, and Dirk Weuster-Botz. 2023. "Continuous Production of Ethanol, 1-Butanol and 1-Hexanol from CO with a Synthetic Co-Culture of Clostridia Applying a Cascade of Stirred-Tank Bioreactors" Microorganisms 11, no. 4: 1003. https://doi.org/10.3390/microorganisms11041003