
Heterologous 1,3-Propanediol Production Using Different Recombinant Clostridium beijerinckii DSM 6423 Strains

Abstract
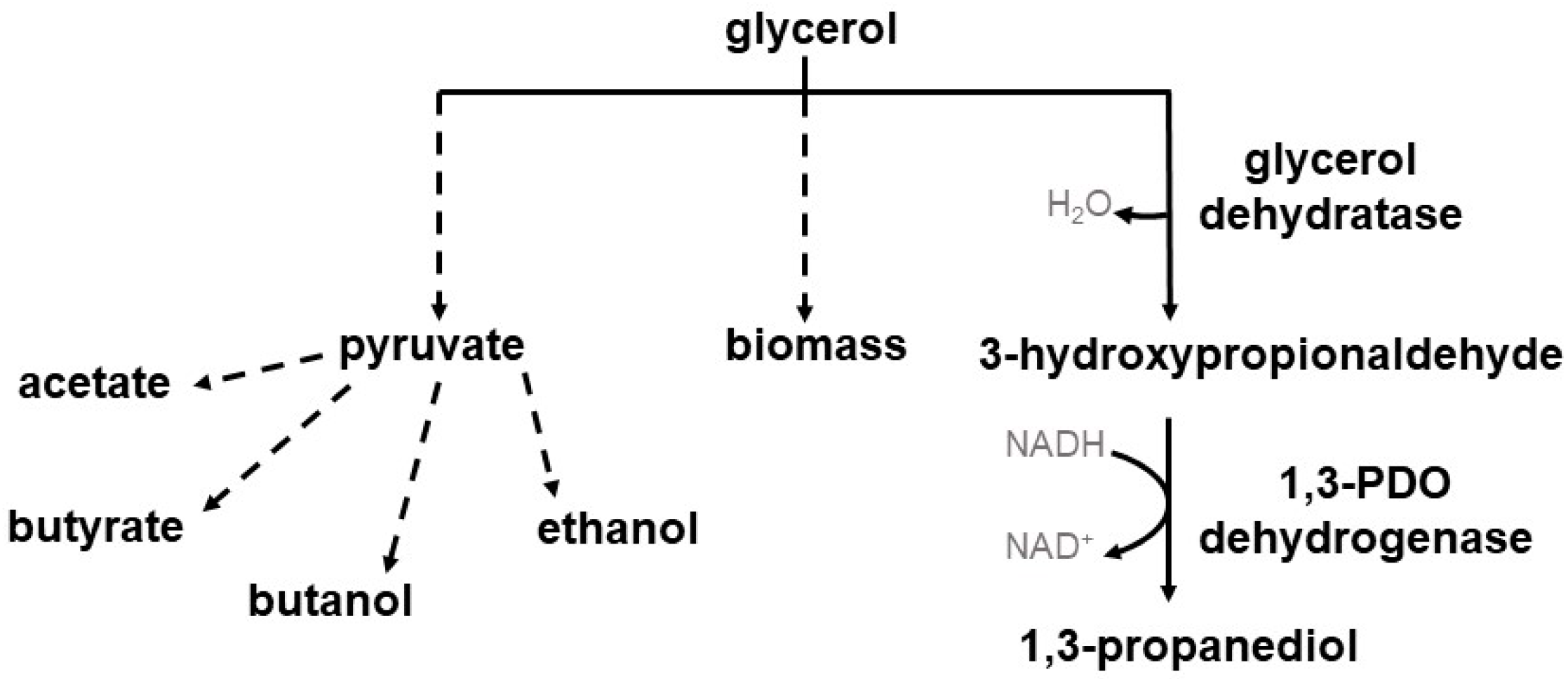
:1. Introduction
2. Material & Methods
2.1. Strains and Cultivation
2.2. Construction of Recombinant C. beijerinckii Strains
2.3. Strain Verification
2.4. Growth Conditions of Batch Experiments
2.5. Analytical Methods
3. Results
3.1. Construction of Recombinant 1,3-PDO Production C. beijerinckii Strains
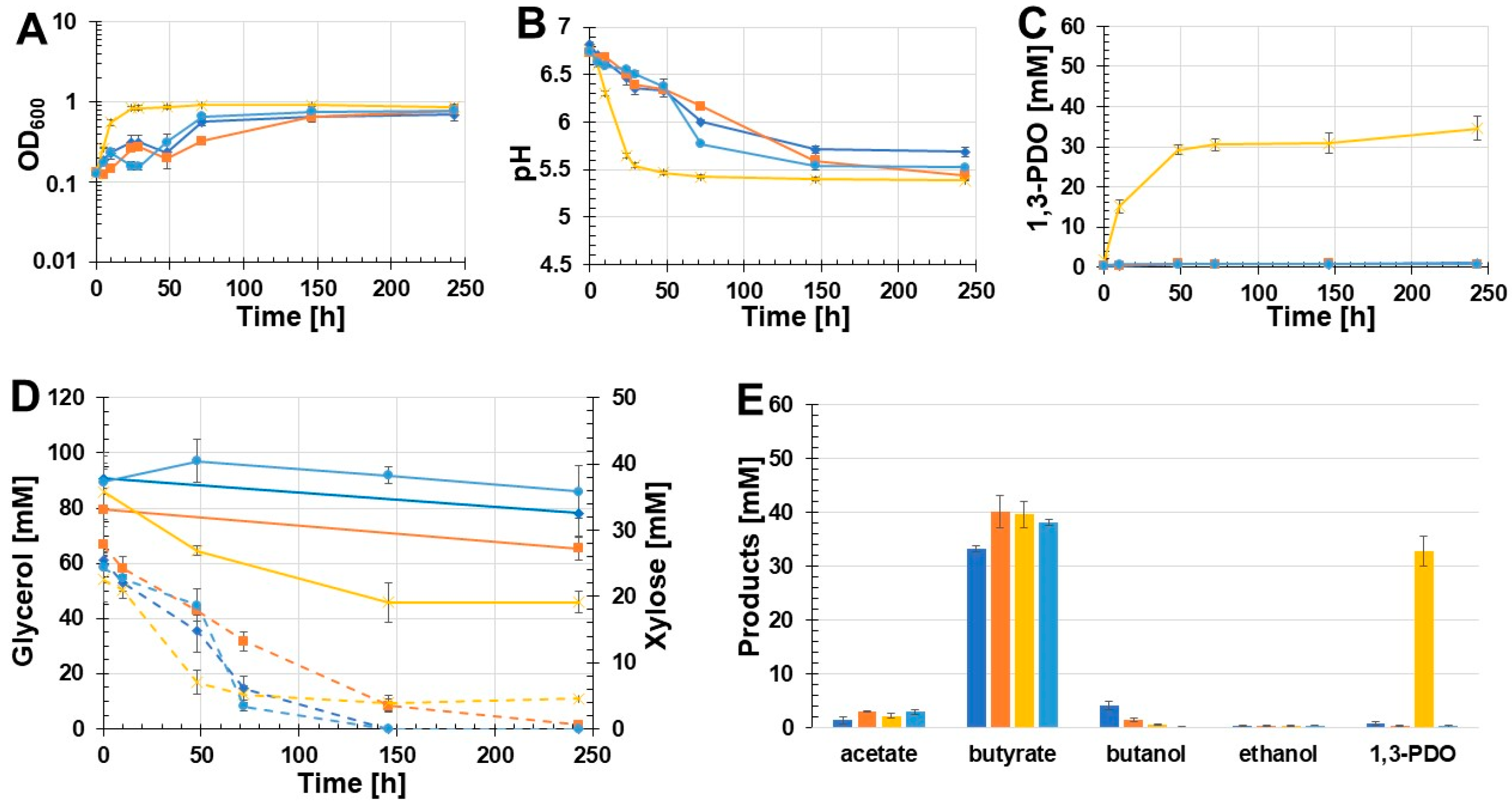
3.2. Recombinant 1,3-PDO Production
3.3. Influence of Buffered Glycerol Medium on 1,3-PDO Production
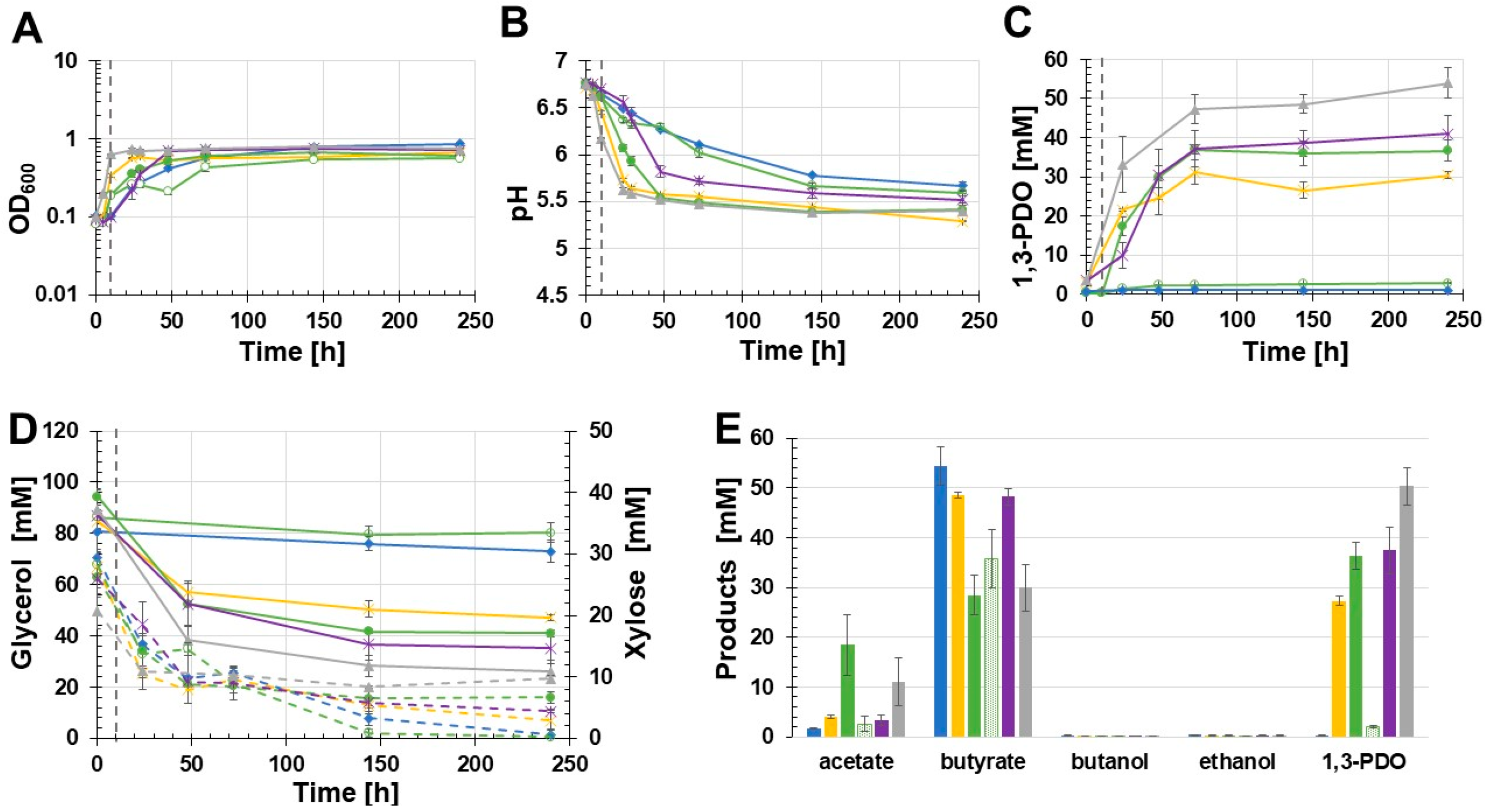
3.4. Effect of Vitamin B12 Supplementation
3.5. Effect of Different Promoters on Recombinant 1,3-PDO Production
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- da Silva Ruy, A.D.; de Brito Alves, R.M.; Hewer, T.L.R.; de Aguiar Pontes, D.; Teixeira, L.S.G.; Pontes, L.A.M. Catalysts for glycerol hydrogenolysis to 1,3-propanediol: A review of chemical routes and market. Catal. Today 2020, 381, 243–253. [Google Scholar] [CrossRef]
- Zhu, F.; Liu, D.; Chen, Z. Recent advances in biological production of 1,3-propanediol: New routes and engineering strategies. Green Chem. 2022, 24, 1390–1403. [Google Scholar] [CrossRef]
- Kaur, G.; Srivastava, A.K.; Chand, S. Advances in biotechnological production of 1,3-propanediol. Biochem. Eng. J. 2012, 64, 106–118. [Google Scholar] [CrossRef]
- Przystałowska, H.; Zeyland, J.; Szymanowska-Powałowska, D.; Szalata, M.; Słomski, R.; Lipiński, D. 1,3-Propanediol production by new recombinant Escherichia coli containing genes from pathogenic bacteria. Microbiol. Res. 2015, 171, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Xu, F.; Liu, H.; Liu, D. Downstream processing of 1,3-propanediol fermentation broth. J. Chem. Technol. Biotechnol. Int. Res. Process Environ. Clean Technol. 2006, 81, 102–108. [Google Scholar] [CrossRef]
- Cameron, D.C.; Altaras, N.E.; Hoffman, M.L.; Shaw, A.J. Metabolic engineering of propanediol pathways. Biotechnol. Prog. 1998, 14, 116–125. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Y.; Gao, H.; Wang, H.; Wan, Z.; Jiang, Y.; Xin, F.; Zhang, W.; Jiang, M. Current advances in microbial production of 1,3-propanediol. Biofuels Bioprod. Biorefining 2021, 15, 1566–1583. [Google Scholar] [CrossRef]
- Nakamura, C.E.; Whited, G.M. Metabolic engineering for the microbial production of 1,3-propanediol. Curr. Opin. Biotechnol. 2003, 14, 454–459. [Google Scholar] [CrossRef]
- Dürre, P. Technical alcohols and ketones. In Industrial Microbiology; Wilson, D.B., Sahm, H., Stahmann, K.P., Koffas, M., Eds.; Wiley-VCH: Weinheim, Germany, 2020; pp. 95–116. [Google Scholar]
- Biotechnology Innovation Organization. Advancing the biobased economy: Renewable chemical biorefinery commercialization, progress, and market opportunities, 2016 and beyond. Ind. Biotechnol. 2016, 12, 290–294. [Google Scholar] [CrossRef]
- Kurian, J.V. A new polymer platform for the future—Sorona® from corn derived 1,3-propanediol. J. Polym. Environ. 2005, 13, 159–167. [Google Scholar] [CrossRef]
- Clomburg, J.M.; Gonzalez, R. Anaerobic fermentation of glycerol: A platform for renewable fuels and chemicals. Trends Biotechnol. 2013, 31, 20–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazdani, S.S.; Gonzalez, R. Anaerobic fermentation of glycerol: A path to economic viability for the biofuels industry. Curr. Opin. Biotechnol. 2007, 18, 213–219. [Google Scholar] [CrossRef]
- Yang, F.; Hanna, M.A.; Sun, R. Value-added uses for crude glycerol—A byproduct of biodiesel production. Biotechnol. Biofuels 2012, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Anand, P.; Saxena, R.K. A comparative study of solvent-assisted pretreatment of biodiesel derived crude glycerol on growth and 1,3-propanediol production from Citrobacter freundii. New Biotechnol. 2012, 29, 199–205. [Google Scholar] [CrossRef]
- Hermann, B.G.; Patel, M. Today’s and tomorrow’s bio-based bulk chemicals from white biotechnology. Appl. Biochem. Biotechnol. 2007, 136, 361–388. [Google Scholar] [CrossRef] [PubMed]
- Biebl, H.; Zeng, A.P.; Menzel, K.; Deckwer, W.D. Fermentation of glycerol to 1,3-propanediol and 2,3-butanediol by Klebsiella pneumoniae. Appl. Microbiol. Biotechnol. 1998, 50, 24–29. [Google Scholar] [CrossRef]
- Barbirato, F.; Camarasa-Claret, C.; Grivet, J.P.; Bories, A. Glycerol fermentation by a new 1,3-propanediol-producing microorganism: Enterobacter agglomerans. Appl. Microbiol. Biotechnol. 1995, 43, 786–793. [Google Scholar] [CrossRef]
- Homann, T.; Tag, C.; Biebl, H.; Deckwer, W.D.; Schink, B. Fermentation of glycerol to 1,3-propanediol by Klebsiella and Citrobacter strains. Appl. Microbiol. Biotechnol. 1990, 33, 121–126. [Google Scholar] [CrossRef] [Green Version]
- Leja, K.; Czaczyk, K.; Myszka, K. Biotechnological synthesis of 1,3-propanediol using Clostridium ssp. Afr. J. Biotechnol. 2011, 10, 11093–11101. [Google Scholar]
- Kobayashi, H.; Tanizawa, Y.; Sakamoto, M.; Nakamura, Y.; Ohkuma, M.; Tohno, M. Reclassification of Clostridium diolis Biebl and Spröer 2003 as a later heterotypic synonym of Clostridium beijerinckii Donker 1926 (Approved Lists 1980) emend. Keis et al. 2001. Int. J. Syst. Evol. Microbiol. 2020, 70, 2463–2466. [Google Scholar] [CrossRef]
- Gungormusler, M.; Gonen, C.; Azbar, N. Continuous production of 1,3-propanediol using raw glycerol with immobilized Clostridium beijerinckii NRRL B-593 in comparison to suspended culture. Bioprocess Biosyst. Eng. 2011, 34, 727–733. [Google Scholar] [CrossRef]
- Biebl, H.; Spröer, C. Taxonomy of the glycerol fermenting clostridia and description of Clostridium diolis sp. nov. Syst. Appl. Microbiol. 2002, 25, 491–497. [Google Scholar] [CrossRef]
- Biebl, H. Fermentation of glycerol by Clostridium pasteurianum—Batch and continuous culture studies. J. Ind. Microbiol. Biotechnol. 2001, 27, 18–26. [Google Scholar] [CrossRef]
- Biebl, H.; Marten, S.; Hippe, H.; Deckwer, W.D. Glycerol conversion to 1,3-propanediol by newly isolated clostridia. Appl. Microbiol. Biotechnol. 1992, 36, 592–597. [Google Scholar] [CrossRef]
- Heyndrickx, M.; De Vos, P.; Vancanneyt, M.; De Ley, J. The fermentation of glycerol by Clostridium butyricum LMG 1212t2 and 1213t1 and C. pasteurianum LMG 3285. Appl. Microbiol. Biotechnol. 1991, 34, 637–642. [Google Scholar] [CrossRef]
- Luers, F.; Seyfried, M.; Daniel, R.; Gottschalk, G. Glycerol conversion to 1,3-propanediol by Clostridium pasteurianum: Cloning and expression of the gene encoding 1,3-propanediol dehydrogenase. FEMS Microbiol. Lett. 1997, 154, 337–345. [Google Scholar] [CrossRef]
- Sedlar, K.; Vasylkivska, M.; Musilova, J.; Branska, B.; Provaznik, I.; Patakova, P. Phenotypic and genomic analysis of isopropanol and 1,3-propanediol producer Clostridium diolis DSM 15410. Genomics 2021, 113, 1109–1119. [Google Scholar] [CrossRef]
- Raynaud, C.; Sarçabal, P.; Meynial-Salles, I.; Croux, C.; Soucaille, P. Molecular characterization of the 1,3-propanediol (1,3-PD) operon of Clostridium butyricum. Proc. Natl. Acad. Sci. USA 2003, 100, 5010–5015. [Google Scholar] [CrossRef] [Green Version]
- Celińska, E. Debottlenecking the 1,3-propanediol pathway by metabolic engineering. Biotechnol. Adv. 2010, 28, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Diallo, M.; Hocq, R.; Collas, F.; Chartier, G.; Wasels, F.; Wijaya, H.S.; Werten, M.W.T.; Wolbert, E.J.H.; Kengen, S.W.M.; von der Oost, J.; et al. Adaptation and application of a two-plasmid inducible CRISPR-Cas9 system in Clostridium beijerinckii. Methods 2020, 172, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Pyne, M.E.; Moo-Young, M.; Chung, D.A.; Chou, C.P. Development of an electrotransformation protocol for genetic manipulation of Clostridium pasteurianum. Biotechnol. Biofuels 2013, 6, 50. [Google Scholar] [CrossRef] [Green Version]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor: New York, NY, USA, 2012. [Google Scholar]
- Heap, J.T.; Pennington, O.J.; Cartman, S.T.; Minton, N.P. A modular system for Clostridium shuttle plasmids. J. Microbiol. Methods 2009, 78, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Flaiz, M.; Ludwig, G.; Bengelsdorf, F.R.; Dürre, P. Production of the biocommodities butanol and acetone from methanol with fluorescent FAST-tagged proteins using metabolically engineered strains of Eubacterium limosum. Biotechnol. Biofuels 2021, 14, 117. [Google Scholar] [CrossRef]
- Baur, T.; Wentzel, A.; Dürre, P. Production of propionate using metabolically engineered strains of Clostridium saccharoperbutylacetonicum. Appl. Microbiol. Biotechnol. 2022, 106, 7547–7562. [Google Scholar] [CrossRef]
- Gungormusler, M.; Gonen, C.; Azbar, N. 1,3-Propanediol production potential by a locally isolated strain of Klebsiella pneumoniae in comparison to Clostridium beijerinckii NRRL B593 from waste glycerol. J. Polym. Environ. 2011, 19, 812–817. [Google Scholar] [CrossRef]
- Macis, L.; Daniel, R.; Gottschalk, G. Properties and sequence of the coenzyme B12-dependent glycerol dehydratase of Clostridium pasteurianum. FEMS Microbiol. Lett. 1998, 164, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Seifert, C.; Bowien, S.; Gottschalk, G.; Daniel, R. Identification and expression of the genes and purification and characterization of the gene products involved in reactivation of coenzyme B12-dependent glycerol dehydratase of Citrobacter freundii. Eur. J. Biochem. 2001, 268, 2369–2378. [Google Scholar] [CrossRef] [PubMed]
- Mera, P.E.; Escalante-Semerena, J.C. Multiple roles of ATP: Cob (I) alamin adenosyltransferases in the conversion of B12 to coenzyme B12. Appl. Microbiol. Biotechnol. 2010, 88, 41–48. [Google Scholar] [CrossRef]
- Hoffmeister, S.; Gerdom, M.; Bengelsdorf, F.R.; Linder, S.; Flüchter, S.; Öztürk, H.; Blümke, W.; May, A.; Fischer, R.J.; Bahl, H.; et al. Acetone production with metabolically engineered strains of Acetobacterium woodii. Metab. Eng. 2016, 36, 37–47. [Google Scholar] [CrossRef]
- Tummala, S.B.; Welker, N.E.; Papoutsakis, E.T. Development and characterization of a gene expression reporter system for Clostridium acetobutylicum ATCC 824. Appl. Environ. Microbiol. 1999, 65, 3793–3799. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, T.; Nakayama, S.; Nakaya, K.; Yoshino, S.; Furukawa, K. Characterization of the sol operon in butanol-hyperproducing Clostridium saccharoperbutylacetonicum strain N1-4 and its degeneration mechanism. Biosci. Biotechnol. Biochem. 2007, 71, 58–68. [Google Scholar] [CrossRef] [Green Version]
- Hartman, A.H.; Liu, H.; Melville, S.B. Construction and characterization of a lactose-inducible promoter system for controlled gene expression in Clostridium perfringens. Appl. Environ. Microbiol. 2011, 77, 471–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gungormusler, M.; Gonen, C.; Azbar, N. Comparative evaluation of Clostridium beijerinckii (NRRL B-593) and Klebsiella pneumoniae for 1,3 propanediol production. J. Biotechnol. 2010, 150, 210–211. [Google Scholar] [CrossRef]
- Johnson, E.A.; Lin, E.C. Klebsiella pneumoniae 1,3-propanediol: NAD+ oxidoreductase. J. Bacteriol. 1987, 169, 2050–2054. [Google Scholar] [CrossRef] [Green Version]
- Nasir, A.; Ashok, S.; Shim, J.Y.; Park, S.; Yoo, T.H. Recent progress in the understanding and engineering of coenzyme B12-dependent glycerol dehydratase. Front. Bioeng. Biotechnol. 2020, 8, 500867. [Google Scholar] [CrossRef]
- Fokum, E.; Zabed, H.M.; Ravikumar, Y.; Elshobary, M.E.; Chandankere, R.; Zhang, Y.; Yun, J.; Qi, X. Co-fermentation of glycerol and sugars by Clostridium beijerinckii: Enhancing the biosynthesis of 1,3-propanediol. Food Biosci. 2021, 41, 101028. [Google Scholar] [CrossRef]
- Vivek, N.; Pandey, A.; Binod, P. Biological valorization of pure and crude glycerol into 1,3-propanediol using a novel isolate Lactobacillus brevis N1E9. 3.3. Bioresour. Technol. 2016, 213, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Moscoviz, R.; Trably, E.; Bernet, N. Consistent 1,3-propanediol production from glycerol in mixed culture fermentation over a wide range of pH. Biotechnol. Biofuels 2016, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Nariya, H.; Miyata, S.; Kuwahara, T.; Okabe, A. Development and characterization of a xylose-inducible gene expression system for Clostridium perfringens. Appl. Environ. Microbiol. 2011, 77, 8439–8441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poehlein, A.; Solano, J.D.M.; Flitsch, S.K.; Krabben, P.; Winzer, K.; Reid, S.J.; Jones, D.T.; Green, E.; Minton, N.P.; Daniel, R.; et al. Microbial solvent formation revisited by comparative genome analysis. Biotechnol. Biofuels 2017, 10, 58. [Google Scholar] [CrossRef] [Green Version]
- González-Pajuelo, M.; Meynial-Salles, I.; Mendes, F.; Andrade, J.C.; Vasconcelos, I.; Soucaille, P. Metabolic engineering of Clostridium acetobutylicum for the industrial production of 1,3-propanediol from glycerol. Metab. Eng. 2005, 7, 329–336. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Features | Reference |
|---|---|---|
| C. beijerinckii DSM 6423 | wild-type | DSMZ * GmbH, Brunswick, Germany |
| E. coli XL1-Blue MRF’ | Δ(mrcA)183, Δ(mrcCB-hsdSMR-mrr)173, endA1, supE44, thi-1, recA1, gyrA96, relA1, lac, [F’ proAB lacIq ZΔM15 Tn10 (TetR)] | Agilent Technologies, Santa Clara, CA, USA |
| E. coli SCS110 | rpsL, (StrR), thr, leu, endA, thi-1, lacy, galK, galT, ara, tonA, tsx, dam, dcm, supE44D, (lac-proAB), [F’ traD36 proAB lacIqZΔM15] | Agilent Technologies, Santa Clara, CA, USA |
| pMTL83251 | clostridial shuttle vector, EmR | [34] |
| pMTL83251_PthlA_FAST | pMTL83251 containing PthlA from C. acetobutylicum and feg (FAST-encoding gene) | [35] |
| pMTL83251_PbgaL_FAST | pMTL83251 containing PbgaL from C. perfringens and feg | [35] |
| pMTL83251_Pbld_FAST | pMTL83251 containing Pbld from C. saccharoperbutylacetonicum and feg | [36] |
| pMTL83251_Ppta-ack_1,3-PDO.CLOBI | pMTL83251 containing 1,3-PDO gene cluster from C. beijerinckii DSM 6423 and pta-ack promoter from C. ljungdahlii | This study |
| pMTL83251_Ppta-ack_1,3-PDO.diolis | pMTL83251 containing 1,3-PDO gene cluster from C. beijerinckii DSM 15410 and pta-ack promoter from C. ljungdahlii | This study |
| pMTL83251_PthlA_dhaBCET.Cpas | pMTL83251 containing dhaBCE and dhaT from C. pasteurianum DSM 525 and thlA promoter from C. acetobutylicum | This study |
| pMTL83251_PbgaL_1,3-PDO.diolis | pMTL83251 containing 1,3-PDO gene cluster from C. beijerinckii DSM 15410 and bgaL promoter from C. perfringens | This study |
| pMTL83251_PthlA_1,3-PDO.diolis | pMTL83251 containing 1,3-PDO gene cluster from C. beijerinckii DSM 15410 and thlA promoter from C. acetobutylicum | This study |
| pMTL83251_Pbld_1,3-PDO.diolis | pMTL83251 containing 1,3-PDO gene cluster from C. beijerinckii DSM 15410 and bld promoter from C. saccharoperbutylacetonicum | This study |
| Primer | Sequence | Purpose |
|---|---|---|
| dhaB1/2CoT.diol_fwd | ttaaatttaaagggaggactctagaatgataagtaaaggatttagtacc | amplification of 1,3-PDO gene cluster from C. beijerinckii DSM 15410 |
| dhaB1/2CoT.diol_rev | gcaggcttcttatttttatgctagcttaataagcagctttaaatatatttacg | |
| PthlA.Cpas_fwd | cgaattcgagctcggtacccgggtcaagaagaggcacctcatc | amplification of thlA promoter from C. acetobutylicum |
| PthlA.Cpas_rev | ccctcctggtcaccaaattttgatacggggtaacag | |
| dhaBCE_fwd | aaatttggtgaccaggaggggatcccatgaagtcaaaacgatttcaag | amplification of dhaBCE from C. pasteurianum DSM 525 |
| dhaBCE_rev | aattcctcctctagtcctctattctaactttatttc | |
| dhaT_fwd | agaggactagaggaggaattataaaatgagaatg | amplification of dhaT gene from C. pasteurianum DSM 525 |
| dhaT_rev | atggacgcgtgacgtcgactttaaaatgcttctctaaatattttaactatatc | |
| PbgaL_1,3-PDO_fwd | cgaattcgagctcggtacccgggtaatttagatattaattctaaattaagtgaaat | amplification of bgaL promoter from pMTL83251_PbgaL_FAST |
| PbgaL_1,3-PDO_rev | taaatcctttacttatcattctcgagaccctcccaatacatttaaaataa | |
| PthlA_1,3-PDO_fwd | cgaattcgagctcggtacccgggtcaagaagaggcacctcatc | amplification of thlA promoter from pMTL83251_PthlA_FAST |
| PthlA_1,3-PDO_rev | taaatcctttacttatcattctcgagacctcctaaattttgatacgg | |
| Pbld_1,3-PDO_fwd | cgaattcgagctcggtacccggggatatttcccccataagtaaag | amplification of bld promoter from pMTL83251_Pbld_FAST |
| Pbld_1,3-PDO_rev | taaatcctttacttatcattctcgagtcctccttatgatttaaaaattaataac | |
| 16S-27F | ataagcttggatccagagtttgatcctggctcag | amplification of 16S rRNA gene |
| 1492r | actcgaggatatcggttaccttgttacgactt |
| 1,3-PDO Producing C. beijerinckii Strain | Glycerol Consumption [mM] | 1,3-PDO Production [mM] | Yield [mol1,3-PDO/molglycerol | Improvement [%] 1 |
|---|---|---|---|---|
| C. beijerinckii [pMTL83251_Ppta-ack_1,3-PDO.diolis] (unbuffered glycerol medium) | 33.3 | 18.9 | 0.57 | --- |
| C. beijerinckii [pMTL83251_Ppta-ack_1,3-PDO.diolis] (MOPS buffered glycerol medium) | 48.3 2 | 34.4 2 | 0.71 2 | 82 2 |
| C. beijerinckii [pMTL83251_PbgaL_1,3-PDO.diolis] induced | 53.1 | 36.3 | 0.68 | 92 |
| C. beijerinckii [pMTL83251_Pbld_1,3-PDO.diolis] | 52.1 | 37.4 | 0.72 | 98 |
| C. beijerinckii [pMTL83251_PthlA_1,3-PDO.diolis] | 63.4 | 50.4 | 0.79 | 167 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schoch, T.; Baur, T.; Kunz, J.; Stöferle, S.; Dürre, P. Heterologous 1,3-Propanediol Production Using Different Recombinant Clostridium beijerinckii DSM 6423 Strains. Microorganisms 2023, 11, 784. https://doi.org/10.3390/microorganisms11030784
Schoch T, Baur T, Kunz J, Stöferle S, Dürre P. Heterologous 1,3-Propanediol Production Using Different Recombinant Clostridium beijerinckii DSM 6423 Strains. Microorganisms. 2023; 11(3):784. https://doi.org/10.3390/microorganisms11030784
Chicago/Turabian StyleSchoch, Teresa, Tina Baur, Johanna Kunz, Sophia Stöferle, and Peter Dürre. 2023. "Heterologous 1,3-Propanediol Production Using Different Recombinant Clostridium beijerinckii DSM 6423 Strains" Microorganisms 11, no. 3: 784. https://doi.org/10.3390/microorganisms11030784