
Evaluation of Biofilm Cultivation Models for Efficacy Testing of Disinfectants against Salmonella Typhimurium Biofilms
 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Disinfection and Neutralization Conditions
2.3. Definition of Successful Disinfection
2.4. Disinfectant Testing in Suspension Assay
2.5. Disinfectant Testing in Biofilm Assay
2.5.1. Glass Bead Assay (GBA)
2.5.2. PVC Coupon Assay (PCA)
2.5.3. Stainless Steel Coupon Assay (SSCA)
2.6. Determination of Repeatability and Reproducibility
2.7. Statistical Analysis
3. Results
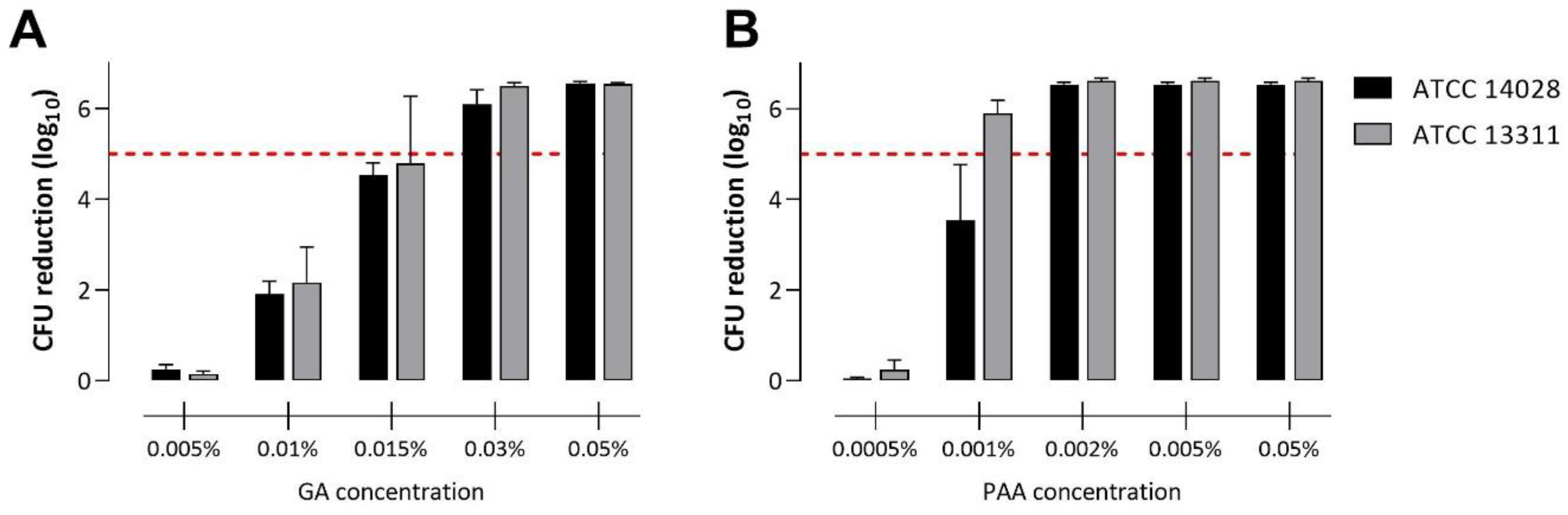
3.1. Disinfection of Planktonic Salmonella in Suspension
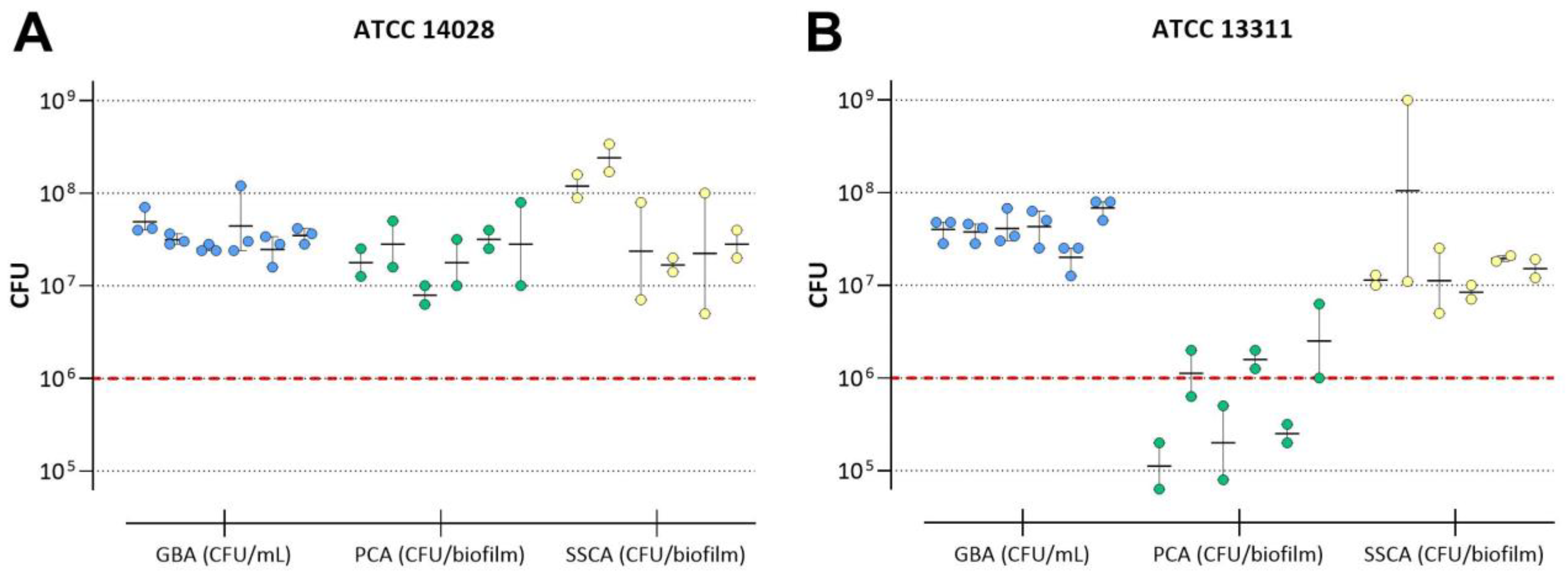
3.2. Cultivation of Salmonella Biofilms
3.2.1. Achievable CFU in Untreated Biofilms
3.2.2. Repeatability of CFU in Untreated Biofilms
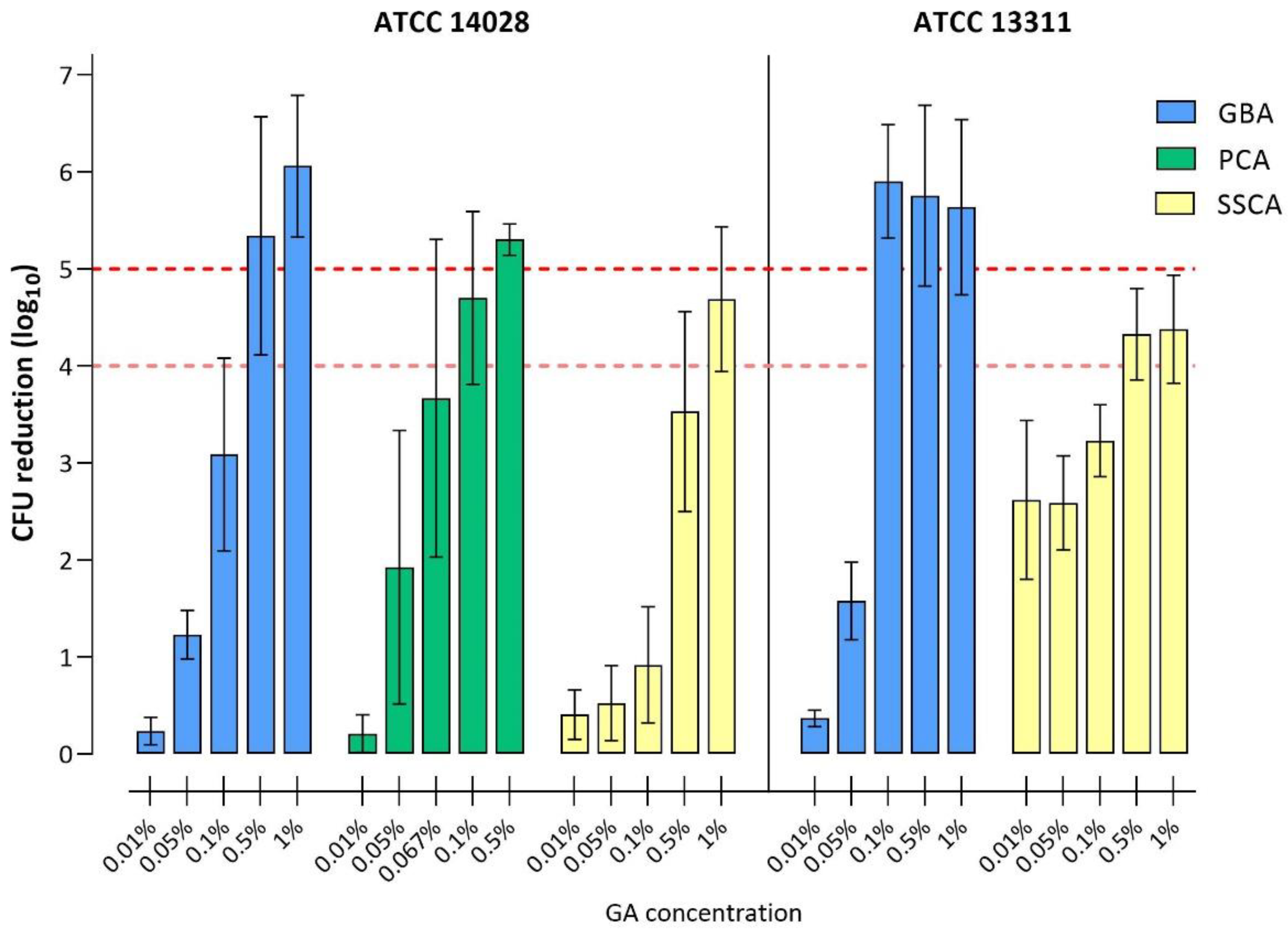
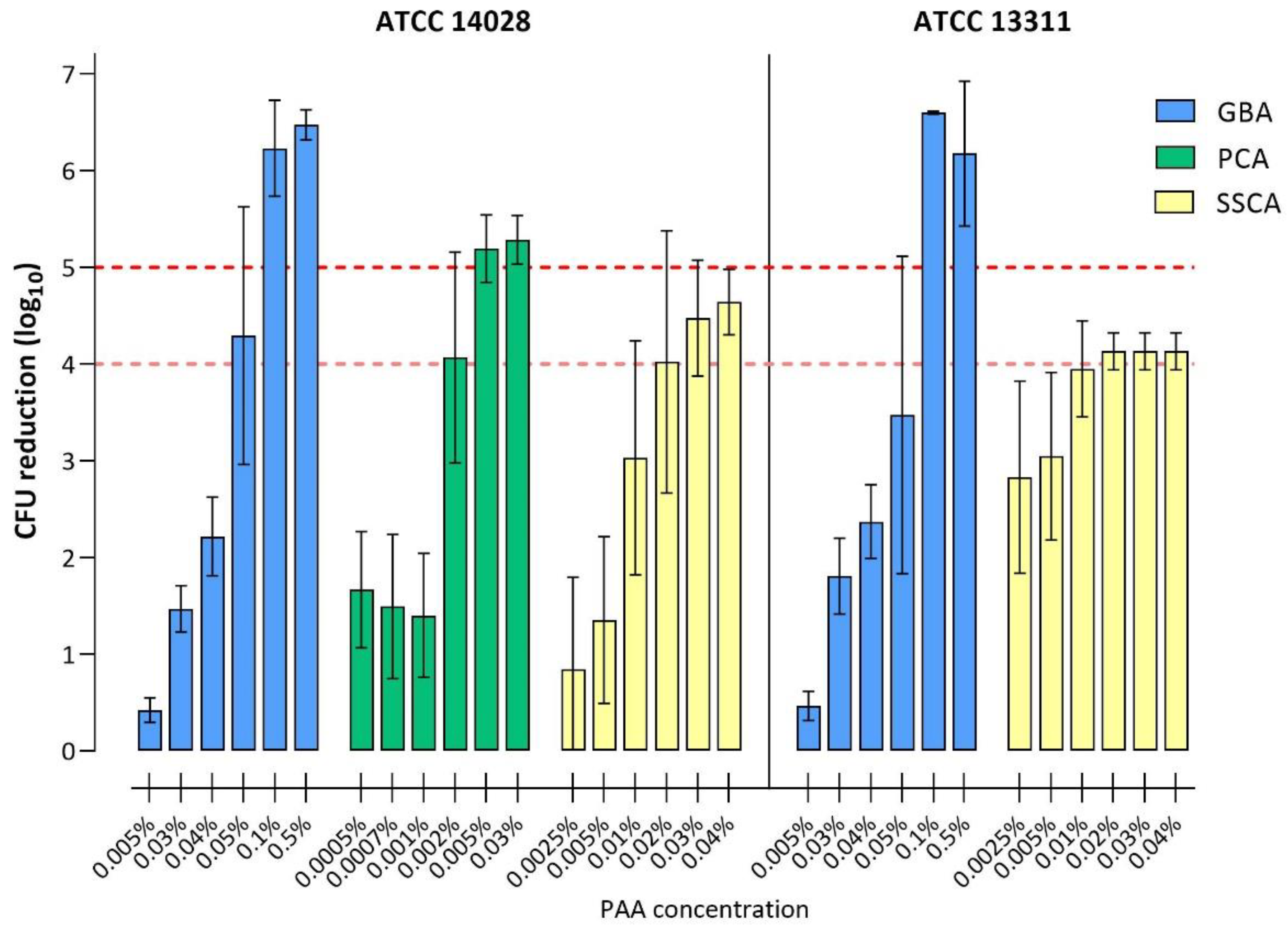
3.3. Disinfectant Efficacy Testing of Biofilms
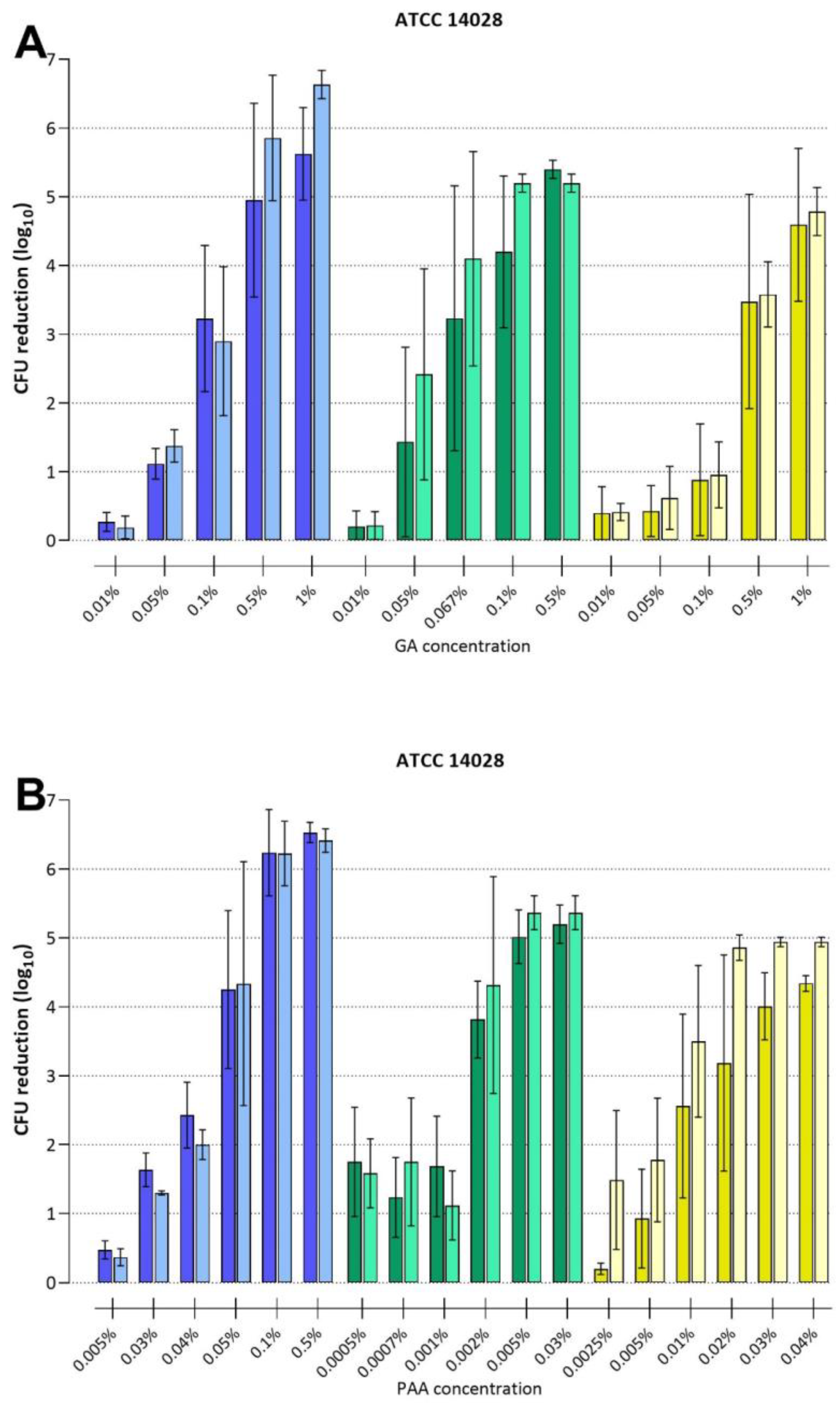
3.3.1. Disinfection with Glutaraldehyde
3.3.2. Disinfection with Peracetic Acid
3.4. Intra-Laboratory Reproducibility
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Centre for Disease Prevention and Control, European Food Safety Authority. Salmonellosis. In ECDC. Annual Epidemiological Report for 2017; ECDC: Stockholm, Sweden, 2020; Available online: https://www.ecdc.europa.eu/sites/default/files/documents/salmonellosis-annual-epidemiological-report-2017.pdf (accessed on 30 November 2022).
- ECDC. Surveillance Atlas of Infectious Diseases; ECDC: Stockholm, Sweden, 2022; Available online: https://atlas.ecdc.europa.eu/public/index.aspx?Dataset=27&HealthTopic=46 (accessed on 9 November 2022).
- Publications Office of the European Union. RASFF—The Rapid Alert System for Food and Feed—Annual Report 2020; Publications Office of the European Union: Luxembourg, 2021; Available online: https://food.ec.europa.eu/system/files/2021-08/rasff_pub_annual-report_2020.pdf (accessed on 30 November 2022).
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as complex differentiated communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef] [Green Version]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Ciofu, O.; Tolker-Nielsen, T. Tolerance and Resistance of Pseudomonas aeruginosa Biofilms to Antimicrobial Agents-How P. aeruginosa Can Escape Antibiotics. Front. Microbiol. 2019, 10, 913. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Bridier, A.; Briandet, R.; Thomas, V.; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: A review. Biofouling 2011, 27, 1017–1032. [Google Scholar] [CrossRef]
- Peng, J.S.; Tsai, W.C.; Chou, C.C. Inactivation and removal of Bacillus cereus by sanitizer and detergent. Int. J. Food Microbiol. 2002, 77, 11–18. [Google Scholar] [CrossRef]
- Zogaj, X.; Nimtz, M.; Rohde, M.; Bokranz, W.; Römling, U. The multicellular morphotypes of Salmonella typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Mol. Microbiol. 2001, 39, 1452–1463. [Google Scholar] [CrossRef] [PubMed]
- Solano, C.; García, B.; Valle, J.; Berasain, C.; Ghigo, J.M.; Gamazo, C.; Lasa, I. Genetic analysis of Salmonella enteritidis biofilm formation: Critical role of cellulose. Mol. Microbiol. 2002, 43, 793–808. [Google Scholar] [CrossRef]
- White, A.P.; Gibson, D.L.; Collinson, S.K.; Banser, P.A.; Kay, W.W. Extracellular polysaccharides associated with thin aggregative fimbriae of Salmonella enterica serovar Enteritidis. J. Bacteriol. 2003, 185, 5398–5407. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Shi, H.; Li, Y.; Shi, Z.; Zhang, X.; Baek, C.H.; Mothershead, T.; Curtiss, R., 3rd. A colanic acid operon deletion mutation enhances induction of early antibody responses by live attenuated Salmonella vaccine strains. Infect. Immun. 2013, 81, 3148–3162. [Google Scholar] [CrossRef] [Green Version]
- Latasa, C.; Roux, A.; Toledo-Arana, A.; Ghigo, J.M.; Gamazo, C.; Penadés, J.R.; Lasa, I. BapA, a large secreted protein required for biofilm formation and host colonization of Salmonella enterica serovar Enteritidis. Mol. Microbiol. 2005, 58, 1322–1339. [Google Scholar] [CrossRef]
- Vestby, L.K.; Møretrø, T.; Langsrud, S.; Heir, E.; Nesse, L.L. Biofilm forming abilities of Salmonella are correlated with persistence in fish meal- and feed factories. BMC Vet. Res. 2009, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Stoller, A.; Stevens, M.J.A.; Stephan, R.; Guldimann, C. Characteristics of Listeria Monocytogenes Strains Persisting in a Meat Processing Facility over a 4-Year Period. Pathogens 2019, 8, 32. [Google Scholar] [CrossRef] [Green Version]
- Maes, S.; Vackier, T.; Huu, S.N.; Heyndrickx, M.; Steenackers, H.; Sampers, I.; Raes, K.; Verplaetse, A.; De Reu, K. Occurrence and characterisation of biofilms in drinking water systems of broiler houses. BMC Microbiol. 2019, 19, 77. [Google Scholar] [CrossRef] [Green Version]
- Giaouris, E.; Heir, E.; Hébraud, M.; Chorianopoulos, N.; Langsrud, S.; Møretrø, T.; Habimana, O.; Desvaux, M.; Renier, S.; Nychas, G.J. Attachment and biofilm formation by foodborne bacteria in meat processing environments: Causes, implications, role of bacterial interactions and control by alternative novel methods. Meat Sci. 2014, 97, 298–309. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, G.; Russell, A.D. Antiseptics and disinfectants: Activity, action, and resistance. Clin. Microbiol. Rev. 1999, 12, 147–179. [Google Scholar] [CrossRef] [Green Version]
- EN 14885:2022-10; Chemical Disinfectants and Antiseptics—Application of European Standards for Chemical Disinfectants and Antiseptics. European Committee for Standardization (CEN): Brussels, Belgium, 2022.
- Møretrø, T.; Vestby, L.K.; Nesse, L.L.; Storheim, S.E.; Kotlarz, K.; Langsrud, S. Evaluation of efficacy of disinfectants against Salmonella from the feed industry. J. Appl. Microbiol. 2009, 106, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Brunke, M.S.; Konrat, K.; Schaudinn, C.; Piening, B.; Pfeifer, Y.; Becker, L.; Schwebke, I.; Arvand, M. Tolerance of biofilm of a carbapenem-resistant Klebsiella pneumoniae involved in a duodenoscopy-associated outbreak to the disinfectant used in reprocessing. Antimicrob. Resist. Infect. Control. 2022, 11, 81. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.S.; Townsend, K.M.; Fenwick, S.G.; Trengove, R.D.; O’Handley, R.M. Comparative susceptibility of planktonic and 3-day-old Salmonella Typhimurium biofilms to disinfectants. J. Appl. Microbiol. 2010, 108, 2222–2228. [Google Scholar] [CrossRef] [PubMed]
- Merchel Piovesan Pereira, B.; Wang, X.; Tagkopoulos, I. Biocide-Induced Emergence of Antibiotic Resistance in Escherichia coli. Front. Microbiol. 2021, 12, 640923. [Google Scholar] [CrossRef]
- Nordholt, N.; Kanaris, O.; Schmidt, S.B.I.; Schreiber, F. Persistence against benzalkonium chloride promotes rapid evolution of tolerance during periodic disinfection. Nat. Commun. 2021, 12, 6792. [Google Scholar] [CrossRef]
- Konrat, K.; Schwebke, I.; Laue, M.; Dittmann, C.; Levin, K.; Andrich, R.; Arvand, M.; Schaudinn, C. The Bead Assay for Biofilms: A Quick, Easy and Robust Method for Testing Disinfectants. PLoS ONE 2016, 11, e0157663. [Google Scholar] [CrossRef] [Green Version]
- Vestby, L.K.; Nesse, L.L. Wound care antiseptics—performance differences against Staphylococcus aureus in biofilm. Acta Vet. Scand. 2015, 57, 22. [Google Scholar] [CrossRef] [Green Version]
- Vestby, L.K.; Lönn-Stensrud, J.; Møretrø, T.; Langsrud, S.; Aamdal-Scheie, A.; Benneche, T.; Nesse, L.L. A synthetic furanone potentiates the effect of disinfectants on Salmonella in biofilm. J. Appl. Microbiol. 2010, 108, 771–778. [Google Scholar] [CrossRef] [Green Version]
- EN 1656:2019-10; Chemical Disinfectants and Antiseptics—Quantitative Suspension Test for the Evaluation of Bactericidal Activity of Chemical Disinfectants and Antiseptics Used in the Veterinary Area—Test Method and Requirements (Phase 2, Step 1). European Committee for Standardization (CEN): Brussels, Belgium, 2019; 2.
- Römling, U.; Rohde, M.; Olsén, A.; Normark, S.; Reinköster, J. AgfD, the checkpoint of multicellular and aggregative behaviour in Salmonella typhimurium regulates at least two independent pathways. Mol. Microbiol. 2000, 36, 10–23. [Google Scholar] [CrossRef]
- Osland, A.M.; Vestby, L.K.; Nesse, L.L. The Effect of Disinfectants on Quinolone Resistant E. coli (QREC) in Biofilm. Microorganisms 2020, 8, 1831. [Google Scholar] [CrossRef]
- Fernandes, S.; Gomes, I.B.; Sousa, S.F.; Simões, M. Antimicrobial Susceptibility of Persister Biofilm Cells of Bacillus cereus and Pseudomonas fluorescens. Microorganisms 2022, 10, 160. [Google Scholar] [CrossRef]
- Gosling, R.J.; Mawhinney, I.; Vaughan, K.; Davies, R.H.; Smith, R.P. Efficacy of disinfectants and detergents intended for a pig farm environment where Salmonella is present. Vet. Microbiol. 2017, 204, 46–53. [Google Scholar] [CrossRef]
- Corcoran, M.; Morris, D.; De Lappe, N.; O’Connor, J.; Lalor, P.; Dockery, P.; Cormican, M. Commonly used disinfectants fail to eradicate Salmonella enterica biofilms from food contact surface materials. Appl. Environ. Microbiol. 2014, 80, 1507–1514. [Google Scholar] [CrossRef] [Green Version]
- Desinfektionsmittel-Kommission im VAH. Anforderungen und Methoden zur VAH-Zertifizierung Chemischer Desinfektionsverfahren; Section; VAH: Bonn, Germany, 2022; Available online: https://vah-online.de/files/download/ebooks/eBook_VAH_Methoden_Anforderungen.pdf (accessed on 9 November 2022).
- Chino, T.; Nukui, Y.; Morishita, Y.; Moriya, K. Morphological bactericidal fast-acting effects of peracetic acid, a high-level disinfectant, against Staphylococcus aureus and Pseudomonas aeruginosa biofilms in tubing. Antimicrob. Resist. Infect. Control. 2017, 6, 122. [Google Scholar] [CrossRef] [Green Version]
- von Hertwig, A.M.; Prestes, F.S.; Nascimento, M.S. Biofilm formation and resistance to sanitizers by Salmonella spp. Isolated from the peanut supply chain. Food Res. Int. 2022, 152, 110882. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, D.C.; Junior, A.F.; Kaneno, R.; Silva, M.G.; Junior, J.P.A.; Silva, N.C.; Rall, V.L. Ability of Salmonella spp. to produce biofilm is dependent on temperature and surface material. Foodborne Pathog. Dis. 2014, 11, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Ben Abdallah, F.; Lagha, R.; Said, K.; Kallel, H.; Gharbi, J. Detection of cell surface hydrophobicity, biofilm and fimbirae genes in Salmonella isolated from tunisian clinical and poultry meat. Iran J. Public Health 2014, 43, 423–431. [Google Scholar] [PubMed]
- Römling, U.; Bokranz, W.; Rabsch, W.; Zogaj, X.; Nimtz, M.; Tschäpe, H. Occurrence and regulation of the multicellular morphotype in Salmonella serovars important in human disease. Int. J. Med. Microbiol. 2003, 293, 273–285. [Google Scholar] [CrossRef]
- Moraes, J.O.; Cruz, E.A.; Pinheiro, I.; Oliveira, T.C.M.; Alvarenga, V.; Sant’Ana, A.S.; Magnani, M. An ordinal logistic regression approach to predict the variability on biofilm formation stages by five Salmonella enterica strains on polypropylene and glass surfaces as affected by pH, temperature and NaCl. Food Microbiol. 2019, 8, 95–103. [Google Scholar] [CrossRef]
- Rodríguez-Melcón, C.; Riesco-Peláez, F.; Carballo, J.; García-Fernández, C.; Capita, R.; Alonso-Calleja, C. Structure and viability of 24- and 72-h-old biofilms formed by four pathogenic bacteria on polystyrene and glass contact surfaces. Food Microbiol. 2018, 76, 513–517. [Google Scholar] [CrossRef]
- Ban, G.H.; Park, S.H.; Kim, S.O.; Ryu, S.; Kang, D.H. Synergistic effect of steam and lactic acid against Escherichia coli O157:H7, Salmonella Typhimurium, and Listeria monocytogenes biofilms on polyvinyl chloride and stainless steel. Int. J. Food Microbiol. 2012, 157, 218–223. [Google Scholar] [CrossRef]
- Merino, L.; Trejo, F.M.; De Antoni, G.; Golowczyc, M.A. Lactobacillus strains inhibit biofilm formation of Salmonella sp. isolates from poultry. Food Res. Int. 2019, 123, 258–265. [Google Scholar] [CrossRef]
- Uhlich, G.A.; Cooke, P.H.; Solomon, E.B. Analyses of the red-dry-rough phenotype of an Escherichia coli O157:H7 strain and its role in biofilm formation and resistance to antibacterial agents. Appl. Environ. Microbiol. 2006, 72, 2564–2572. [Google Scholar] [CrossRef] [Green Version]
- Chmielewski, R.A.N.; Frank, J.F. Biofilm Formation and Control in Food Processing Facilities. Compr. Rev. Food Sci. Food Saf. 2003, 2, 22–32. [Google Scholar] [CrossRef]
- Simões, L.C.; Gomes, I.B.; Sousa, H.; Borges, A.; Simões, M. Biofilm formation under high shear stress increases resilience to chemical and mechanical challenges. Biofouling 2022, 38, 1–12. [Google Scholar] [CrossRef]
- Fanesi, A.; Lavayssière, M.; Breton, C.; Bernard, O.; Briandet, R.; Lopes, F. Shear stress affects the architecture and cohesion of Chlorella vulgaris biofilms. Sci. Rep. 2021, 11, 4002. [Google Scholar] [CrossRef]
- Buckingham-Meyer, K.; Goeres, D.M.; Hamilton, M.A. Comparative evaluation of biofilm disinfectant efficacy tests. J. Microbiol. Methods 2007, 70, 236–244. [Google Scholar] [CrossRef]
- Pereira, M.O.; Kuehn, M.; Wuertz, S.; Neu, T.; Melo, L.F. Effect of flow regime on the architecture of a Pseudomonas fluorescens biofilm. Biotechnol. Bioeng. 2002, 78, 164–171. [Google Scholar] [CrossRef]
- Goeres, D.M.; Loetterle, L.R.; Hamilton, M.A.; Murga, R.; Kirby, D.W.; Donlan, R.M. Statistical assessment of a laboratory method for growing biofilms. Microbiology 2005, 151 Pt 3, 757–762. [Google Scholar] [CrossRef] [Green Version]
- Goeres, D.M.; Hamilton, M.A.; Beck, N.A.; Buckingham-Meyer, K.; Hilyard, J.D.; Loetterle, L.R.; Lorenz, L.A.; Walker, D.K.; Stewart, P.S. A method for growing a biofilm under low shear at the air-liquid interface using the drip flow biofilm reactor. Nat. Protoc. 2009, 4, 783–788. [Google Scholar] [CrossRef]
- Sternberg, C.; Tolker-Nielsen, T. Growing and analyzing biofilms in flow cells. Curr. Protoc. Microbiol. 2006, 1, 1B.2.1–1B.2.15. [Google Scholar] [CrossRef]
- Schonewille, E.; Nesse, L.L.; Hauck, R.; Windhorst, D.; Hafez, H.M.; Vestby, L.K. Biofilm building capacity of Salmonella enterica strains from the poultry farm environment. FEMS Immunol. Med. Microbiol. 2012, 65, 360–365. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Veterinary Area | Food, Industrial, Domestic, and Institutional Area | Medical Area | |
|---|---|---|---|
| Phase 2, Step 1 | Quantitative suspension test EN 1656:2019 (≥5 log10) | Quantitative suspension test EN 1276:2019 (≥5 log10) | Quantitative suspension test EN 13727:2012+A2:2015 (≥5 log10) |
| Phase 2, Step 2 | Quantitative non-porous surface test EN 14349:2012 (≥4 log10) Quantitative porous surface test EN 16437:2014+A1:2019 (≥4 log10) | Quantitative non-porous surface test EN 13697:2015+A1:2019 (≥4 log10) | Quantitative carrier test EN 14561:2006 (instruments, ≥5 log10) Quantitative non-porous surface test EN 16615:2015 (surfaces w/mechanical action, ≥5 log10) Quantitative non-porous surface test EN 17387:2021 (surfaces w/o mechanical action, ≥5 log10) |
| Glutaraldehyde | GBA | PCA | SSCA | |||
|---|---|---|---|---|---|---|
| CFU Reduction | ATCC 14028 | ATCC 13311 | ATCC 14028 | ATCC 13311 | ATCC 14028 | ATCC 13311 |
| ≥4 log10 | 0.5% | 0.1% | 0.1% | - | 1% | 0.5% |
| ≥5 log10 | 0.5% | 0.1% | 0.5% | - | n.a. * | n.a. * |
| Peracetic Acid | GBA | PCA | SSCA | |||
|---|---|---|---|---|---|---|
| CFU Reduction | ATCC 14028 | ATCC 13311 | ATCC 14028 | ATCC 13311 | ATCC 14028 | ATCC 13311 |
| ≥4 log10 | 0.05% | 0.1% | 0.002% | - | 0.02% | 0.02% |
| ≥5 log10 | 0.1% | 0.1% | 0.005% | - | n.a. * | n.a. * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Richter, A.M.; Konrat, K.; Osland, A.M.; Brook, E.; Oastler, C.; Vestby, L.K.; Gosling, R.J.; Nesse, L.L.; Arvand, M. Evaluation of Biofilm Cultivation Models for Efficacy Testing of Disinfectants against Salmonella Typhimurium Biofilms. Microorganisms 2023, 11, 761. https://doi.org/10.3390/microorganisms11030761
Richter AM, Konrat K, Osland AM, Brook E, Oastler C, Vestby LK, Gosling RJ, Nesse LL, Arvand M. Evaluation of Biofilm Cultivation Models for Efficacy Testing of Disinfectants against Salmonella Typhimurium Biofilms. Microorganisms. 2023; 11(3):761. https://doi.org/10.3390/microorganisms11030761
Chicago/Turabian StyleRichter, Anja M., Katharina Konrat, Ane M. Osland, Emma Brook, Claire Oastler, Lene K. Vestby, Rebecca J. Gosling, Live L. Nesse, and Mardjan Arvand. 2023. "Evaluation of Biofilm Cultivation Models for Efficacy Testing of Disinfectants against Salmonella Typhimurium Biofilms" Microorganisms 11, no. 3: 761. https://doi.org/10.3390/microorganisms11030761