
First Detection and Molecular Characterization of Usutu Virus in Culex pipiens Mosquitoes Collected in Romania
 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
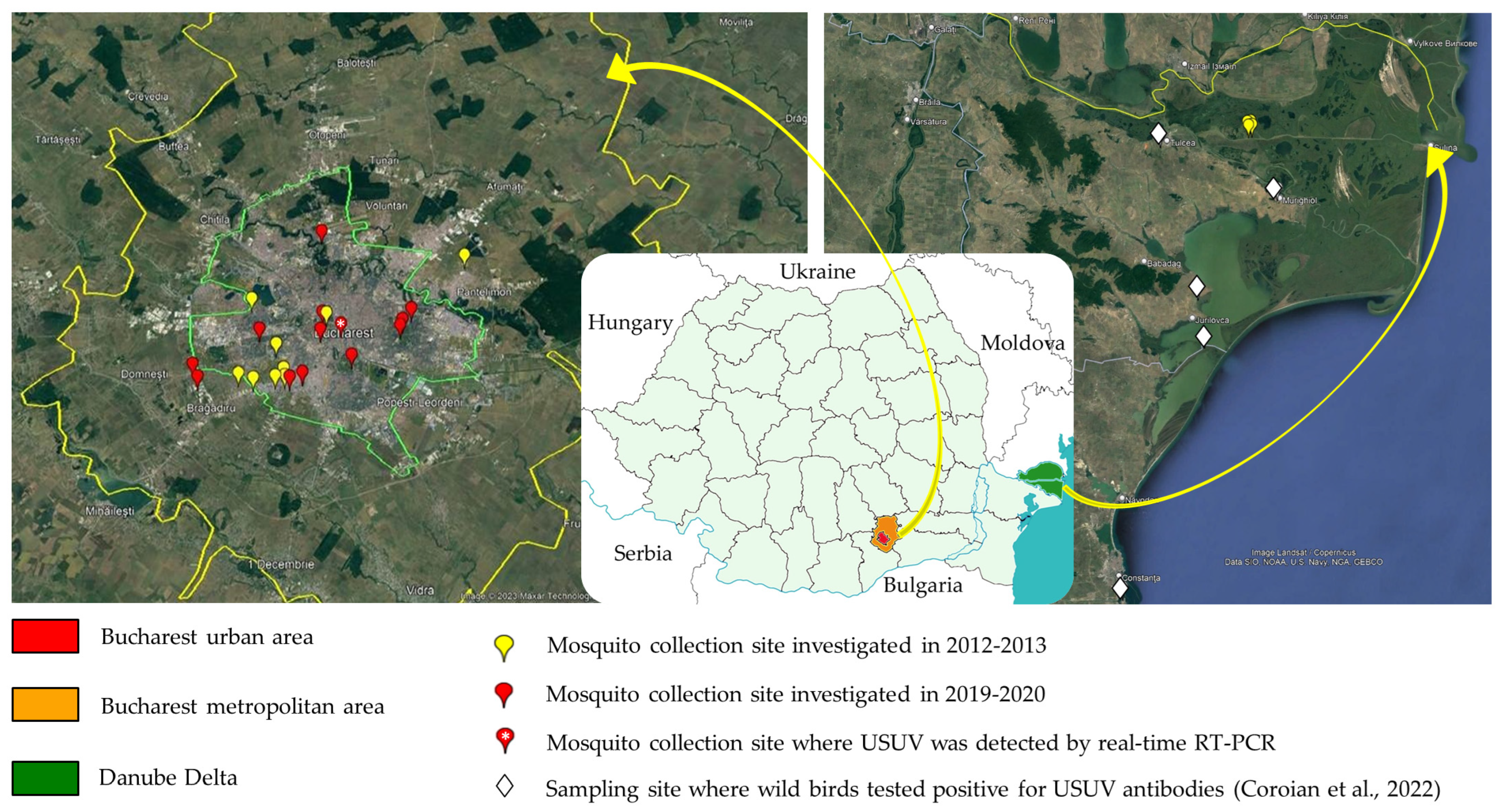
2.1. Sampling Sites and Mosquito Collection
2.2. Mosquito Processing and RNA Extraction
2.3. Detection of USUV by Real-Time RT-PCR
2.4. Sequencing and Phylogenetic Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zannoli, S.; Sambri, V. West Nile Virus and Usutu Virus Co-Circulation in Europe: Epidemiology and Implications. Microorganisms 2019, 7, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilibic-Cavlek, T.; Petrovic, T.; Savic, V.; Barbic, L.; Tabain, I.; Stevanovic, V.; Klobucar, A.; Mrzljak, A.; Ilic, M.; Bogdanic, M.; et al. Epidemiology of Usutu Virus: The European Scenario. Pathogens 2020, 9, 699. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.C.; Simpson, D.I.H.; Haddow, A.J.; Knight, E.M. The Isolation of West Nile Virus from Man and of Usutu Virus from the Bird-Biting Mosquito Mansonia Aurites(Theobald) in the Entebbe Area of Uganda. Ann. Trop. Med. Parasitol. 1964, 58, 367–374. [Google Scholar] [CrossRef]
- Weissenböck, H.; Kolodziejek, J.; Url, A.; Lussy, H.; Rebel-Bauder, B.; Nowotny, N. Emergence of Usutu virus, an African Mosquito-Borne Flavivirus of the Japanese Encephalitis Virus Group, Central Europe. Emerg. Infect. Dis. 2002, 8, 652–656. [Google Scholar] [CrossRef]
- Weissenböck, H.; Bakonyi, T.; Rossi, G.; Mani, P.; Nowotny, N. Usutu Virus, Italy, 1996. Emerg. Infect. Dis. 2013, 19, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Bakonyi, T.; Erdélyi, K.; Ursu, K.; Ferenczi, E.; Csorgo, T.; Lussy, H.; Chvala, S.; Bukovsky, C.; Meister, T.; Weissenböck, H.; et al. Emergence of Usutu Virus in Hungary. J. Clin. Microbiol. 2007, 45, 3870–3874. [Google Scholar] [CrossRef] [Green Version]
- Steinmetz, H.W.; Bakonyi, T.; Weissenböck, H.; Hatt, J.-M.; Eulenberger, U.; Robert, N.; Hoop, R.; Nowotny, N. Emergence and establishment of Usutu virus infection in wild and captive avian species in and around Zurich, Switzerland—Genomic and pathologic comparison to other central European outbreaks. Veter- Microbiol. 2011, 148, 207–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manarolla, G.; Bakonyi, T.; Gallazzi, D.; Crosta, L.; Weissenböck, H.; Dorrestein, G.; Nowotny, N. Usutu virus in wild birds in northern Italy. Veter- Microbiol. 2010, 141, 159–163. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, U.; Bergmann, F.; Fischer, D.; Müller, K.; Holicki, C.M.; Sadeghi, B.; Sieg, M.; Keller, M.; Schwehn, R.; Reuschel, M.; et al. Spread of West Nile Virus and Usutu Virus in the German Bird Population, 2019–2020. Microorganisms 2022, 10, 807. [Google Scholar] [CrossRef]
- Fros, J.J.; Miesen, P.; Vogels, C.B.; Gaibani, P.; Sambri, V.; Martina, B.E.; Koenraadt, C.J.; van Rij, R.P.; Vlak, J.M.; Takken, W.; et al. Comparative Usutu and West Nile virus transmission potential by local Culex pipiens mosquitoes in north-western Europe. One Health 2015, 1, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Klobucar, A.; Benic, N.; Krajcar, D.; Kosanovic-Licina, M.L.; Tesic, V.; Merdic, E.; Vrucina, I.; Savić, V.; Barbic, L.; Stevanovic, V.; et al. An overview of mosquitoes and emerging arboviral infections in the Zagreb area, Croatia. J. Infect. Dev. Ctries. 2016, 10, 1286–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camp, J.V.; Kolodziejek, J.; Nowotny, N. Targeted surveillance reveals native and invasive mosquito species infected with Usutu virus. Parasites Vectors 2019, 12, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engel, D.; Jöst, H.; Wink, M.; Börstler, J.; Bosch, S.; Garigliany, M.-M.; Jöst, A.; Czajka, C.; Lühken, R.; Ziegler, U.; et al. Reconstruction of the Evolutionary History and Dispersal of Usutu Virus, a Neglected Emerging Arbovirus in Europe and Africa. Mbio 2016, 7, e01938-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadar, D.; Lühken, R.; van der Jeugd, H.; Garigliany, M.; Ziegler, U.; Keller, M.; Lahoreau, J.; Lachmann, L.; Becker, N.; Kik, M.; et al. Widespread activity of multiple lineages of Usutu virus, western Europe, 2016. Eurosurveillance 2017, 22, 30452. [Google Scholar] [CrossRef] [Green Version]
- Cadar, D.; Simonin, Y. Human Usutu Virus Infections in Europe: A New Risk on Horizon? Viruses 2022, 15, 77. [Google Scholar] [CrossRef]
- Coroian, M.; Silaghi, C.; Tews, B.A.; Baltag, E.; Marinov, M.; Alexe, V.; Kalmár, Z.; Cintia, H.; Lupșe, M.S.; Mihalca, A.D. Serological Survey of Mosquito-Borne Arboviruses in Wild Birds from Important Migratory Hotspots in Romania. Pathogens 2022, 11, 1270. [Google Scholar] [CrossRef]
- Coroian, M.; Mihalca, A.D.; Dobler, G.; Euringer, K.; Girl, P.; Borșan, S.-D.; Kalmár, Z.; Briciu, V.T.; Flonta, M.; Topan, A.; et al. Seroprevalence Rates against West Nile, Usutu, and Tick-Borne Encephalitis Viruses in Blood-Donors from North-Western Romania. Int. J. Environ. Res. Public Heal. 2022, 19, 8182. [Google Scholar] [CrossRef]
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes and their Control, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2010; p. 577. [Google Scholar]
- Prioteasa, F.L.; Tiron, G.V.; Dinu, S.; Fălcuţă, E. Using Rapid Analyte Measurement Platform (RAMP) as a Tool for an Early Warning System Assessing West Nile Virus Epidemiological Risk in Bucharest, Romania. Trop. Med. Infect. Dis. 2022, 7, 327. [Google Scholar] [CrossRef]
- Del Amo, J.; Sotelo, E.; Fernández-Pinero, J.; Gallardo, C.; Llorente, F.; Agüero, M.; Jiménez-Clavero, M.A. A novel quantitative multiplex real-time RT-PCR for the simultaneous detection and differentiation of West Nile virus lineages 1 and 2, and of Usutu virus. J. Virol. Methods 2013, 189, 321–327. [Google Scholar] [CrossRef]
- Nikolay, B.; Weidmann, M.; Dupressoir, A.; Faye, O.; Boye, C.; Diallo, M.; Sall, A. Development of a Usutu virus specific real-time reverse transcription PCR assay based on sequenced strains from Africa and Europe. J. Virol. Methods 2014, 197, 51–54. [Google Scholar] [CrossRef] [Green Version]
- Hönig, V.; Palus, M.; Kaspar, T.; Zemanova, M.; Majerova, K.; Hofmannova, L.; Papezik, P.; Sikutova, S.; Rettich, F.; Hubalek, Z.; et al. Multiple Lineages of Usutu Virus (Flaviviridae, Flavivirus) in Blackbirds (Turdus merula) and Mosquitoes (Culex pipiens, Cx. modestus) in the Czech Republic (2016–2019). Microorganisms 2019, 7, 568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.-H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef]
- Zecchin, B.; Fusaro, A.; Milani, A.; Schivo, A.; Ravagnan, S.; Ormelli, S.; Mavian, C.; Michelutti, A.; Toniolo, F.; Barzon, L.; et al. The central role of Italy in the spatial spread of USUTU virus in Europe. Virus Evol. 2021, 7, veab048. [Google Scholar] [CrossRef]
- Gaibani, P.; Cavrini, F.; Gould, E.A.; Rossini, G.; Pierro, A.; Landini, M.P.; Sambri, V. Comparative Genomic and Phylogenetic Analysis of the First Usutu Virus Isolate from a Human Patient Presenting with Neurological Symptoms. PLoS ONE 2013, 8, e64761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolodziejek, J.; Marinov, M.; Kiss, B.J.; Alexe, V.; Nowotny, N. The Complete Sequence of a West Nile Virus Lineage 2 Strain Detected in a Hyalomma marginatum marginatum Tick Collected from a Song Thrush (Turdus philomelos) in Eastern Romania in 2013 Revealed Closest Genetic Relationship to Strain Volgograd 2007. PLoS ONE 2014, 9, e109905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinu, S.; Cotar, A.I.; Panculescu-Gatej, I.R.; Falcuta, E.; Prioteasa, F.L.; Sirbu, A.; Oprisan, G.; Badescu, D.; Reiter, P.; Ceianu, C.S. West Nile virus circulation in south-eastern Romania, 2011 to 2013. Euro Surveill. Bull. Eur. Sur Les Mal. Transm. = Eur. Commun. Dis. Bull. 2015, 20, 21130. [Google Scholar] [CrossRef] [Green Version]
- Cotar, A.I.; Falcuta, E.; Prioteasa, L.F.; Dinu, S.; Ceianu, C.S.; Paz, S. Transmission Dynamics of the West Nile Virus in Mosquito Vector Populations under the Influence of Weather Factors in the Danube Delta, Romania. EcoHealth 2016, 13, 796–807. [Google Scholar] [CrossRef]
- Tomazatos, A.; Jansen, S.; Pfister, S.; Török, E.; Maranda, I.; Horváth, C.; Keresztes, L.; Spînu, M.; Tannich, E.; Jöst, H.; et al. Ecology of West Nile Virus in the Danube Delta, Romania: Phylogeography, Xenosurveillance and Mosquito Host-Feeding Patterns. Viruses 2019, 11, 1159. [Google Scholar] [CrossRef] [Green Version]
- Tsai, T.F.; Popovici, F.; Cernescu, C.; Campbell, G.L.; Nedelcu, N.I. West Nile encephalitis epidemic in southeastern Romania. Lancet 1998, 352, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Sirbu, A.; Ceianu, C.S.; Panculescu-Gatej, R.I.; Vazquez, A.; Tenorio, A.; Rebreanu, R.; Niedrig, M.; Nicolescu, G.; Pistol, A. Outbreak of West Nile virus infection in humans, Romania, July to October 2010. Euro Surveill. Bull. Eur. Sur Les Mal. Transm. = Eur. Commun. Dis. Bull. 2011, 16, 19762. [Google Scholar] [CrossRef]
- Popescu, C.P.; Florescu, S.A.; Cotar, A.I.; Badescu, D.; Ceianu, C.S.; Zaharia, M.; Tardei, G.; Codreanu, D.; Ceausu, E.; Ruta, S.M. Re-emergence of severe West Nile virus neuroinvasive disease in humans in Romania, 2012 to 2017–implications for travel medicine. Travel Med. Infect. Dis. 2018, 22, 30–35. [Google Scholar] [CrossRef]
- Savage, H.M.; Romanca, C.; Vladimirescu, A.; Tsai, T.F.; Ceianu, C.; Karabatsos, N.; Lanciotti, R.; Ungureanu, A.; Laiv, L.; Nicolescu, G. Entomologic and avian investigations of an epidemic of West Nile fever in Romania in 1996, with serologic and molecular characterization of a virus isolate from mosquitoes. Am. J. Trop. Med. Hyg. 1999, 61, 600–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotar, A.I.; Fălcuță, E.; Dinu, S.; Necula, A.; Bîrluțiu, V.; Ceianu, C.S.; Prioteasa, F.L. West Nile virus lineage 2 in Romania, 2015–2016: Co-circulation and strain replacement. Parasites Vectors 2018, 11, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilibic-Cavlek, T.; Savic, V.; Petrovic, T.; Toplak, I.; Barbic, L.; Petric, D.; Tabain, I.; Hrnjakovic-Cvjetkovic, I.; Bogdanic, M.; Klobucar, A.; et al. Emerging Trends in the Epidemiology of West Nile and Usutu Virus Infections in Southern Europe. Front. Veter- Sci. 2019, 6, 437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgado, R.; Hawks, S.A.; Frere, F.; Vázquez, A.; Huang, C.Y.-H.; Duggal, N.K. West Nile Virus Vaccination Protects against Usutu Virus Disease in Mice. Viruses 2021, 13, 2352. [Google Scholar] [CrossRef]
- Wang, H.; Abbo, S.R.; Visser, T.M.; Westenberg, M.; Geertsema, C.; Fros, J.J.; Koenraadt, C.J.M.; Pijlman, G.P. Competition between Usutu virus and West Nile virus during simultaneous and sequential infection of Culex pipiens mosquitoes. Emerg. Microbes Infect. 2020, 9, 2642–2652. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Transmission of West Nile Virus, July to December 2019–Table of Cases, 2019 Transmission Season. Available online: https://www.ecdc.europa.eu/en/publications-data/transmission-west-nile-virus-july-december-2019-table-cases-2019-transmission (accessed on 23 February 2023).




{kind=link}
{kind=link}
{kind=link}
| Collection Area | Mosquito Species | 2012 | 2013 | 2019 | 2020 | Total per Species |
|---|---|---|---|---|---|---|
| Number of Mosquitoes Collected/Number of Pools | ||||||
| Bucharest | Culex pipiens s.l. | 1791/53 | 1904/56 | 5053/139 * | 1785/49 | 10,533/297 |
| Aedes albopictius | 0/0 | 0/0 | 59/2 | 0/0 | 59/2 | |
| Danube Delta | Culex pipiens | 1201/44 | 1616/58 | 0/0 | 0/0 | 2817/102 |
| Culex modestus | 236/11 | 135/5 | 0/0 | 0/0 | 371/16 | |
| Anopheles hyrcanus | 0/0 | 13/1 | 0/0 | 0/0 | 13/1 | |
| Anopheles maculipennis | 0/0 | 30/1 | 0/0 | 0/0 | 30/1 | |
| Total per study | 13,823/419 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prioteasa, F.L.; Dinu, S.; Tiron, G.V.; Stancu, I.G.; Fălcuță, E.; Ceianu, C.S.; Cotar, A.I. First Detection and Molecular Characterization of Usutu Virus in Culex pipiens Mosquitoes Collected in Romania. Microorganisms 2023, 11, 684. https://doi.org/10.3390/microorganisms11030684
Prioteasa FL, Dinu S, Tiron GV, Stancu IG, Fălcuță E, Ceianu CS, Cotar AI. First Detection and Molecular Characterization of Usutu Virus in Culex pipiens Mosquitoes Collected in Romania. Microorganisms. 2023; 11(3):684. https://doi.org/10.3390/microorganisms11030684
Chicago/Turabian StylePrioteasa, Florian Liviu, Sorin Dinu, Georgiana Victorița Tiron, Ioana Georgeta Stancu, Elena Fălcuță, Cornelia Svetlana Ceianu, and Ani Ioana Cotar. 2023. "First Detection and Molecular Characterization of Usutu Virus in Culex pipiens Mosquitoes Collected in Romania" Microorganisms 11, no. 3: 684. https://doi.org/10.3390/microorganisms11030684