
Dietary Live Yeast Supplementation Influence on Cow’s Milk, Teat and Bedding Microbiota in a Grass-Diet Dairy System
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Diets
2.3. Milk Production Parameters Measurements
2.4. Microbial Analyses
2.4.1. Sampling Procedure
2.4.2. Microbial Enumeration
2.4.3. DNA Extraction
2.4.4. High-Throughput Sequencing of 16S and ITS Amplicons
2.5. Statistical Analyses
2.5.1. Milk Production Parameters
2.5.2. Microbial Data
3. Results
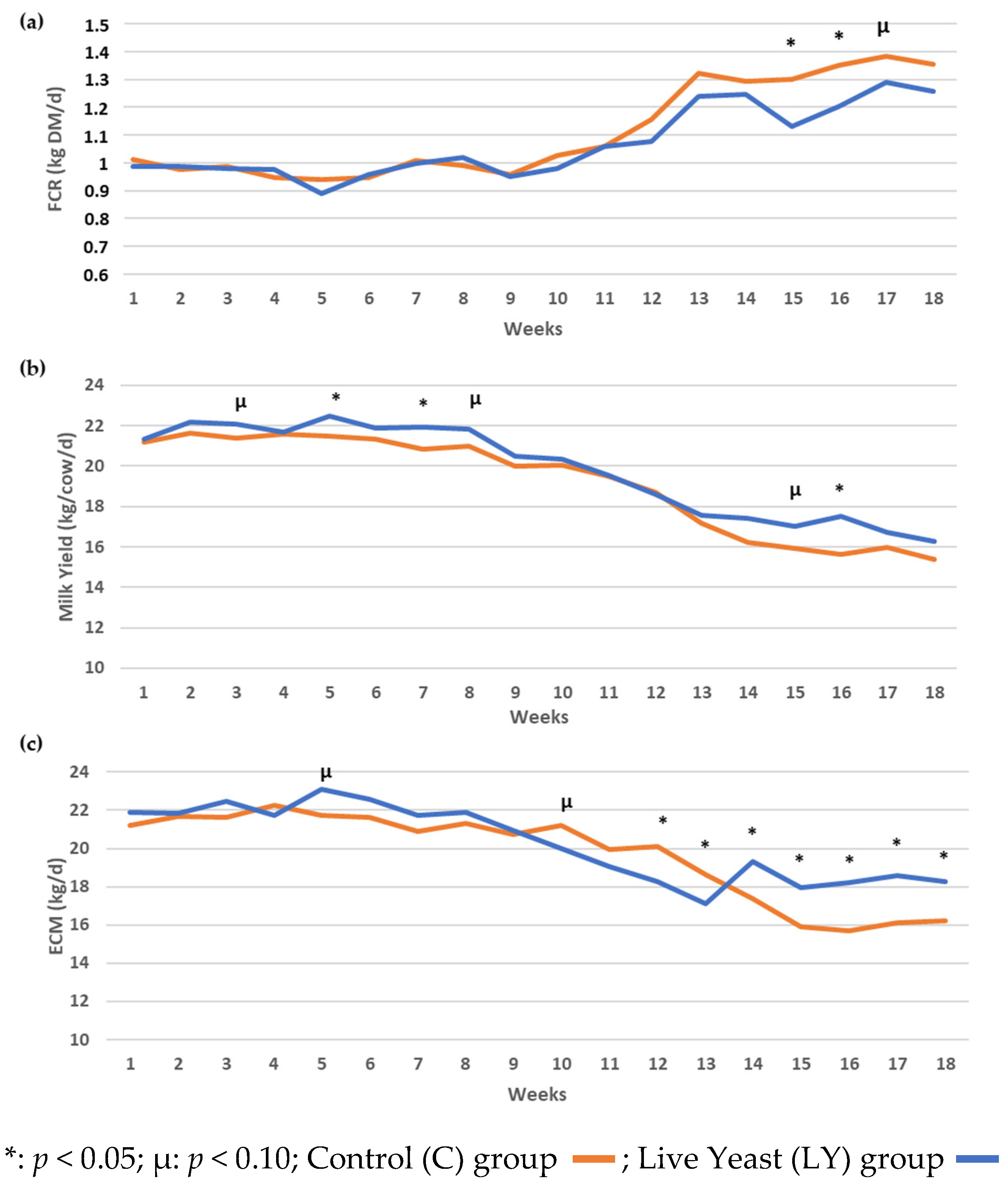
3.1. Animal Performance Data
3.2. Milk Characteristics
3.2.1. Chemical Composition
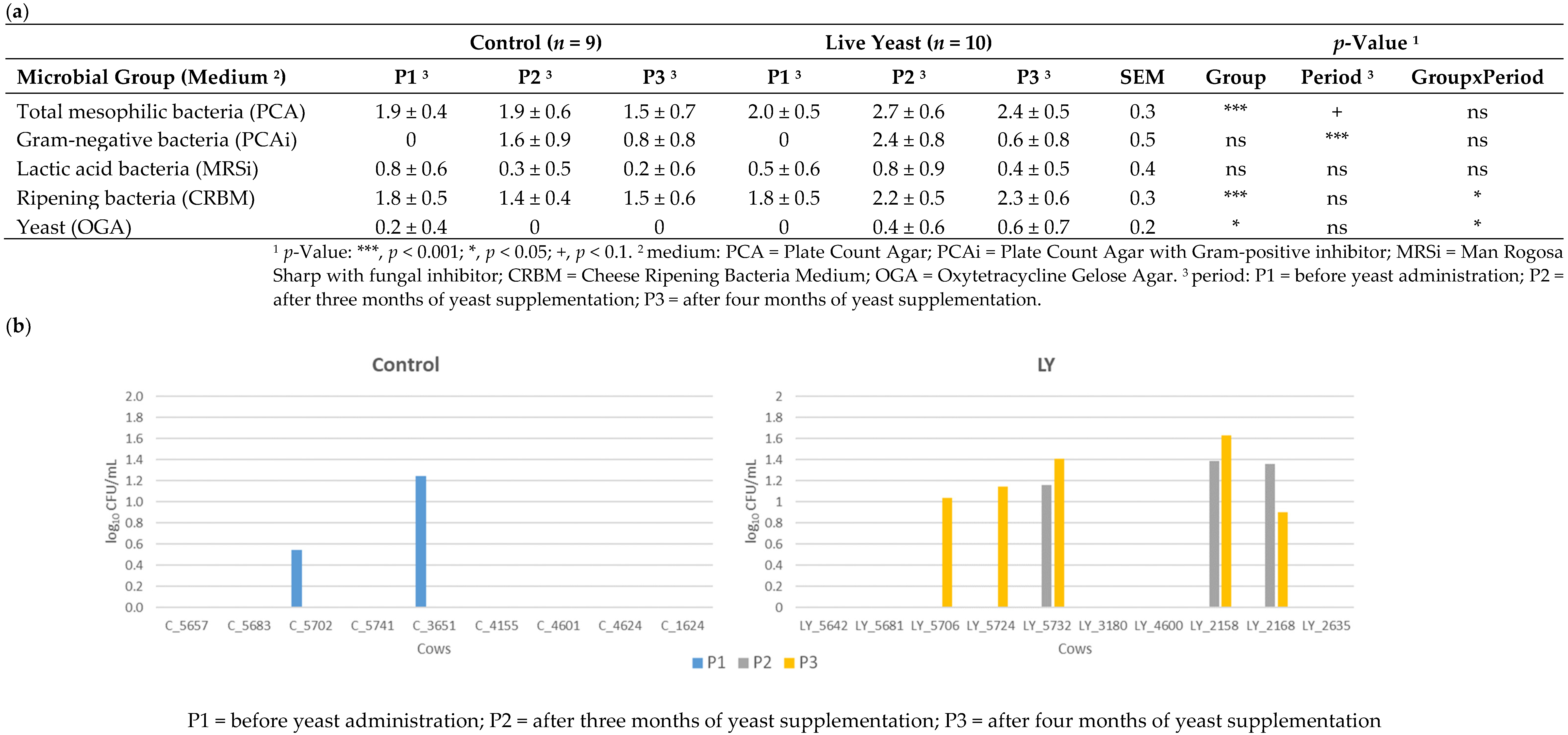
3.2.2. Microbial Composition
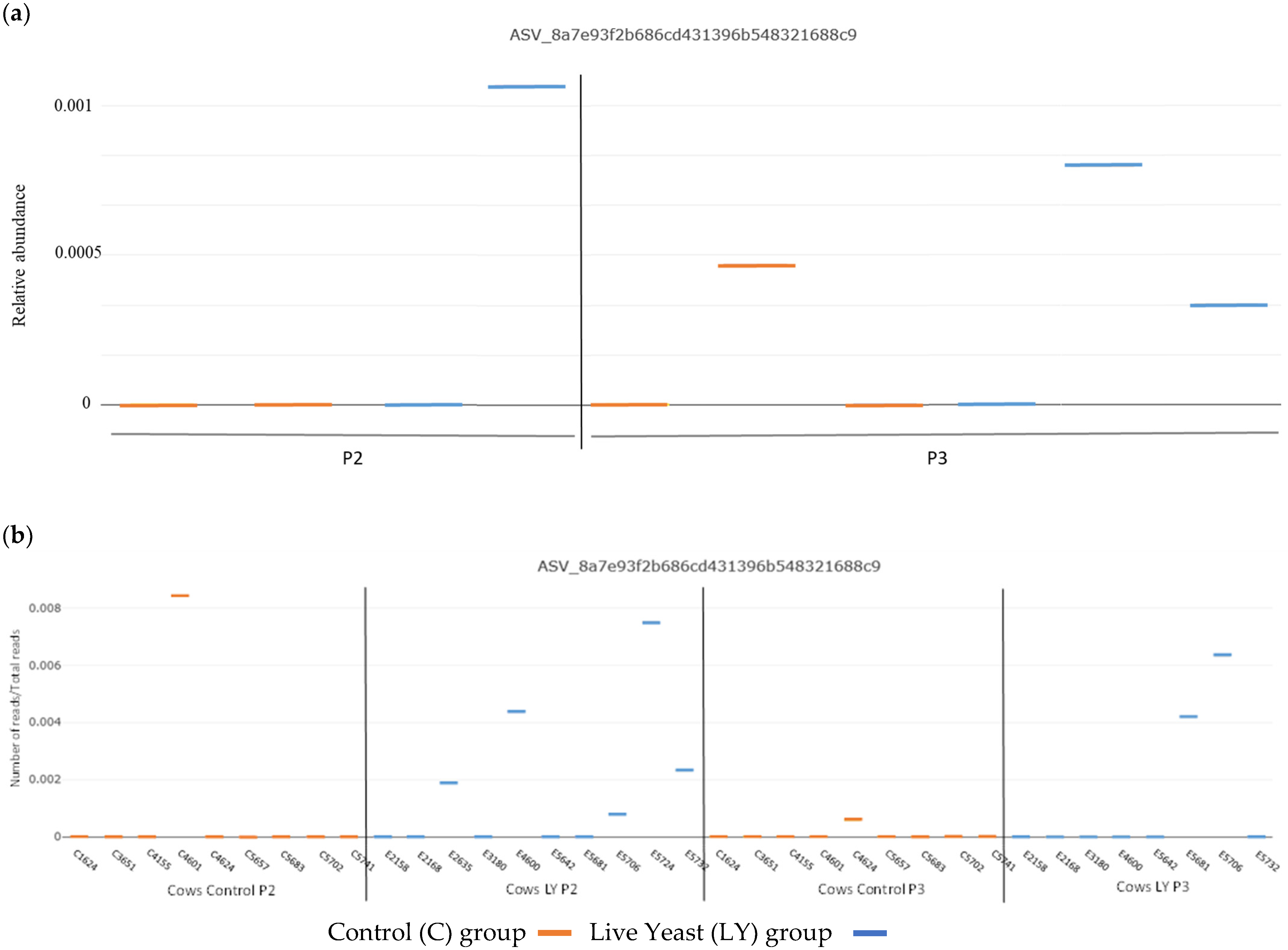
3.3. Dispersion of Saccharomyces Cerevisiae CNCM I-1077 like DNA in the Environment
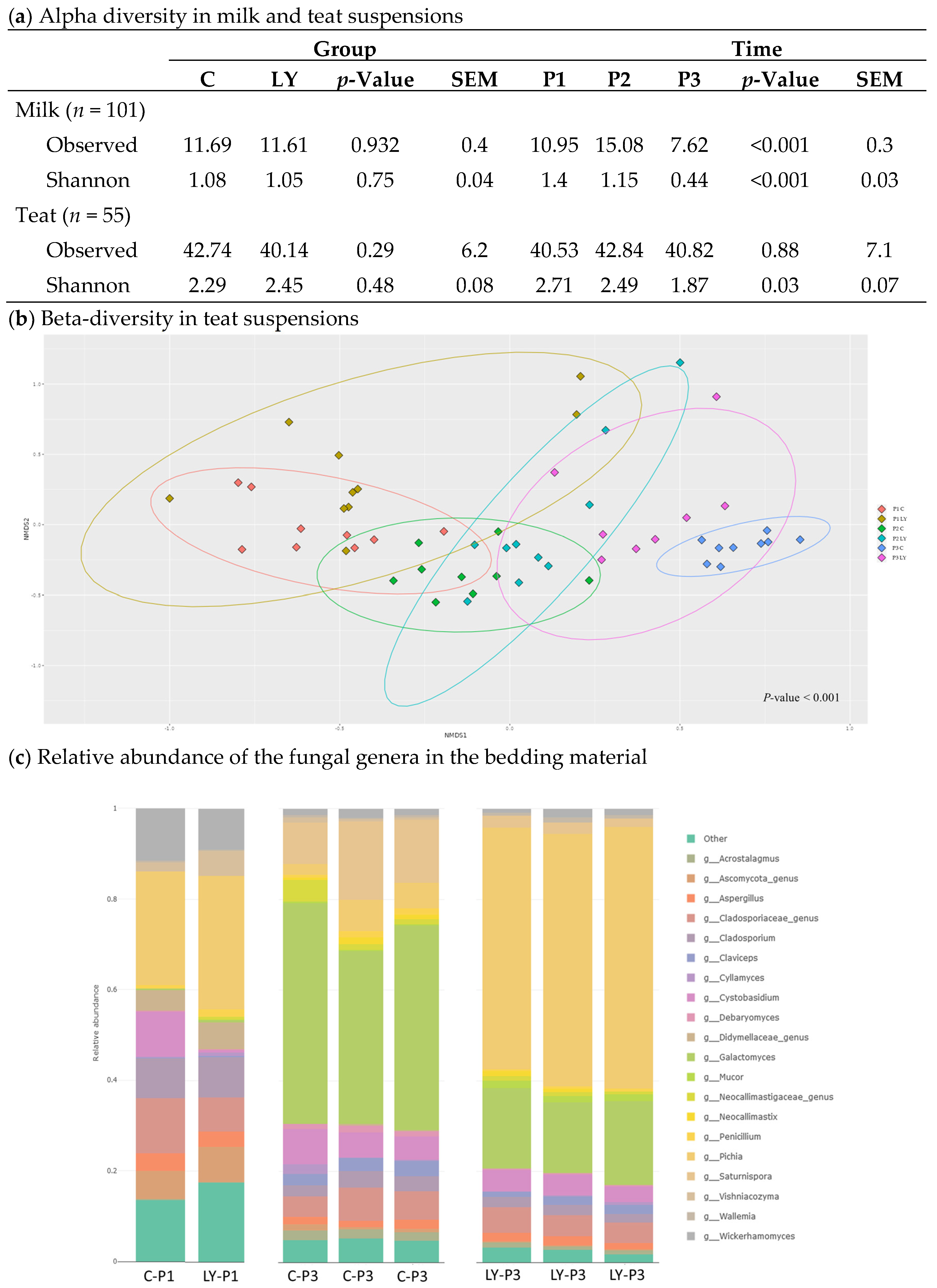
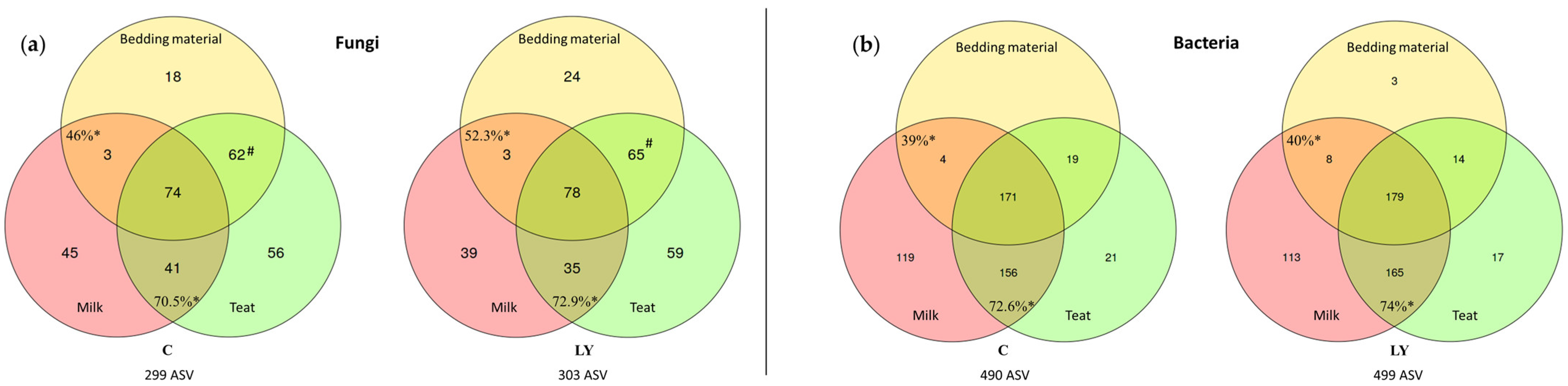
3.4. Diversity and Composition of the Fungal Communities
3.5. Shared Fungal ASVs between Teat and Milk
3.6. Diversity and Composition of the Bacterial Communities
3.7. Shared Bacterial ASVs between Teat and Milk
3.8. Correlation between Bacterial and Fungal Dominant Species in Teat Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Reviews Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rouillé, B.; Devun, J.; Brunschiwig, P. PROTEIN SOURCES IN ANIMAL FEED Les sources de protéines dans l’alimentation du bétail. Oilseeds Fats Crops Lipids 2014, 21, D404. [Google Scholar] [CrossRef] [Green Version]
- Coppa, M.; Cabbidu, A.; Elsässer, M.; Hulin, S.; Lind, V.; Martin, B.; Mosquera-Losada, R.; Peeters, A.; Prache, S.; van den Pol, A.; et al. Grassland resources for extensive farming systems in marginal lands: Major drivers and future scenarios. In Proceedings of the 19th Symposium of the European Grassland Federation, Alghero, Italy, 7–10 May 2017; pp. 39–60. [Google Scholar]
- Shurson, G.C. Yeast and yeast derivatives in feed additives and ingredients: Sources, characteristics, animal responses, and quantification methods. Anim. Feed Sci. Technol. 2018, 235, 60–76. [Google Scholar] [CrossRef]
- Moallem, U.; Lehrer, H.; Livshitz, L.; Zachut, M.; Yakoby, S. The effects of live yeast supplementation to dairy cows during the hot season on production, feed efficiency, and digestibility. J. Dairy Sci. 2009, 92, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Marden, J.P.; Bayourthe, C.; Auclair, E.; Moncoulon, R. A Bioenergetic-Redox Approach to the Effect of Live Yeast on Ruminal pH during Induced Acidosis in Dairy Cow. Am. J. Anal. Chem. 2013, 4, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Humer, E.; Petri, R.M.; Aschenbach, J.R.; Bradford, B.J.; Penner, G.B.; Tafaj, M.; Südekum, K.H.; Zebeli, Q. Invited review: Practical feeding management recommendations to mitigate the risk of subacute ruminal acidosis in dairy cattle. J. Dairy Sci. 2018, 101, 872–888. [Google Scholar] [CrossRef] [Green Version]
- Rossow, H.A.; Riordan, T.; Riordan, A. Effects of addition of a live yeast product on dairy cattle performance. J. Appl. Anim. Res. 2018, 46, 159–163. [Google Scholar] [CrossRef] [Green Version]
- Sturaro, E.; Marchiori, E.; Cocca, G.; Penasa, M.; Ramanzin, M.; Bittante, G. Dairy systems in mountainous areas: Farm animal biodiversity, milk production and destination, and land use. Livest. Sci. 2013, 158, 157–168. [Google Scholar] [CrossRef]
- Deneufbourg, C.; Tormo, H. Évaluation de l ’ impact d ’ une supplémentation en levure vivante sur la microflore du lait cru de vache Evaluation of the impact of a live yeast supplementation on the microflora of raw cow. In Proceedings of the Rencontres autour des Recherches sur les Ruminants, Paris, France, 3–4 December 2014; p. 152. [Google Scholar]
- Grilli, E.; Tormo, H.; Fustini, M.; Deneufbourg, C.; Losio, M.; Formigoni, A.; Chaucheyras-Durand, F.; Durand, H. Is raw milk microbiota influenced by the use of live yeast (Saccharomyces cerevisiae) as ruminant feed additive? Int. J. Dairy Sci. 2016, 11, 124–129. [Google Scholar] [CrossRef] [Green Version]
- Verdier-Metz, I.; Delbes-Paus, C.; Monsallier, F.; Callon, C.; Montel, M.C. Management of microbial diversity at milk production for raw milk cheese making. In Proceedings of the FoodMicro, Istanbul, Turkey, 3–7 September 2012. [Google Scholar]
- Frétin, M.; Martin, B.; Rifa, E.; Verdier-Metz, I.; Pomiès, D.; Ferlay, A.; Montel, M.C.; Delbès, C. Bacterial community assembly from cow teat skin to ripened cheeses is influenced by grazing systems. Sci. Rep. 2018, 8, 200. [Google Scholar] [CrossRef] [Green Version]
- Chemidlin Prévost-Bouré, N.; Karimi, B.; Sadet-Bourgeteau, S.; Djemiel, C.; Brie, M.; Dumont, J.; Campedelli, M.; Nowak, V.; Guyot, P.; Letourneur, C.; et al. Microbial transfers from permanent grassland ecosystems to milk in dairy farms in the Comté cheese area. Sci. Rep. 2021, 11, 18144. [Google Scholar] [CrossRef]
- Levante, A.; Bertani, G.; Bottari, B.; Bernini, V.; Lazzi, C.; Gatti, M.; Neviani, E. How new molecular approaches have contributed to shedding light on microbial dynamics in Parmigiano Reggiano cheese. Curr. Opin. Food Sci. 2021, 38, 131–140. [Google Scholar] [CrossRef]
- Tremblay, J.; Yergeau, E. Systematic processing of ribosomal RNA gene amplicon sequencing data. Gigascience 2019, 8, giz146. [Google Scholar] [CrossRef] [Green Version]
- Prodan, A.; Tremaroli, V.; Brolin, H.; Zwinderman, A.H.; Nieuwdorp, M.; Levin, E. Comparing bioinformatic pipelines for microbial 16S rRNA amplicon sequencing. PLoS ONE 2020, 15, e0227434. [Google Scholar] [CrossRef] [Green Version]
- Michailidou, S.; Pavlou, E.; Pasentsis, K.; Rhoades, J.; Likotrafiti, E.; Argiriou, A. Microbial profiles of Greek PDO cheeses assessed with amplicon metabarcoding. Food Microbiol. 2021, 99, 103836. [Google Scholar] [CrossRef]
- Zago, M.; Rossetti, L.; Bardelli, T.; Carminati, D.; Nazzicari, N.; Giraffa, G. Generical Hard Cheeses: DNA Metabarcoding and DNA Metafingerprinting Analysis to Assess Similarities and Differences. Foods 2021, 10, 1826. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef] [Green Version]
- Verdier-Metz, I.; Delbès, C.; Bouchon, M.; Pradel, P.; Theil, S.; Rifa, E.; Corbin, A.; Chassard, C. Influence of Post-Milking Treatment on Microbial Diversity on the Cow Teat Skin and in Milk. Dairy 2022, 3, 262–276. [Google Scholar] [CrossRef]
- Monsallier, F.; Verdier-Metz, I.; Agabriel, C.; Martin, B.; Montel, M.C. Variability of microbial teat skin flora in relation to farming practices and individual dairy cow characteristics. Dairy Sci. Technol. 2012, 92, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Verdier-Metz, I.; Gagne, G.; Bornes, S.; Monsallier, F.; Veisseire, P.; Delbès-Paus, C.; Montel, M.C. Cow Teat Skin, a Potential Source of Diverse Microbial Populations for Cheese Production. Appl. Environ. Microbiol. 2012, 78, 326–333. [Google Scholar] [CrossRef] [Green Version]
- Poirier, S.; Rué, O.; Coeuret, G.; Champomier-Vergès, M.C.; Loux, V.; Chaillou, S. Detection of an amplification bias associated to Leuconostocaceae family with a universal primer routinely used for monitoring microbial community structures within food products. BMC Res. Notes 2018, 11, 802. [Google Scholar] [CrossRef]
- Irobi, J.; Schoofs, A.; Goossens, H. Genetic identification of Candida species in HIV-positive patients using the polymerase chain reaction and restriction fragment length polymorphism analysis of its DNA. Mol. Cell. Probes 1999, 13, 401–406. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Mills, D.A. Improved selection of internal transcribed spacer-specific primers enables quantitative, ultra-high-throughput profiling of fungal communities. Appl. Environ. Microbiol. 2013, 79, 2519–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theil, S.; Rifa, E. rANOMALY: AmplicoN wOrkflow for Microbial community AnaLYsis. F1000Research 2021, 10, 7. [Google Scholar] [CrossRef] [PubMed]
- Meola, M.; Rifa, E.; Shani, N.; Delbès, C.; Berthoud, H.; Chassard, C. DAIRYdb: A manually curated gold standard reference database for improved taxonomy annotation of 16S rRNA gene sequences from dairy products. BMC Genomics 2019, 20, 560. [Google Scholar] [CrossRef] [Green Version]
- Theil, S.; Rifa, E. UTOPIA: An automatically UpdaTed, cOmPlete and consistent ITS reference dAtabase. In Proceedings of the Genomique Environnementale, Rochelle, France, 8–10 October 2019; Available online: https://hal.science/hal-02340446 (accessed on 20 January 2023).
- Tyrrell, H.F.; Reid, J.T. Prediction of the Energy Value of Cow’s Milk. J. Dairy Sci. 1965, 48, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Theil, S.; Rifa, E. Erifa1/ExploreMetabar: V1.0.0. Zenodo. Available online: https://zenodo.org/record/5245392(accessed on 20 January 2023).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Paulson, J.N.; Talukder, H.; Bravo, H.C. Longitudinal differential abundance analysis of microbial marker-gene surveys using smoothing splines. bioRxiv 2017. [Google Scholar] [CrossRef] [Green Version]
- Foster, Z.S.L.; Sharpton, T.J.; Grünwald, N.J. Metacoder: An R package for visualization and manipulation of community taxonomic diversity data. PLoS Comput. Biol. 2017, 13, e1005404. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix. (Version 0.92). 2021. Available online: https://github.com/taiyun/corrplot (accessed on 20 January 2023).
- de Ondarza, M.B.; Sniffen, C.J.; Dussert, L.; Chevaux, E.; Sullivan, J.; Walker, N. Case Study: Multiple-Study Analysis of the Effect of Live Yeast on Milk Yield, Milk Component Content and Yield, and Feed Efficiency. Prof. Anim. Sci. 2010, 26, 661–666. [Google Scholar] [CrossRef]
- Chaucheyras-Durand, F.; Ameilbonne, A.; Bichat, A.; Mosoni, P.; Ossa, F.; Forano, E. Live yeasts enhance fibre degradation in the cow rumen through an increase in plant substrate colonization by fibrolytic bacteria and fungi. J. Appl. Microbiol. 2016, 120, 560–570. [Google Scholar] [CrossRef] [Green Version]
- Chaucheyras-Durand, F.; Durand, H. Probiotics in animal nutrition and health. Benef. Microbes 2010, 1, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The rumen microbiome: A crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut Microbes 2019, 10, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Ji, S.; Suen, G.; Wang, F.; Li, S. The Rumen Bacterial Community in Dairy Cows Is Correlated to Production Traits During Freshening Period. Front. Microbiol. 2021, 12, 630605. [Google Scholar] [CrossRef] [PubMed]
- Doyle, C.J.; Gleeson, D.; O’Toole, P.W.; Cotter, P.D. Impacts of seasonal housing and teat preparation on raw milk microbiota: A high-throughput sequencing study. Appl. Environ. Microbiol. 2017, 83, e02694-16. [Google Scholar] [CrossRef] [Green Version]
- Mallet, A.; Guéguen, M.; Kauffmann, F.; Chesneau, C.; Sesboué, A.; Desmasures, N. Quantitative and qualitative microbial analysis of raw milk reveals substantial diversity in fl uenced by herd management practices. Int. Dairy J. 2012, 27, 13–21. [Google Scholar] [CrossRef]
- Marchand, S.; De Block, J.; De Jonghe, V.; Coorevits, A.; Heyndrickx, M.; Herman, L.; De Block, J.; De Jonghe, V.; Coorevits, A.; Heyndrickx, M.; et al. Biofilm Formation in Milk Production and Processing Environments; Influence on Milk Quality and Safety. Compr. Rev. Food Sci. Food Saf. 2012, 11, 133–147. [Google Scholar] [CrossRef]
- Fernandes, T.; Carvalho, B.F.; Mantovani, H.C.; Schwan, R.F.; Ávila, C.L.S. Identification and characterization of yeasts from bovine rumen for potential use as probiotics. J. Appl. Microbiol. 2019, 127, 845–855. [Google Scholar] [CrossRef]
- Lamarche, A.; Lessard, M.H.; Viel, C.; Turgeon, S.L.; St-Gelais, D.; Labrie, S. Quantitative PCR reveals the frequency and distribution of 3 indigenous yeast species across a range of specialty cheeses. J. Dairy Sci. 2022, 105, 8677–8687. [Google Scholar] [CrossRef]
- Ji, Y.; Dong, X.; Liu, Z.; Wang, W.; Yan, H.; Liu, X. Effects of Bovine Pichia kudriavzevii T7, Candida glabrata B14, and Lactobacillus plantarum Y9 on Milk Production, Quality and Digestive Tract Microbiome in Dairy Cows. Microorganisms 2022, 10, 842. [Google Scholar] [CrossRef]
- Bekada, A.M.A.; Benakriche, B.; Hamadi, K.; Bensoltane, A. Modelling of Effects of Water Activity, pH and Temperature on the Growth Rate of Mucor racemosus Isolated from Soft Camembert Cheese. World J. Agric. Sci. 2008, 4, 790–794. [Google Scholar]
- Cox, M.S.; Deblois, C.L.; Suen, G. Assessing the Response of Ruminal Bacterial and Fungal Microbiota to Whole-Rumen Contents Exchange in Dairy Cows. Front. Microbiol. 2021, 12, 665776. [Google Scholar] [CrossRef]
- Fleet, G.H. Yeasts in dairy products. J. Dairy Bacteriol. 1990, 68, 199–211. [Google Scholar] [CrossRef]
- Garnier, L.; Valence, F.; Mounier, J. Diversity and Control of Spoilage Fungi in Dairy Products: An Update. Microorganisms 2017, 5, 42. [Google Scholar] [CrossRef] [Green Version]
- Wuytack, A.; De Visscher, A.; Piepers, S.; Boyen, F.; Haesebrouck, F.; De Vliegher, S. Distribution of non-aureus staphylococci from quarter milk, teat apices, and rectal feces of dairy cows, and their virulence potential. J. Dairy Sci. 2020, 103, 10658–10675. [Google Scholar] [CrossRef]
- Gelsomino, R.; Vancanneyt, M.; Cogan, T.M.; Condon, S.; Swings, J. Source of enterococci in a farmhouse raw-milk cheese. Appl. Environ. Microbiol. 2002, 68, 3560–3565. [Google Scholar] [CrossRef] [Green Version]
- Besnard, A.; Desmasures, N.; Voisin-Anastasie, A.; Gréau, L.; Lelièvre, V.; Bré, J.M.; Dalmasso, M.; Schlusselhuber, M. Aerococcus sp., a promising genus as a source of anti-Salmonella bioprotective agents for the dairy industry revealed by a miniaturised screening method. Int. Dairy J. 2021, 116, 104949. [Google Scholar] [CrossRef]
- Dean, C.J.; Slizovskiy, I.B.; Crone, K.K.; Pfennig, A.X.; Heins, B.J.; Caixeta, L.S.; Noyes, N.R. Investigating the cow skin and teat canal microbiomes of the bovine udder using different sampling and sequencing approaches. J. Dairy Sci. 2021, 104, 644–661. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | LY | SEM | LY Effect * |
|---|---|---|---|---|
| Dry matter intake (DMI, kg/d) | 20.62 | 20.99 | 0.30 | 0.397 |
| Feed Conversion Ratio (FCR) | 1.117 | 1.088 | 0.024 | 0.391 |
| Milk yield (kg/d) | 19.17 | 19.82 | 0.27 | 0.098 |
| Energy corrected milk (ECM, kg/d) | 19.69 | 20.28 | 0.35 | 0.246 |
| Fat content (g/kg) | 37.06 | 37.03 | 0.39 | 0.954 |
| Protein content (g/kg) | 30.50 | 30.51 | 0.21 | 0.969 |
| Milk Fat Yield (kg/d) | 0.702 | 0.731 | 0.01 | 0.098 |
| Milk Protein Yield (kg/d) | 0.583 | 0.602 | 0.01 | 0.287 |
| Casein (mg/kg) | 274.3 | 278.7 | 1.6 | 0.058 |
| Lactose (g/kg) | 49.5 | 50.8 | 0.2 | <0.01 |
| Urea (mg/L) | 312.4 | 313.3 | 7.9 | 0.942 |
| Somatic Cell Count (Log10/mL) | 5.08 | 5.00 | 0.06 | 0.352 |
| Body Weight start (BW, kg) | 635.3 | 648.0 | 5.2 | 0.087 |
| Body Weight end trial (BW, kg) | 636.2 | 667.6 | 13.5 | 0.114 |
| Milk | Teat | Bedding Material | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Microbes Level | Taxa | Methods1 | C | LY | Sign.2 | Methods1 | C | LY | Sign.2 | Methods1 | C | LY | Sign.2 | ||
| n = 38 | n = 40 | n = 18 | n = 20 | n = 3 | n = 3 | ||||||||||
| Fungi | |||||||||||||||
| ASV | Candida__railenensis | - | - | - | 3 | 0 | 1.61 | * | - | - | - | ||||
| Cystobasidium_laryngis | - | - | - | - | - | - | 1 | 4.84 | 1.01 | * | |||||
| Cystobasidium_pallidum | - | - | - | - | - | - | 2 | 0.17 | 1.47 | * | |||||
| Mucor_racemosus | - | - | - | 3 | 0.24 | 2.95 | * | - | - | - | |||||
| Pichia_fermentans | 1 | 1.59 | 0.03 | * | - | - | - | - | - | - | |||||
| Pichia_kudriavzevii | - | - | - | 1, 3 | 0.03 | 10.6 | * | 1, 2 | 0.02 | 53.09 | * | ||||
| Saturnispora_silvae | - | - | - | 2 | 2.3 | 0.46 | * | 1 | 8.36 | 1.52 | * | ||||
| Saturnispora_species | - | - | - | - | - | - | 1 | 2.05 | 0.33 | * | |||||
| Species | Acrostalagmus_species | 3 | 1.1 | 0 | + | 3 | 1.83 | 0.34 | + | - | - | - | |||
| Aspergillus_species | 3 | 1.31 | 2.94 | + | - | - | - | - | - | - | |||||
| Bettsia_alvei | - | - | - | 3 | 0.4 | 4.6 | + | - | - | - | |||||
| Candida__railenensis | - | - | - | 3 | 0 | 1.9 | * | - | - | - | |||||
| Claviceps_purpurea | - | - | - | - | - | - | 1 | 1.62 | 0.37 | * | |||||
| Cystobasidium_laryngis | - | - | - | - | - | - | 1 | 5.16 | 1.01 | * | |||||
| Cystobasidium_pallidum | - | - | - | - | - | - | 1 | 0.81 | 3.25 | * | |||||
| Debaryomyces_hansenii | - | - | - | - | - | - | 1 | 1.26 | 0.23 | * | |||||
| Mucor_racemosus | - | - | - | 3 | 0.24 | 3.45 | * | - | - | - | |||||
| Neocallimastigaceae_sp | - | - | - | - | - | - | 1 | 2.24 | 0.32 | * | |||||
| Pichia_kudriavzevii | - | - | - | 1, 3 | 0.05 | 11.33 | * | 1, 2 | 0.02 | 53.09 | * | ||||
| Saturnispora_species | - | - | - | 2 | 9.45 | 1.1 | * | 1 | 13.44 | 2.31 | * | ||||
| Wickerhamomyces_anomalus | - | - | - | 1, 3 | 21.27 | 6.14 | * | - | - | - | |||||
| Genus | Acrostalagmus | 3 | 1.1 | 0 | + | - | - | - | 3 | 1.98 | 0.98 | + | |||
| Aspergillus | 3 | 1.31 | 5.87 | * | - | - | - | 1 | 1.6 | 1.76 | * | ||||
| Cladosporium | - | - | - | - | - | - | 3 | 3.09 | 2.08 | + | |||||
| Claviceps | - | - | - | - | - | - | 3 | 2.95 | 1.67 | + | |||||
| Cystobasidium | - | - | - | - | - | - | 3 | 6.15 | 4.26 | + | |||||
| Debaryomyces | - | - | - | - | - | - | 1 | 1.26 | 0.23 | * | |||||
| Galactomyces | - | - | - | - | - | - | 3 | 44.06 | 17.15 | + | |||||
| Kurtzmaniella | - | - | - | 1, 3 | 0 | 1.9 | * | - | - | - | |||||
| Mucor | - | - | - | - | - | - | 1, 2 | 0.29 | 1.53 | * | |||||
| Neocallimastigaceae_genus | - | - | - | - | - | - | 3 | 2.36 | 0.85 | + | |||||
| Penicillium | - | - | - | - | - | - | 3 | 1.16 | 0.51 | + | |||||
| Pichia | - | - | - | - | - | - | 1, 2 | 4.99 | 55.56 | * | |||||
| Saturnispora | - | - | - | 2 | 9.45 | 1.1 | * | 1 | 13.44 | 2.31 | * | ||||
| Wickerhamomyces | - | - | - | 1, 3 | 21.27 | 6.14 | * | - | - | - | |||||
| Bacteria | |||||||||||||||
| ASV | Aerococcus_suis | 3 | 0.33 | 1.24 | * | 3 | 1.13 | 4.41 | * | - | - | - | |||
| Staphylococcus_hominis | 1 | 0.08 | 1.41 | * | 3 | 0.03 | 2.86 | * | - | - | - | ||||
| Species | Acinetobacter_johnsonii | 3 | 0.06 | 1.83 | * | - | - | - | - | - | - | ||||
| Acinetobacter_species | - | - | - | 1 | 8.57 | 1.6 | * | - | - | - | |||||
| Pseudomonas_alcaliphila | 3 | 1.63 | 1.03 | * | - | - | - | - | - | - | |||||
| Staphylococcus_species | - | - | - | 1, 3 | 1.6 | 7.24 | * | - | - | - | |||||
| Vibrionimonas_magnilacihabitans | 3 | 1.63 | 0.23 | * | - | - | - | - | - | - | |||||
| Genus | Enterococcus | 1 | 0.37 | 5.34 | * | - | - | - | - | - | - | ||||
| Jeotgalicoccus | - | - | - | 3 | 2.73 | 1.96 | * | - | - | - | |||||
| Staphylococcus | - | - | - | 1, 3 | 2.29 | 9.98 | * | - | - | - | |||||
| Vibrionimonas | 3 | 1.65 | 0.24 | * | - | - | - | - | - | - | |||||
| C 1 | LY 1 | p-Value 3 | ||||||
|---|---|---|---|---|---|---|---|---|
| P1 2 | P2 2 | P3 2 | P1 2 | P2 2 | P3 2 | Group | Period | |
| Fungi | ||||||||
| Number of cows | 9 | 9 | 6 | 10 | 10 | 6 | ||
| Mean number of teat ASVs total | 46 | 44 | 40 | 36 | 42 | 44 | ns | ns |
| Mean number of milk ASVs total | 13 | 16 | 9 | 12 | 18 | 11 | ns | + |
| Mean number of ASVs shared by individual milk and teat | 4 | 7 | 5 | 4 | 6 | 6 | ns | ns |
| Mean fraction of milk ASVs shared with teat (%) | 37 | 44 | 51 | 33 | 34 | 65 | ns | ** |
| Mean cumulative abundance in milk of shared ASVs (%) | 35 | 68 | 65 | 36 | 28 | 47 | ns | + |
| Bacteria | ||||||||
| Number of cows | 9 | 9 | 9 | 10 | 9 | 9 | ||
| Mean number of teat ASVs total | 113 | 107 | 70 | 104 | 103 | 74 | ns | *** |
| Mean number of milk ASVs total | 70 | 63 | 52 | 74 | 65 | 54 | ns | ** |
| Mean number of ASVs shared by individual milk and teat | 47 | 34 | 25 | 47 | 33 | 27 | ns | *** |
| Mean fraction of milk ASVs shared with teat (%) | 67 | 54 | 44 | 64 | 50 | 50 | ns | *** |
| Mean cumulative abundance in milk of shared ASVs (%) | 89 | 54 | 57 | 82 | 71 | 74 | ns | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verdier-Metz, I.; Delbès, C.; Bouchon, M.; Rifa, E.; Theil, S.; Chaucheyras-Durand, F.; Chevaux, E.; Dunière, L.; Chassard, C. Dietary Live Yeast Supplementation Influence on Cow’s Milk, Teat and Bedding Microbiota in a Grass-Diet Dairy System. Microorganisms 2023, 11, 673. https://doi.org/10.3390/microorganisms11030673
Verdier-Metz I, Delbès C, Bouchon M, Rifa E, Theil S, Chaucheyras-Durand F, Chevaux E, Dunière L, Chassard C. Dietary Live Yeast Supplementation Influence on Cow’s Milk, Teat and Bedding Microbiota in a Grass-Diet Dairy System. Microorganisms. 2023; 11(3):673. https://doi.org/10.3390/microorganisms11030673
Chicago/Turabian StyleVerdier-Metz, Isabelle, Céline Delbès, Matthieu Bouchon, Etienne Rifa, Sébastien Theil, Frédérique Chaucheyras-Durand, Eric Chevaux, Lysiane Dunière, and Christophe Chassard. 2023. "Dietary Live Yeast Supplementation Influence on Cow’s Milk, Teat and Bedding Microbiota in a Grass-Diet Dairy System" Microorganisms 11, no. 3: 673. https://doi.org/10.3390/microorganisms11030673