
Antibodies to Combat Fungal Infections: Development Strategies and Progress
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
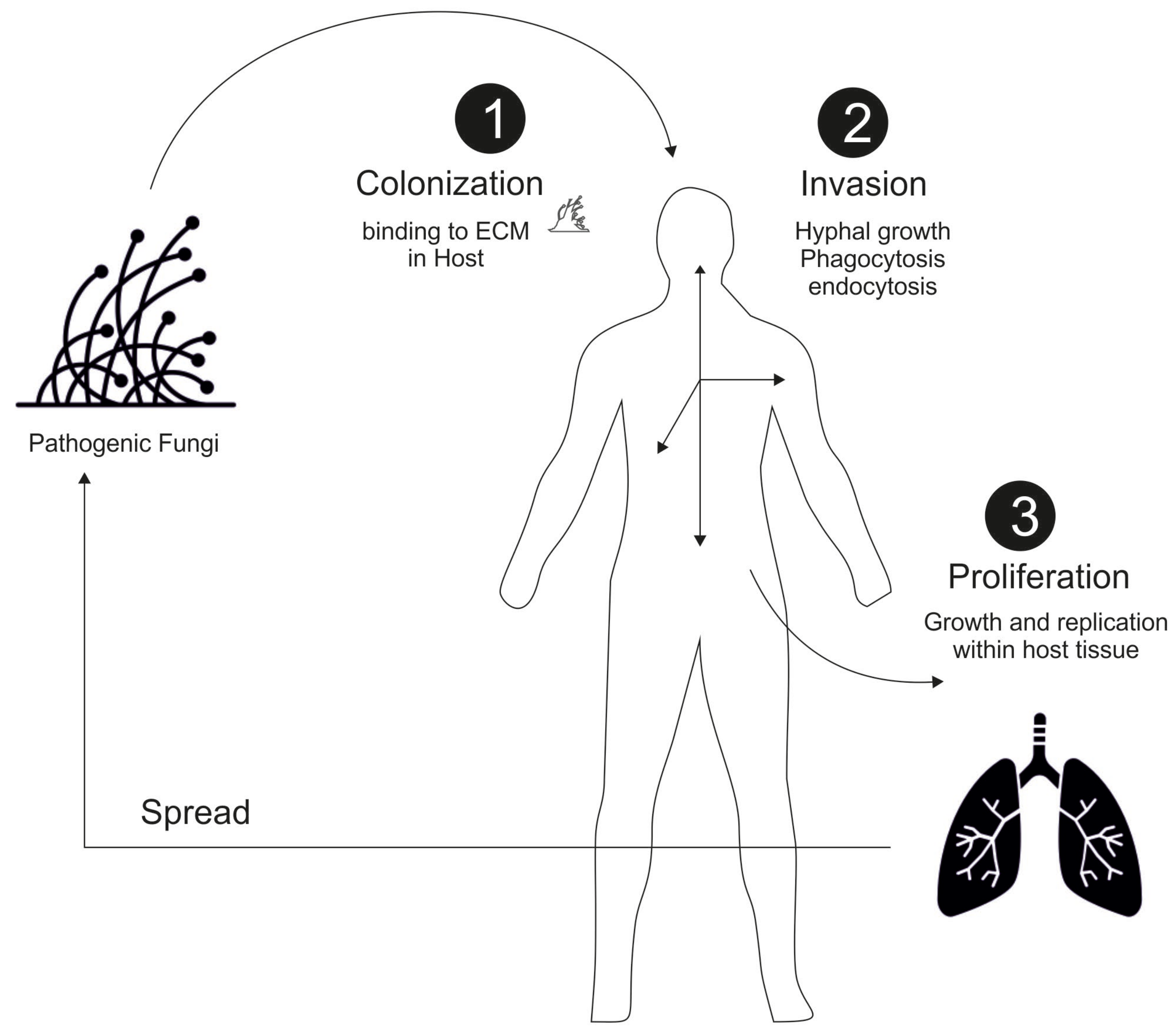
2. General Mechanism of Fungal Infection
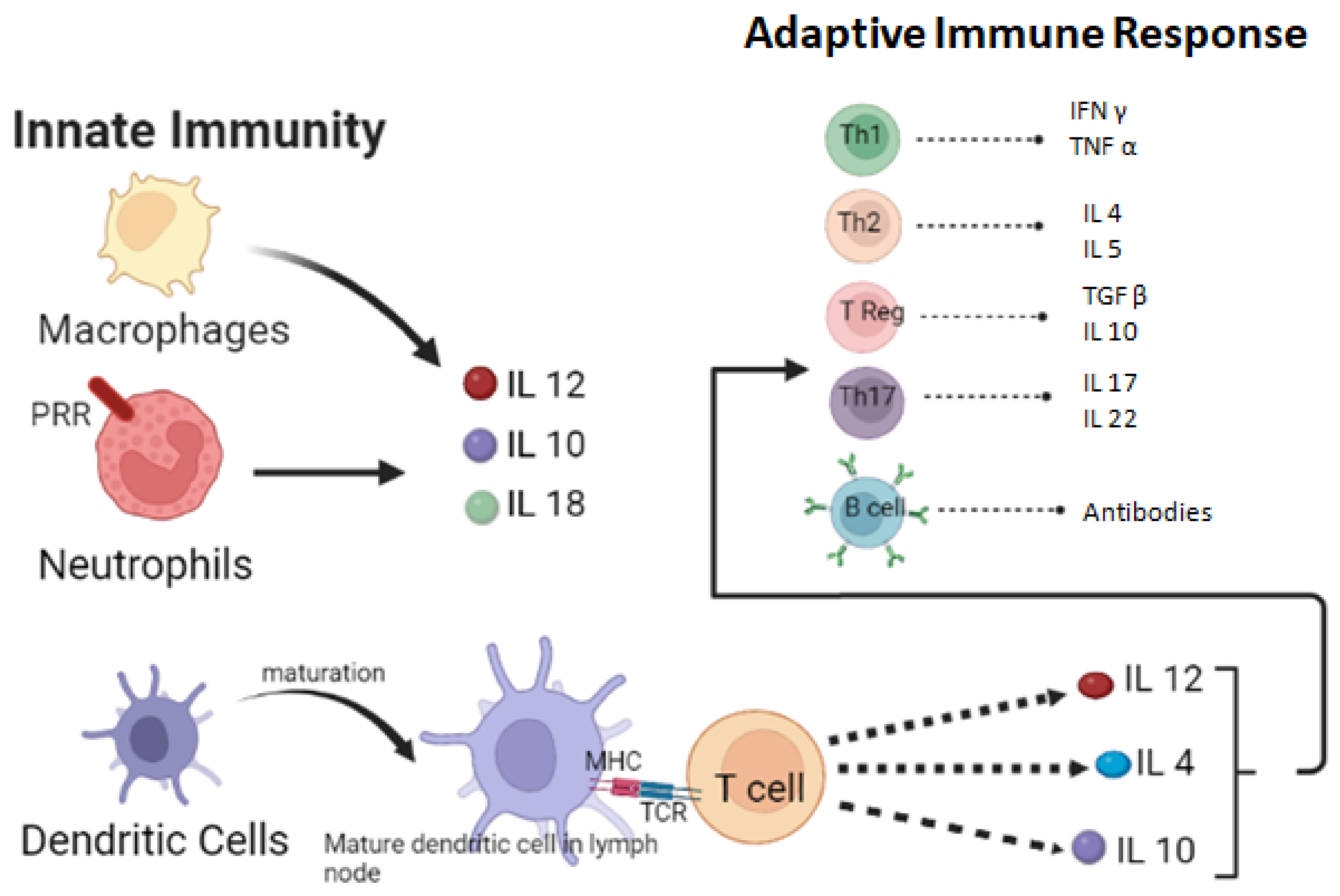
3. Host Defense Mechanism against Fungi
4. Cell-Mediated Response
5. Humoral Immune Response
6. Antibodies against Fungal Infection
- (a) Should be a broad spectrum for a variety of fungi;
- (b) Should be fungicidal as opposed to fungistatic;
- (c) Should be against specific fungal target region;
- (d) Should have spare interference with host targets;
- (e) Should have minimal side effects or toxicities.
7. Future Prospects
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cairns, T.C.; Nai, C.; Meyer, V. How a fungus shapes biotechnology: 100 years of Aspergillus niger research. Fungal Biol. Biotechnol. 2018, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Spagnuolo, M.; Yaguchi, A.; Blenner, M. Oleaginous yeast for biofuel and oleochemical production. Curr. Opin. Biotechnol. 2019, 57, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Chang, R. Bioactive polysaccharides from traditional Chinese medicine herbs as anticancer adjuvants. J. Altern. Complement. Med. 2002, 8, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Rop, O.; Mlcek, J.; Jurikova, T. Beta-glucans in higher fungi and their health effects. Nutr. Rev. 2009, 67, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Pelley, R.P.; Strickland, F.M. Plants, polysaccharides, and the treatment and prevention of neoplasia. Crit. Rev. ™ Oncog. 2000, 11, 189–225. [Google Scholar] [CrossRef]
- Leal, F. Fungal Pathogenesis in Humans: The Growing Threat. Genes 2019, 10, 136. [Google Scholar] [CrossRef]
- Van Burik, J.-A.H.; Magee, P.T. Aspects of fungal pathogenesis in humans. Annu. Rev. Microbiol. 2001, 55, 743–772. [Google Scholar] [CrossRef]
- Bitew, A. Dermatophytosis: Prevalence of dermatophytes and non-dermatophyte fungi from patients attending Arsho advanced medical laboratory, Addis Ababa, Ethiopia. Dermatol. Res. Pract. 2018, 2018, 8164757. [Google Scholar] [CrossRef]
- Khan, H.A.; Baig, F.K.; Mehboob, R. Nosocomial infections: Epidemiology, prevention, control and surveillance. Asian Pac. J. Trop. Biomed. 2017, 7, 478–482. [Google Scholar] [CrossRef]
- Gnat, S.; Łagowski, D.; Nowakiewicz, A.; Dyląg, M. A global view on fungal infections in humans and animals: Opportunistic infections and microsporidioses. J. Appl. Microbiol. 2021, 131, 2095–2113. [Google Scholar] [CrossRef]
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and multi-national prevalence of fungal diseases—Estimate precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef]
- Benedict, K.; Jackson, B.R.; Chiller, T.; Beer, K.D. Estimation of direct healthcare costs of fungal diseases in the United States. Clin. Infect. Dis. 2019, 68, 1791–1797. [Google Scholar] [CrossRef]
- Almeida, F.; Rodrigues, M.L.; Coelho, C. The still underestimated problem of fungal diseases worldwide. Front. Microbiol. 2019, 10, 214. [Google Scholar] [CrossRef]
- Darwin, J. Improving Outcomes for Patients with Fungal Infections across the World a Road Map for the Next Decade; Global Action Fund for Fungal Infections: Geneva, Switzerland, 2015. [Google Scholar]
- Köhler, J.R.; Casadevall, A.; Perfect, J. The spectrum of fungi that infects humans. Cold Spring Harb. Perspect. Med. 2015, 5, a019273. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J.; Fraser, J.A.; Doering, T.L.; Wang, Z.A.; Janbon, G.; Idnurm, A.; Bahn, Y.-S. Cryptococcus neoformans and Cryptococcus gattii, the etiologic agents of cryptococcosis. Cold Spring Harb. Perspect. Med. 2014, 4, a019760. [Google Scholar] [CrossRef]
- Limper, A.H.; Adenis, A.; Le, T.; Harrison, T.S. Fungal infections in HIV/AIDS. Lancet Infect. Dis. 2017, 17, e334–e343. [Google Scholar] [CrossRef]
- Sipsas, N.V.; Kontoyiannis, D.P. Invasive fungal infections in patients with cancer in the Intensive Care Unit. Int. J. Antimicrob. Agents 2012, 39, 464–471. [Google Scholar] [CrossRef]
- Martinez, R. New trends in paracoccidioidomycosis epidemiology. J. Fungi 2017, 3, 1. [Google Scholar] [CrossRef]
- Pachl, J.; Svoboda, P.; Jacobs, F.; Vandewoude, K.; Van Der Hoven, B.; Spronk, P.; Masterson, G.; Malbrain, M.; Aoun, M.; Garbino, J. A randomized, blinded, multicenter trial of lipid-associated amphotericin B alone versus in combination with an antibody-based inhibitor of heat shock protein 90 in patients with invasive candidiasis. Clin. Infect. Dis. 2006, 42, 1404–1413. [Google Scholar] [CrossRef]
- Wharton, R.E.; Stefanov, E.K.; King, R.G.; Kearney, J.F. Antibodies generated against Streptococci protect in a mouse model of disseminated aspergillosis. J. Immunol. 2015, 194, 4387–4396. [Google Scholar] [CrossRef]
- Strickland, A.B.; Shi, M. Mechanisms of fungal dissemination. Cell. Mol. Life Sci. 2021, 78, 3219–3238. [Google Scholar] [CrossRef] [PubMed]
- Moyes, D.L.; Richardson, J.P.; Naglik, J.R. Candida albicans-epithelial interactions and pathogenicity mechanisms: Scratching the surface. Virulence 2015, 6, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Dagenais, T.R.; Keller, N.P. Pathogenesis of Aspergillus fumigatus in invasive aspergillosis. Clin. Microbiol. Rev. 2009, 22, 447–465. [Google Scholar] [CrossRef] [PubMed]
- Mittal, J.; Ponce, M.G.; Gendlina, I.; Nosanchuk, J.D. Histoplasma capsulatum: Mechanisms for pathogenesis. Fungal Physiol. Immunopathogenesis 2019, 422, 157–191. [Google Scholar]
- Richardson, J.P.; Moyes, D.L. Adaptive immune responses to Candida albicans infection. Virulence 2015, 6, 327–337. [Google Scholar] [CrossRef]
- Romani, L. Immunity to fungal infections. Nat. Rev. Immunol. 2004, 4, 11–24. [Google Scholar] [CrossRef]
- Heninger, E.; Hogan, L.H.; Karman, J.; Macvilay, S.; Hill, B.; Woods, J.P.; Sandor, M. Characterization of the Histoplasma capsulatum-induced granuloma. J. Immunol. 2006, 177, 3303–3313. [Google Scholar] [CrossRef]
- Shibuya, K.; Hirata, A.; Omuta, J.; Sugamata, M.; Katori, S.; Saito, N.; Murata, N.; Morita, A.; Takahashi, K.; Hasegawa, C. Granuloma and cryptococcosis. J. Infect. Chemother. 2005, 11, 115–122. [Google Scholar] [CrossRef]
- Netea, M.G.; Gijzen, K.; Coolen, N.; Verschueren, I.; Figdor, C.; Van der Meer, J.W.; Torensma, R.; Kullberg, B.J. Human dendritic cells are less potent at killing Candida albicans than both monocytes and macrophages. Microbes Infect. 2004, 6, 985–989. [Google Scholar] [CrossRef]
- Ramirez-Ortiz, Z.G.; Lee, C.K.; Wang, J.P.; Boon, L.; Specht, C.A.; Levitz, S.M. A nonredundant role for plasmacytoid dendritic cells in host defense against the human fungal pathogen Aspergillus fumigatus. Cell Host Microbe 2011, 9, 415–424. [Google Scholar] [CrossRef]
- Ziegler-Heitbrock, L.; Ancuta, P.; Crowe, S.; Dalod, M.; Grau, V.; Hart, D.N.; Leenen, P.J.; Liu, Y.-J.; MacPherson, G.; Randolph, G.J. Nomenclature of monocytes and dendritic cells in blood. Blood J. Am. Soc. Hematol. 2010, 116, e74–e80. [Google Scholar] [CrossRef]
- Smeekens, S.P.; van de Veerdonk, F.L.; Joosten, L.A.; Jacobs, L.; Jansen, T.; Williams, D.L.; van der Meer, J.W.; Kullberg, B.J.; Netea, M.G. The classical CD14++ CD16− monocytes, but not the patrolling CD14+ CD16+ monocytes, promote Th17 responses to Candida albicans. Eur. J. Immunol. 2011, 41, 2915–2924. [Google Scholar] [CrossRef]
- Campbell, K.S.; Hasegawa, J. Natural killer cell biology: An update and future directions. J. Allergy Clin. Immunol. 2013, 132, 536–544. [Google Scholar] [CrossRef]
- Voigt, J.; Hünniger, K.; Bouzani, M.; Jacobsen, I.D.; Barz, D.; Hube, B.; Löffler, J.; Kurzai, O. Human natural killer cells acting as phagocytes against Candida albicans and mounting an inflammatory response that modulates neutrophil antifungal activity. J. Infect. Dis. 2014, 209, 616–626. [Google Scholar] [CrossRef]
- Schmidt, S.; Tramsen, L.; Hanisch, M.; Latgé, J.-P.; Huenecke, S.; Koehl, U.; Lehrnbecher, T. Human natural killer cells exhibit direct activity against Aspergillus fumigatus hyphae, but not against resting conidia. J. Infect. Dis. 2011, 203, 430–435. [Google Scholar] [CrossRef]
- Nakamatsu, M.; Yamamoto, N.; Hatta, M.; Nakasone, C.; Kinjo, T.; Miyagi, K.; Uezu, K.; Nakamura, K.; Nakayama, T.; Taniguchi, M. Role of interferon-γ in Vα14+ natural killer T cell-mediated host defense against Streptococcus pneumoniae infection in murine lungs. Microbes Infect. 2007, 9, 364–374. [Google Scholar] [CrossRef]
- Cohen, N.R.; Tatituri, R.V.; Rivera, A.; Watts, G.F.; Kim, E.Y.; Chiba, A.; Fuchs, B.B.; Mylonakis, E.; Besra, G.S.; Levitz, S.M. Innate recognition of cell wall β-glucans drives invariant natural killer T cell responses against fungi. Cell Host Microbe 2011, 10, 437–450. [Google Scholar] [CrossRef]
- Horn, D.L.; Neofytos, D.; Anaissie, E.J.; Fishman, J.A.; Steinbach, W.J.; Olyaei, A.J.; Marr, K.A.; Pfaller, M.A.; Chang, C.-H.; Webster, K.M. Epidemiology and outcomes of candidemia in 2019 patients: Data from the prospective antifungal therapy alliance registry. Clin. Infect. Dis. 2009, 48, 1695–1703. [Google Scholar] [CrossRef]
- Feldmesser, M. Prospects of vaccines for medically important fungi. Sabouraudia 2005, 43, 571–587. [Google Scholar] [CrossRef]
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-derived suppressor cells as regulators of the immune system. Nat. Rev. Immunol. 2009, 9, 162–174. [Google Scholar] [CrossRef]
- Hayes, B.M.; Bleackley, M.R.; Wiltshire, J.L.; Anderson, M.A.; Traven, A.; van der Weerden, N.L. Identification and mechanism of action of the plant defensin NaD1 as a new member of the antifungal drug arsenal against Candida albicans. Antimicrob. Agents Chemother. 2013, 57, 3667–3675. [Google Scholar] [CrossRef] [PubMed]
- LeibundGut-Landmann, S.; Wüthrich, M.; Hohl, T.M. Immunity to fungi. Curr. Opin. Immunol. 2012, 24, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Lindell, D.M.; Moore, T.A.; McDonald, R.A.; Toews, G.B.; Huffnagle, G.B. Generation of antifungal effector CD8+ T cells in the absence of CD4+ T cells during Cryptococcus neoformans infection. J. Immunol. 2005, 174, 7920–7928. [Google Scholar] [CrossRef] [PubMed]
- Nanjappa, S.G.; Heninger, E.; Wüthrich, M.; Gasper, D.J.; Klein, B.S. Tc17 cells mediate vaccine immunity against lethal fungal pneumonia in immune deficient hosts lacking CD4+ T cells. PLoS Pathog. 2012, 8, e1002771. [Google Scholar] [CrossRef] [PubMed]
- Berland, R.; Wortis, H.H. Origins and Functions of B-1 Cells. Annu. Rev. Immunol 2002, 20, 253–300. [Google Scholar] [CrossRef]
- Hardy, R.R. B-1 B cells: Development, selection, natural autoantibody and leukemia. Curr. Opin. Immunol. 2006, 18, 547–555. [Google Scholar] [CrossRef]
- Kohler, H.; Bayry, J.; Nicoletti, A.; Kaveri, S. Natural autoantibodies as tools to predict the outcome of immune response? Scand. J. Immunol. 2003, 58, 285–289. [Google Scholar] [CrossRef]
- Zhou, Z.-h.; Wild, T.; Xiong, Y.; Sylvers, L.H.; Zhang, Y.; Zhang, L.; Wahl, L.; Wahl, S.M.; Kozlowski, S.; Notkins, A.L. Polyreactive antibodies plus complement enhance the phagocytosis of cells made apoptotic by UV-light or HIV. Sci. Rep. 2013, 3, 2271. [Google Scholar] [CrossRef]
- Notkins, A.L. Polyreactivity of antibody molecules. Trends Immunol. 2004, 25, 174–179. [Google Scholar] [CrossRef]
- Ehrenstein, M.R.; Notley, C.A. The importance of natural IgM: Scavenger, protector and regulator. Nat. Rev. Immunol. 2010, 10, 778–786. [Google Scholar] [CrossRef]
- Coutinho, A.; Kazatchkine, M.D.; Avrameas, S. Natural autoantibodies. Curr. Opin. Immunol. 1995, 7, 812–818. [Google Scholar] [CrossRef]
- López-Ribot, J.L.; Casanova, M.; Murgui, A.; Martínez, J.P. Antibody response to Candida albicans cell wall antigens. FEMS Immunol. Med. Microbiol. 2004, 41, 187–196. [Google Scholar] [CrossRef]
- Kuruganti, U.; Henderson, L.A.; Garner, R.E.; Asofsky, R.; Baker, P.J.; Domer, J.E. Nonspecific and Candida-Specific Immune Responses in Mice Suppressed by Chronic Administration of Anti-μ. J. Leukoc. Biol. 1988, 44, 422–433. [Google Scholar] [CrossRef]
- Han, Y.; Morrison, R.P.; Cutler, J.E. A vaccine and monoclonal antibodies that enhance mouse resistance to Candida albicans vaginal infection. Infect. Immun. 1998, 66, 5771–5776. [Google Scholar] [CrossRef]
- Mukherjee, J.; Scharff, M.D.; Casadevall, A. Protective murine monoclonal antibodies to Cryptococcus neoformans. Infect. Immun. 1992, 60, 4534–4541. [Google Scholar] [CrossRef]
- Montagnoli, C.; Bozza, S.; Bacci, A.; Gaziano, R.; Mosci, P.; Morschhäuser, J.; Pitzurra, L.; Kopf, M.; Cutler, J.; Romani, L. A role for antibodies in the generation of memory antifungal immunity. Eur. J. Immunol. 2003, 33, 1193–1204. [Google Scholar] [CrossRef]
- Li, W.; Fu, M.; An, J.G.; Xing, Y.; Zhang, P.; Zhang, X.; Wang, Y.C.; Li, C.X.; Tian, R.; Su, W.J. Host defence against C. albicans infections in IgH transgenic mice with VH derived from a natural anti-keratin antibody. Cell. Microbiol. 2007, 9, 306–315. [Google Scholar] [CrossRef]
- Tian, R.; Fu, M.; Zhang, Z.; Ren, J.; An, J.; Liu, Y.; Li, W. In situ IgM production and clonal expansion of B-1 cells in peritoneal cavity promote elimination of C. albicans infection in IgH transgenic mice with VH derived from a natural antibody. PLoS ONE 2013, 8, e60779. [Google Scholar] [CrossRef]
- Zaragoza, O.; Taborda, C.P.; Casadevall, A. The efficacy of complement-mediated phagocytosis of Cryptococcus neoformans is dependent on the location of C3 in the polysaccharide capsule and involves both direct and indirectC3-mediated interactions. Eur. J. Immunol. 2003, 33, 1957–1967. [Google Scholar] [CrossRef]
- Shapiro, S.; Beenhouwer, D.O.; Feldmesser, M.; Taborda, C.; Carroll, M.C.; Casadevall, A.; Scharff, M.D. Immunoglobulin G monoclonal antibodies to Cryptococcus neoformans protect mice deficient in complement component C3. Infect. Immun. 2002, 70, 2598–2604. [Google Scholar] [CrossRef]
- Casadevall, A.; Pirofski, L.A. Immunoglobulins in defense, pathogenesis, and therapy of fungal diseases. Cell Host Microbe 2012, 11, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Van Spriel, A.B.; Sofi, M.; Gartlan, K.H.; van der Schaaf, A.; Verschueren, I.; Torensma, R.; Raymakers, R.A.; Loveland, B.E.; Netea, M.G.; Adema, G.J. The tetraspanin protein CD37 regulates IgA responses and anti-fungal immunity. PLoS Pathog. 2009, 5, e1000338. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.F.; Mitchell, T.G.; Storkus, W.; Dawson, J.R. Human natural killer cells do not inhibit growth of Cryptococcus neoformans in the absence of antibody. Infect. Immun. 1990, 58, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, N.; Murphy, J.W. Antibody-dependent natural killer cell-mediated growth inhibition of Cryptococcus neoformans. Infect. Immun. 1986, 51, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Magliani, W.; Conti, S.; Frazzi, R.; Ravanetti, L.; Maffei, D.; Polonelli, L. Protective antifungal yeast killer toxin-like antibodies. Curr. Mol. Med. 2005, 5, 443–452. [Google Scholar] [CrossRef]
- Kainz, K.; Bauer, M.A.; Madeo, F.; Carmona-Gutierrez, D. Fungal infections in humans: The silent crisis. Microb. Cell 2020, 7, 143. [Google Scholar] [CrossRef]
- Marcos, C.M.; De Oliveira, H.C.; de Melo, W.d.C.M.A.; Da Silva, J.D.F.; Assato, P.A.; Scorzoni, L.; Rossi, S.A.; de Paula e Silva, A.C.; Mendes-Giannini, M.J.; Fusco-Almeida, A.M. Anti-immune strategies of pathogenic fungi. Front. Cell. Infect. Microbiol. 2016, 6, 142. [Google Scholar] [CrossRef]
- Patin, E.C.; Thompson, A.; Orr, S.J. Pattern recognition receptors in fungal immunity. Semin. Cell Dev. Biol. 2019, 89, 24–33. [Google Scholar] [CrossRef]
- Gupta, S.; Ellis, M.; Cesario, T.; Ruhling, M.; Vayuvegula, B. Disseminated cryptococcal infection in a patient with hypogammaglobulinemia and normal T cell functions. Am. J. Med. 1987, 82, 129–131. [Google Scholar] [CrossRef]
- Wahab, J.; Hanifah, M.; Choo, K. Bruton’s agamma globulinaemia in a child presenting with cryptococcal, empyemia thoracis and periauricular pyogenic abscess. Singap. Med. J. 1995, 36, 686–689. [Google Scholar]
- Pires, N.; Guimarães, M.C.; Moya, M.J.; Oliveira, F.R.; Louzada, J.; Martinez, R. Hypogammaglobulinemia as predisposing factor for Cryptococcus neoformans infection: Report of two cases. Rev. Da Soc. Bras. De Med. Trop. 2000, 33, 603–608. [Google Scholar]
- Agematsu, K.; Nagumo, H.; Shinozaki, K.; Hokibara, S.; Yasui, K.; Terada, K.; Kawamura, N.; Toba, T.; Nonoyama, S.; Ochs, H.D. Absence of IgD-CD27 (+) memory B cell population in X-linked hyper-IgM syndrome. J. Clin. Investig. 1998, 102, 853–860. [Google Scholar] [CrossRef]
- Marr, K.A.; Datta, K.; Pirofski, L.-a.; Barnes, R. Cryptococcus gattii infection in healthy hosts: A sentinel for subclinical immunodeficiency? Clin. Infect. Dis. 2012, 54, 153–154. [Google Scholar] [CrossRef]
- Abadi, J.; Pirofski, L.-A. Antibodies reactive with the cryptococcal capsular polysaccharide glucuronoxylomannan are present in sera from children with and without human immunodeficiency virus infection. J. Infect. Dis. 1999, 180, 915–919. [Google Scholar] [CrossRef]
- Deshaw, M.; Pirofski, L.A. Antibodies to the Cryptococcus neoformans capsular glucuronoxylomannan are ubiquitous in serum from HIV+ and HIV− individuals. Clin. Exp. Immunol. 1995, 99, 425–432. [Google Scholar] [CrossRef]
- Pirofski, L.-a. Polysaccharides, mimotopes and vaccines for fungal and encapsulated pathogens. Trends Microbiol. 2001, 9, 445–451. [Google Scholar] [CrossRef]
- Subramaniam, K.S.; Datta, K.; Quintero, E.; Manix, C.; Marks, M.S.; Pirofski, L.-A. The absence of serum IgM enhances the susceptibility of mice to pulmonary challenge with Cryptococcus neoformans. J. Immunol. 2010, 184, 5755–5767. [Google Scholar] [CrossRef]
- Rapaka, R.R.; Ricks, D.M.; Alcorn, J.F.; Chen, K.; Khader, S.A.; Zheng, M.; Plevy, S.; Bengtén, E.; Kolls, J.K. Conserved natural IgM antibodies mediate innate and adaptive immunity against the opportunistic fungus Pneumocystis murina. J. Exp. Med. 2010, 207, 2907–2919. [Google Scholar] [CrossRef]
- Szymczak, W.A.; Davis, M.J.; Lundy, S.K.; Dufaud, C.; Olszewski, M.; Pirofski, L.-a. X-linked immunodeficient mice exhibit enhanced susceptibility to Cryptococcus neoformans infection. MBio 2013, 4, e00265-13. [Google Scholar] [CrossRef]
- Moragues, M.D.; Omaetxebarria, M.J.; Elguezabal, N.; Sevilla, M.J.; Conti, S.; Polonelli, L.; Pontón, J. A monoclonal antibody directed against a Candida albicans cell wall mannoprotein exerts three anti-C. albicans activities. Infect. Immun. 2003, 71, 5273–5279. [Google Scholar] [CrossRef]
- Torosantucci, A.; Bromuro, C.; Chiani, P.; De Bernardis, F.; Berti, F.; Galli, C.; Norelli, F.; Bellucci, C.; Polonelli, L.; Costantino, P. A novel glyco-conjugate vaccine against fungal pathogens. J. Exp. Med. 2005, 202, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Torosantucci, A.; Chiani, P.; Bromuro, C.; De Bernardis, F.; Palma, A.S.; Liu, Y.; Mignogna, G.; Maras, B.; Colone, M.; Stringaro, A. Protection by anti-β-glucan antibodies is associated with restricted β-1,3 glucan binding specificity and inhibition of fungal growth and adherence. PLoS ONE 2009, 4, e5392. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.; Saylor, C.; Casadevall, A. Antibody action after phagocytosis promotes Cryptococcus neoformans and Cryptococcus gattii macrophage exocytosis with biofilm-like microcolony formation. Cell. Microbiol. 2008, 10, 1622–1633. [Google Scholar] [CrossRef] [PubMed]
- Maiti, P.K.; Kumar, A.; Kumar, R.; Mohapatra, L. Role of antibodies and effect of BCG vaccination in experimental candidiasis in mice. Mycopathologia 1985, 91, 79–85. [Google Scholar] [CrossRef]
- Cantorna, M.T.; Balish, E. Acquired immunity to systemic candidiasis in immunodeficient mice. J. Infect. Dis. 1991, 164, 936–943. [Google Scholar] [CrossRef]
- Narayanan, R.; Joyce, W.; Greenfield, R. Gastrointestinal candidiasis in a murine model of severe combined immunodeficiency syndrome. Infect. Immun. 1991, 59, 2116–2119. [Google Scholar] [CrossRef]
- Carrow, E.W.; Hector, R.; Domer, J.E. Immunodeficient CBA/N mice respond effectively to Candida albicans. Clin. Immunol. Immunopathol. 1984, 33, 371–380. [Google Scholar] [CrossRef]
- Sinha, B.; Prasad, S.; Monga, D. Studies of the role of B-cells in the resistance of mice to experimental candidiasis. Zent. Für Bakteriol. Mikrobiol. Und Hygiene. Ser. A Med. Microbiol. Infect. Dis. Virol. Parasitol. 1987, 266, 316–322. [Google Scholar] [CrossRef]
- Tristão, F.; Panagio, L.; Rocha, F.; Cavassani, K.; Moreira, A.; Rossi, M.; Silva, J. B cell-deficient mice display enhanced susceptibility to Paracoccidioides brasiliensis infection. Mycopathologia 2013, 176, 1–10. [Google Scholar] [CrossRef]
- K Mazu, T.; A Bricker, B.; Flores-Rozas, H.; Y Ablordeppey, S. The mechanistic targets of antifungal agents: An overview. Mini Rev. Med. Chem. 2016, 16, 555–578. [Google Scholar] [CrossRef]
- Nami, S.; Mohammadi, R.; Vakili, M.; Khezripour, K.; Mirzaei, H.; Morovati, H. Fungal vaccines, mechanism of actions and immunology: A comprehensive review. Biomed. Pharmacother. 2019, 109, 333–344. [Google Scholar] [CrossRef]
- Schmidt, C.S.; White, C.J.; Ibrahim, A.S.; Filler, S.G.; Fu, Y.; Yeaman, M.R.; Edwards, J.E., Jr.; Hennessey, J.P., Jr. NDV-3, a recombinant alum-adjuvanted vaccine for Candida and Staphylococcus aureus, is safe and immunogenic in healthy adults. Vaccine 2012, 30, 7594–7600. [Google Scholar] [CrossRef]
- Spellberg, B.J.; Ibrahim, A.S.; Avanesian, V.; Fu, Y.; Myers, C.; Phan, Q.T.; Filler, S.G.; Yeaman, M.R.; Edwards, J.E., Jr. Efficacy of the anti-Candida rAls3p-N or rAls1p-N vaccines against disseminated and mucosal candidiasis. J. Infect. Dis. 2006, 194, 256–260. [Google Scholar] [CrossRef]
- De Bernardis, F.; Amacker, M.; Arancia, S.; Sandini, S.; Gremion, C.; Zurbriggen, R.; Moser, C.; Cassone, A. A virosomal vaccine against candidal vaginitis: Immunogenicity, efficacy and safety profile in animal models. Vaccine 2012, 30, 4490–4498. [Google Scholar] [CrossRef]
- Ibrahim, A.S.; Spellberg, B.J.; Avanesian, V.; Fu, Y.; Edwards, J.E., Jr. The anti-Candida vaccine based on the recombinant N-terminal domain of Als1p is broadly active against disseminated candidiasis. Infect. Immun. 2006, 74, 3039–3041. [Google Scholar] [CrossRef]
- Baquir, B.; Lin, L.; Ibrahim, A.S.; Fu, Y.; Avanesian, V.; Tu, A.; Edwards, J., Jr.; Spellberg, B. Immunological reactivity of blood from healthy humans to the rAls3p-N vaccine protein. J. Infect. Dis. 2010, 201, 473–477. [Google Scholar] [CrossRef]
- Bozza, S.; Clavaud, C.; Giovannini, G.; Fontaine, T.; Beauvais, A.; Sarfati, J.; D’Angelo, C.; Perruccio, K.; Bonifazi, P.; Zagarella, S. Immune sensing of Aspergillus fumigatus proteins, glycolipids, and polysaccharides and the impact on Th immunity and vaccination. J. Immunol. 2009, 183, 2407–2414. [Google Scholar] [CrossRef]
- Sandini, S.; La Valle, R.; Deaglio, S.; Malavasi, F.; Cassone, A.; De Bernardis, F. A highly immunogenic recombinant and truncated protein of the secreted aspartic proteases family (rSap2t) of Candida albicans as a mucosal anticandidal vaccine. FEMS Immunol. Med. Microbiol. 2011, 62, 215–224. [Google Scholar] [CrossRef]
- Saville, S.P.; Lazzell, A.L.; Chaturvedi, A.K.; Monteagudo, C.; Lopez-Ribot, J.L. Efficacy of a genetically engineered Candida albicans tet-NRG1 strain as an experimental live attenuated vaccine against hematogenously disseminated candidiasis. Clin. Vaccine Immunol. 2009, 16, 430–432. [Google Scholar] [CrossRef]
- Martínez-López, R.; Nombela, C.; Diez-Orejas, R.; Monteoliva, L.; Gil, C. Immunoproteomic analysis of the protective response obtained from vaccination with Candida albicans ecm33 cell wall mutant in mice. Proteomics 2008, 8, 2651–2664. [Google Scholar] [CrossRef]
- Bistoni, F.; Vecchiarelli, A.; Cenci, E.; Puccetti, P.; Marconi, P.; Cassone, A. Evidence for macrophage-mediated protection against lethal Candida albicans infection. Infect. Immun. 1986, 51, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, C.; Correia, A.; Lima, T.; Vilanova, M.; Pais, C.; Gomes, A.C.; Oliveira, M.E.C.R.; Sampaio, P. Protective effect of antigen delivery using monoolein-based liposomes in experimental hematogenously disseminated candidiasis. Acta Biomater. 2016, 39, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Ulrich, M.A.; Cutler, J.E. Candida albicans mannan extract—Protein conjugates induce a protective immune response against experimental candidiasis. J. Infect. Dis. 1999, 179, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Pietrella, D.; Rachini, A.; Torosantucci, A.; Chiani, P.; Brown, A.J.; Bistoni, F.; Costantino, P.; Mosci, P.; d’Enfert, C.; Rappuoli, R. A β-glucan-conjugate vaccine and anti-β-glucan antibodies are effective against murine vaginal candidiasis as assessed by a novel in vivo imaging technique. Vaccine 2010, 28, 1717–1725. [Google Scholar] [CrossRef]
- Chiani, P.; Bromuro, C.; Cassone, A.; Torosantucci, A. Anti-β-glucan antibodies in healthy human subjects. Vaccine 2009, 27, 513–519. [Google Scholar] [CrossRef]
- Bromuro, C.; Romano, M.; Chiani, P.; Berti, F.; Tontini, M.; Proietti, D.; Mori, E.; Torosantucci, A.; Costantino, P.; Rappuoli, R. Beta-glucan-CRM197 conjugates as candidates antifungal vaccines. Vaccine 2010, 28, 2615–2623. [Google Scholar] [CrossRef]
- Paulovičová, E.; Machová, E.; Tulinská, J.; Bystrický, S. Cell and antibody mediated immunity induced by vaccination with novel Candida dubliniensis mannan immunogenic conjugate. Int. Immunopharmacol. 2007, 7, 1325–1333. [Google Scholar] [CrossRef]
- Cutler, J.E.; Corti, M.; Lambert, P.; Ferris, M.; Xin, H. Horizontal transmission of Candida albicans and evidence of a vaccine response in mice colonized with the fungus. PLoS ONE 2011, 6, e22030. [Google Scholar] [CrossRef]
- Xin, H.; Cutler, J.E. Vaccine and monoclonal antibody that enhance mouse resistance to candidiasis. Clin. Vaccine Immunol. 2011, 18, 1656–1667. [Google Scholar] [CrossRef]
- Cárdenas-Freytag, L.; Cheng, E.; Mayeux, P.; Domer, J.E.; Clements, J.D. Effectiveness of a vaccine composed of heat-killed Candida albicans and a novel mucosal adjuvant, LT (R192G), against systemic candidiasis. Infect. Immun. 1999, 67, 826–833. [Google Scholar] [CrossRef]
- Luo, G.; Ibrahim, A.S.; French, S.W.; Edwards, J.E., Jr.; Fu, Y. Active and passive immunization with rHyr1p-N protects mice against hematogenously disseminated candidiasis. PLoS ONE 2011, 6, e25909. [Google Scholar] [CrossRef]
- Xin, H.; Dziadek, S.; Bundle, D.R.; Cutler, J.E. Synthetic glycopeptide vaccines combining β-mannan and peptide epitopes induce protection against candidiasis. Proc. Natl. Acad. Sci. USA 2008, 105, 13526–13531. [Google Scholar] [CrossRef]
- Cenci, E.; Mencacci, A.; Bacci, A.; Bistoni, F.; Kurup, V.P.; Romani, L. T cell vaccination in mice with invasive pulmonary aspergillosis. J. Immunol. 2000, 165, 381–388. [Google Scholar] [CrossRef]
- Diaz-Arevalo, D.; Bagramyan, K.; Hong, T.B.; Ito, J.I.; Kalkum, M. CD4+ T cells mediate the protective effect of the recombinant Asp f3-based anti-aspergillosis vaccine. Infect. Immun. 2011, 79, 2257–2266. [Google Scholar] [CrossRef]
- Ito, J.I.; Lyons, J.M. Vaccination of corticosteroid immunosuppressed mice against invasive pulmonary aspergillosis. J. Infect. Dis. 2002, 186, 869–871. [Google Scholar] [CrossRef]
- Ito, J.I.; Lyons, J.M.; Hong, T.B.; Tamae, D.; Liu, Y.-K.; Wilczynski, S.P.; Kalkum, M. Vaccinations with recombinant variants of Aspergillus fumigatus allergen Asp f 3 protect mice against invasive aspergillosis. Infect. Immun. 2006, 74, 5075–5084. [Google Scholar] [CrossRef]
- Clemons, K.V.; Martinez, M.; Chen, V.; Liu, M.; Yoon, H.J.; Stevens, D.A. Protection against experimental aspergillosis by heat-killed yeast is not antibody dependent. Sabouraudia 2014, 52, 422–426. [Google Scholar] [CrossRef]
- Stuehler, C.; Khanna, N.; Bozza, S.; Zelante, T.; Moretti, S.; Kruhm, M.; Lurati, S.; Conrad, B.; Worschech, E.; Stevanović, S. Cross-protective TH1 immunity against Aspergillus fumigatus and Candida albicans. Blood J. Am. Soc. Hematol. 2011, 117, 5881–5891. [Google Scholar] [CrossRef]
- Bozza, S.; Gaziano, R.; Lipford, G.B.; Montagnoli, C.; Bacci, A.; Di Francesco, P.; Kurup, V.P.; Wagner, H.; Romani, L. Vaccination of mice against invasive aspergillosis with recombinant Aspergillus proteins and CpG oligodeoxynucleotides as adjuvants. Microbes Infect. 2002, 4, 1281–1290. [Google Scholar] [CrossRef]
- Wüthrich, M.; Filutowicz, H.I.; Warner, T.; Deepe, G.S., Jr.; Klein, B.S. Vaccine immunity to pathogenic fungi overcomes the requirement for CD4 help in exogenous antigen presentation to CD8+ T cells: Implications for vaccine development in immune-deficient hosts. J. Exp. Med. 2003, 197, 1405–1416. [Google Scholar] [CrossRef]
- Pinto, A.R.; Puccia, R.; Diniz, S.N.; Franco, M.F.; Travassos, L.R. DNA-based vaccination against murine paracoccidioidomycosis using the gp43 gene from Paracoccidioides brasiliensis. Vaccine 2000, 18, 3050–3058. [Google Scholar] [CrossRef] [PubMed]
- Assis-Marques, M.A.; Oliveira, A.F.; Ruas, L.P.; Reis, T.F.d.; Roque-Barreira, M.C.; Coelho, P.S.R. Saccharomyces cerevisiae expressing Gp43 protects mice against Paracoccidioides brasiliensis infection. PLoS ONE 2015, 10, e0120201. [Google Scholar] [CrossRef] [PubMed]
- Braga, C.J.; Rittner, G.M.; Munoz Henao, J.E.; Teixeira, A.F.; Massis, L.M.; Sbrogio-Almeida, M.E.; Taborda, C.P.; Travassos, L.R.; Ferreira, L.C. Paracoccidioides brasiliensis vaccine formulations based on the gp43-derived P10 sequence and the Salmonella enterica FliC flagellin. Infect. Immun. 2009, 77, 1700–1707. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, V.C.; Martins, E.M.; Boeloni, J.N.; Coitinho, J.B.; Serakides, R.; Goes, A.M. Additive effect of rPb27 immunization and chemotherapy in experimental paracoccidioidomycosis. PLoS ONE 2011, 6, e17885. [Google Scholar] [CrossRef]
- De Bastos Ascenço Soares, R.; Gomez, F.J.; de Almeida Soares, C.M.; Deepe, G.S., Jr. Vaccination with heat shock protein 60 induces a protective immune response against experimental Paracoccidioides brasiliensis pulmonary infection. Infect. Immun. 2008, 76, 4214–4221. [Google Scholar] [CrossRef]
- Ribeiro, A.M.; Bocca, A.L.; Amaral, A.C.; Souza, A.C.C.; Faccioli, L.H.; Coelho-Castelo, A.A.; Figueiredo, F.; Silva, C.L.; Felipe, M.S.S. HSP65 DNA as therapeutic strategy to treat experimental paracoccidioidomycosis. Vaccine 2010, 28, 1528–1534. [Google Scholar] [CrossRef]
- Devi, S.J. Preclinical efficacy of a glucuronoxylomannan-tetanus toxoid conjugate vaccine of Cryptococcus neoformans in a murine model. Vaccine 1996, 14, 841–844. [Google Scholar] [CrossRef]
- Chow, S.-K.; Casadevall, A. Evaluation of Cryptococcus neoformans galactoxylomannan–protein conjugate as vaccine candidate against murine cryptococcosis. Vaccine 2011, 29, 1891–1898. [Google Scholar] [CrossRef]
- Wozniak, K.L.; Young, M.L.; Wormley, F.L., Jr. Protective immunity against experimental pulmonary cryptococcosis in T cell-depleted mice. Clin. Vaccine Immunol. 2011, 18, 717–723. [Google Scholar] [CrossRef]
- Rella, A.; Mor, V.; Farnoud, A.M.; Singh, A.; Shamseddine, A.A.; Ivanova, E.; Carpino, N.; Montagna, M.T.; Luberto, C.; Del Poeta, M. Role of Sterylglucosidase 1 (Sgl1) on the pathogenicity of Cryptococcus neoformans: Potential applications for vaccine development. Front. Microbiol. 2015, 6, 836. [Google Scholar] [CrossRef]
- Specht, C.A.; Nong, S.; Dan, J.M.; Lee, C.K.; Levitz, S.M. Contribution of glycosylation to T cell responses stimulated by recombinant Cryptococcus neoformans mannoprotein. J. Infect. Dis. 2007, 196, 796–800. [Google Scholar] [CrossRef]
- Oscarson, S.; Alpe, M.; Svahnberg, P.; Nakouzi, A.; Casadevall, A. Synthesis and immunological studies of glycoconjugates of Cryptococcus neoformans capsular glucuronoxylomannan oligosaccharide structures. Vaccine 2005, 23, 3961–3972. [Google Scholar] [CrossRef]
- Rachini, A.; Pietrella, D.; Lupo, P.; Torosantucci, A.; Chiani, P.; Bromuro, C.; Proietti, C.; Bistoni, F.; Cassone, A.; Vecchiarelli, A. An anti-β-glucan monoclonal antibody inhibits growth and capsule formation of Cryptococcus neoformans in vitro and exerts therapeutic, anticryptococcal activity in vivo. Infect. Immun. 2007, 75, 5085–5094. [Google Scholar] [CrossRef]
- Zheng, M.; Ramsay, A.J.; Robichaux, M.B.; Norris, K.A.; Kliment, C.; Crowe, C.; Rapaka, R.R.; Steele, C.; McAllister, F.; Shellito, J.E. CD4+ T cell–independent DNA vaccination against opportunistic infections. J. Clin. Investig. 2005, 115, 3536–3544. [Google Scholar] [CrossRef]
- Feng, Y.; Guo, S.; Jiang, T.; Han, X.; Liu, P.; Wu, T.; Luo, Y. Active immunization against Pneumocystis carinii with p55-v3 DNA vaccine in rats. Can. J. Microbiol. 2011, 57, 375–381. [Google Scholar] [CrossRef]
- Theus, S.A.; Smulian, A.G.; Steele, P.; Linke, M.J.; Walzer, P.D. Immunization with the major surface glycoprotein of Pneumocystis carinii elicits a protective response. Vaccine 1998, 16, 1149–1157. [Google Scholar] [CrossRef]
- Garcia, J.P.; Howard, D.H. Characterization of antigens from the yeast phase of Histoplasma capsulatum. Infect. Immun. 1971, 4, 116–125. [Google Scholar] [CrossRef]
- Tewari, R.; Sharma, D.; Solotorovsky, M.; Lafemina, R.; Balint, J. Adoptive transfer of immunity from mice immunized with ribosomes or live yeast cells of Histoplasma capsulatum. Infect. Immun. 1977, 15, 789–795. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Steenbergen, J.N.; Shi, L.; Deepe, G.S.; Casadevall, A. Antibodies to a cell surface histone-like protein protect against Histoplasma capsulatum. J. Clin. Investig. 2003, 112, 1164–1175. [Google Scholar] [CrossRef]
- Deepe, J.; George, S.; Gibbons, R.S. Cellular and molecular regulation of vaccination with heat shock protein 60 from Histoplasma capsulatum. Infect. Immun. 2002, 70, 3759–3767. [Google Scholar] [CrossRef]
- Gomez, F.; Gomez, A.; Deepe, G., Jr. Protective efficacy of a 62-kilodalton antigen, HIS-62, from the cell wall and cell membrane of Histoplasma capsulatum yeast cells. Infect. Immun. 1991, 59, 4459–4464. [Google Scholar] [CrossRef] [PubMed]
- Gomez, F.J.; Gomez, A.; Deepe, G.S., Jr. An 80-kilodalton antigen from Histoplasma capsulatum that has homology to heat shock protein 70 induces cell-mediated immune responses and protection in mice. Infect. Immun. 1992, 60, 2565–2571. [Google Scholar] [CrossRef] [PubMed]
- Scheckelhoff, M.R.; Deepe, G.S. Pulmonary Vβ4+ T Cells from Histoplasma capsulatum–Infected Mice Respond to a Homologue of Sec31 That Confers a Protective Response. J. Infect. Dis. 2006, 193, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Deepe, G.S., Jr.; Gibbons, R. Protective efficacy of H antigen from Histoplasma capsulatum in a murine model of pulmonary histoplasmosis. Infect. Immun. 2001, 69, 3128–3134. [Google Scholar] [CrossRef]
- Cheah, H.-L.; Lim, V.; Sandai, D. Inhibitors of the glyoxylate cycle enzyme ICL1 in Candida albicans for potential use as antifungal agents. PLoS ONE 2014, 9, e95951. [Google Scholar] [CrossRef]
- Gallis, H.A.; Drew, R.H.; Pickard, W.W. Amphotericin B: 30 years of clinical experience. Rev. Infect. Dis. 1990, 12, 308–329. [Google Scholar] [CrossRef]
- Bolard, J. How do the polyene macrolide antibiotics affect the cellular membrane properties? Biochim. Et Biophys. Acta (BBA)-Rev. Biomembr. 1986, 864, 257–304. [Google Scholar] [CrossRef]
- Villa, N.Y.; Moussatche, P.; Chamberlin, S.G.; Kumar, A.; Lyons, T.J. Phylogenetic and preliminary phenotypic analysis of yeast PAQR receptors: Potential antifungal targets. J. Mol. Evol. 2011, 73, 134–152. [Google Scholar] [CrossRef]
- Warnock, D.W. Amphotericin B: An introduction. J. Antimicrob. Chemother. 1991, 28, 27–38. [Google Scholar] [CrossRef]
- Delmas, G.; Park, S.; Chen, Z.; Tan, F.; Kashiwazaki, R.; Zarif, L.; Perlin, D. Efficacy of orally delivered cochleates containing amphotericin B in a murine model of aspergillosis. Antimicrob. Agents Chemother. 2002, 46, 2704–2707. [Google Scholar] [CrossRef]
- Di Mambro, T.; Guerriero, I.; Aurisicchio, L.; Magnani, M.; Marra, E. The yin and yang of current antifungal therapeutic strategies: How can we harness our natural defenses? Front. Pharmacol. 2019, 10, 80. [Google Scholar] [CrossRef]
- Hata, K.; Kimura, J.; Miki, H.; Toyosawa, T.; Nakamura, T.; Katsu, K. In vitro and in vivo antifungal activities of ER-30346, a novel oral triazole with a broad antifungal spectrum. Antimicrob. Agents Chemother. 1996, 40, 2237–2242. [Google Scholar] [CrossRef]
- Patton, J.L.; Lester, R.L. The phosphoinositol sphingolipids of Saccharomyces cerevisiae are highly localized in the plasma membrane. J. Bacteriol. 1991, 173, 3101–3108. [Google Scholar] [CrossRef]
- Nimrichter, L.; Rodrigues, M.L. Fungal glucosylceramides: From structural components to biologically active targets of new antimicrobials. Front. Microbiol. 2011, 2, 212. [Google Scholar] [CrossRef]
- Monk, B.C.; Mason, A.B.; Abramochkin, G.; Haber, J.E.; Seto-Young, D.; Perlin, D.S. The yeast plasma membrane proton pumping ATPase is a viable antifungal target. I. Effects of the cysteine-modifying reagent omeprazole. Biochim. Biophys. Acta (BBA)-Biomembr. 1995, 1239, 81–90. [Google Scholar] [CrossRef]
- Chen, S.C.-A.; Slavin, M.A.; Sorrell, T.C. Echinocandin antifungal drugs in fungal infections. Drugs 2011, 71, 11–41. [Google Scholar] [CrossRef]
- Perlin, D.S. Current perspectives on echinocandin class drugs. Future Microbiol. 2011, 6, 441–457. [Google Scholar] [CrossRef]
- Mukhopadhyay, T.; Roy, K.; Bhat, R.; Sawant, S.; Blumbach, J.; Ganguli, B.; Fehlhaber, H.; Kogler, H. Deoxymulundocandin-a new echinocandin type antifungal antibiotic. J. Antibiot. 1992, 45, 618–623. [Google Scholar] [CrossRef]
- Walker, S.S.; Xu, Y.; Triantafyllou, I.; Waldman, M.F.; Mendrick, C.; Brown, N.; Mann, P.; Chau, A.; Patel, R.; Bauman, N. Discovery of a novel class of orally active antifungal β-1, 3-d-glucan synthase inhibitors. Antimicrob. Agents Chemother. 2011, 55, 5099–5106. [Google Scholar] [CrossRef]
- Shubitz, L.F.; Trinh, H.T.; Perrill, R.H.; Thompson, C.M.; Hanan, N.J.; Galgiani, J.N.; Nix, D.E. Modeling nikkomycin Z dosing and pharmacology in murine pulmonary coccidioidomycosis preparatory to phase 2 clinical trials. J. Infect. Dis. 2014, 209, 1949–1954. [Google Scholar] [CrossRef]
- Vijayakumar, E.; Roy, K.; Chatterjee, S.; Deshmukh, S.; Ganguli, B.; Fehlhaber, H.-W.; Kogler, H. Arthrichitin. A new cell wall active metabolite from Arthrinium phaeospermum. J. Org. Chem. 1996, 61, 6591–6593. [Google Scholar] [CrossRef] [PubMed]
- Fostel, J.M.; Montgomery, D.A.; Shen, L.L. Characterization of DNA topoisomerase I from Candida albicans as a target for drug discovery. Antimicrob. Agents Chemother. 1992, 36, 2131–2138. [Google Scholar] [CrossRef] [PubMed]
- Fostel, J.; Montgomery, D. Identification of the aminocatechol A-3253 as an in vitro poison of DNA topoisomerase I from Candida albicans. Antimicrob. Agents Chemother. 1995, 39, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Levitz, S.M. Aspergillus vaccines: Hardly worth studying or worthy of hard study? Sabouraudia 2016, 55, 103–108. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Fungal Pathogen | Disease | Organs Affected |
|---|---|---|
| Aspergillus | Aspergillosis | Usually occurs in people with lung diseases or weakened immune systems. |
| Candida | Candidiasis | Commonly affects the mouth and throat, vagina, or the bloodstream. |
| Cryptococcus | Cryptococcosis | Infect the lungs and/or central nervous system (cryptococcal meningitis). |
| Pneumocystis | Pneumocystis pneumonia | Infect mainly lungs when the host immune system becomes compromised. |
| Mucormycetes | Aggressive infections | Organs such the brain, spleen, heart, and skin by spreading locally and through the circulation. |
| Histoplasma | Histoplasmosis | Infect the lungs and/or central nervous system (cryptococcal meningitis). |
| Fungal Infection | Number Affected | Case Fatality Rate | Estimated Deaths |
|---|---|---|---|
| Cryptococcal meningitis | 223,000 in AIDS | 15–20% USA >50% developing world | 180,000 in AIDS |
| Pneumocystis pneumonia | >400,000 in AIDS >100,000 in non-AIDS | ~15% in AIDS with best treatment ~50% in non-AIDS | >200,000 in AIDS >50,000 non-AIDS |
| Disseminated histoplasmosis | ~100,000 | 15–30%, if diagnosed and treated | >80,000 |
| Invasive aspergillosis | >1,000,000 | ~30% mortality in leukemia in HIC | >500,000 |
| Invasive candidiasis | >750,000 | ~40% mortality treated | >350,000 |
| Chronic pulmonary aspergillosis | >3,000,000 | ~15–40% mortality in HIC ~15% mortality in the developed world | >450,000 in non-hospitalized populations |
| Severe asthma with fungal sensitization (SAFS) | >6,500,000 | <1% but no good figures | 350,000–489,000 asthma deaths ~50% related to SAFS |
| Fungal keratitis | 1.0–1.4 million | Blinding > 60% | >600,000 blind eyes |
| Total | ~13,500,000 | >1,600,000 |
| Host PRR | Pathogens | Fungal PAMPs |
|---|---|---|
| CLR Dectin-1 | Candidaspp., A. fumigates, Pneumocystisspp., Coccidioidesspp., Fonsecaeaspp., T. rubrum | β (1,3)- glucan |
| Dectin-2 | Candidaspp., Malasseziaspp., A. fumigates, P. brasiliensis, Pneumocystisspp., C. posadasii, F. pedrosoi, H. capsulatum, T. rubrum, C. neoformans | α-mannan, high mannose structures |
| Dectin-3 | C. albicans | α-mannan |
| Mincle | C. albicans, A. fumigatus, Pneumocystisspp., Fonsecaeaspp., Malasseziaspp. | α-mannose |
| Mcl | C. albicans, B. dermatitidis | α-mannose |
| DC-SIGN | C. albicans, A. fumigates, T. rubrum, P. brasiliensis | Mannose, surface carbohydrate in extracellular vesicles |
| MR | C. neoformans, P. carinii, C. immitis, P. brasiliensis, H. capsulatum, T. rubrum | Mannose |
| Mannose receptor | C. albicans, C. neoformans, A. fumigates, P. brasiliensis | Mannan (N-linked), mannoproteins, mannan, gp43 |
| Galectin-3 | C. albicans | β-1,2-mannosides |
| Scarf1/CDC36 | C. albicans, C. neoformans | β (1,3)- glucan |
| TLR TLR2 | C. albicans, Alternaria, A. fumigatus | Phospholipomannans, α-glucans zymosan |
| TRL3 | A. fumigatus | dsRNA |
| TLR4 | C. albicans, A. fumigatus | α-, β-glucan, and galactomannan N |
| TLR6 | Candidaspp. | Zymosan, unmethylated DNA with CpG motif |
| TLR7 | Candidaspp. | ssRNA |
| TLR9 | C. neoformans, C. albicans, A. fumigatus | Zymosan, CpG-oligodeoxynucleotides |
| NLR NLRP3 | C. albicans, A. fumigatus, C. neoformans, Malasseziaspp., P. brasiliensis, S. schenckii, H. capsulatum | Unknown |
| NLRP4 | C. albicans | Unknown |
| NLRP10 | C. albicans | Unknown |
| NOD1 | A. fumigatus | Unknown |
| NOD2 | C. parapsilosis, A. fumigatus | Chitin |
| RLR MDA5 | C. albicans | Unknown |
| Pathogen | Antigen | Vaccine Type | Underlying Immune Mechanism | Reference |
|---|---|---|---|---|
| Candidiasis | Als3p Als1p | Recombinant protein (NDV-3) | IgG, IL17 A, IFN-γ, and IgA1, | [93,94,95,96,97,98] |
| SAP2 | Recombinant protein | Protective antibodies | [95,99] | |
| aspartyl proteinase protein, Sap2p (Secreted fraction) | Recombinant | Antibodies | [99] | |
| Tet-NRG1 | Recombinant and live attenuated | Elicit immunity mediated by T cells | [100,101] | |
| PCA-2 strain of Candida albicans | Live-attenuated type | Elevated activity of macrophages and polymorphonuclear leukocyte | [102] | |
| Surface proteins of the cell wall | Protein subunit vaccine type | Immunoglobulins- and Th17 cytokine-mediated response | [103] | |
| Mannose derivatives of Candida albicans | Protein conjugate vaccine with mannan derivatives | Immunoglobulin responses | [104] | |
| Laminarin (Lam) β-polysaccharides | Lam diphtheria toxoid CRM197 conjugate | Passive immunoglobulin responses | [82,83,104,105,106,107] | |
| C. dubliniensis mannan and human serum albumin | Conjugate vaccine type | Immunoglobulin (IgG, IgA)-mediated and Th1-mediated responses | [108] | |
| Fructose bisphosphate aldolase (Fba) (cytosolic and cell wall peptides | Subunit vaccine type | Immunoglobulin response | [109,110] | |
| Heat-killed C. albicans (HK-CA) | Recombinant vaccine type | Antibody (IgG, IgA)- Th1 | [111] | |
| Cell surface protein named Hyr 1 from C. albicans. | Recombinant vaccine type (using N-terminal part) | Immunoglobulin response | [112] | |
| Combination of peptide epitopes and β-mannan conjugates | Subunit/conjugate vaccine type | Immunoglobulin-mediated and Th1- mediated responses | [113] | |
| Aspergillosis | Crude culture of Aspergillus fumigatus filtrate | Sonicated and filtered fractions subunit-based vaccine | Th1 cells-mediated IFN-γ and IL-2 cytokine-induced response | [114] |
| Asp f3 | Incomplete Freund’s adjuvant | Immunoglobulin- and T cell-mediated response | [115] | |
| Viable conidia of Aspergillus fumigatus | Sonicated and filtered fractions subunit-based vaccine | Not yet Described | [116] | |
| Aspergillus fumigatus hyphal sonicate (HS) | Recombinant | Immunoglobulin-mediated and T cell-mediated responses | [117] | |
| Heat-killed Saccharomyces sp. | Live-attenuated vaccine types | Mostly cell-mediated immune response mediated by Th1, Th2, and Th17 cytokines | [117,118] | |
| Cell wall epitope (p41 of gluconate, Crfl1) from Aspergillus fumigatus | Subunit | MHC 2-mediated T cell-based protection and protection against lethal infection with C. albicans in cross-section | [119] | |
| Asp 16 f | Recombinant vaccine type | Th1 | [120] | |
| Asp 3 f | Recombinant vaccine type | Th1 | [117] | |
| Various proteins Gel1p, Mep1p, Crf1p, Sod1p, Dpp5p, RNUp, Pep1p, Cat1p, Polysaccharides: β1–3 glucan, β1–3 glucan, GM, glycolipids: GSL, LGM | Recombinant/Subunit type vaccine | Th1 | [98] | |
| Blastomycosis | Adhesin BAD1 gene | Whole organism/live attenuated vaccine | Cytotoxic T cell-mediated immunity | [121] |
| Paracoccidioidomycosis (PCM) | gp 43 (P10) | DNA vaccine (pcDNA3-P10) | T regulatory cells-based immunological memory | [122] |
| gp 43 (P10) | Recombinant protein vaccine | Th1 cell immunity with concurrent role of IL-12 and IFN-γ cytokines | [123] | |
| P10- FliC fusion protein | Recombinant | Th1 | [124] | |
| rPb27 | Recombinant | Immunoglobulin-based humoral immunity | [125] | |
| Heat shock protein (Hsp 60) | Recombinant | Helper cell-mediated immunity | [126] | |
| Mycobacterium leprae derived HSP65 | Recombinant DNA | Th1 | [127] | |
| Cryptococcosis | GXM | Conjugate/soluble antigenic fractions | Anti-GMX antibodies (active immunization) | [128] |
| GalXM | Subunit/conjugate Vaccine | Immunoglobulin (IgG, IgA)-mediated immunity | [129] | |
| Serotype A and Matα of C. neoformans strain H99γ | Live attenuated | Th- and cytotoxic T cell-mediated immunity | [130] | |
| Mutant C. neoformans strain lacking the enzyme steryl glucosidase 1 named (Δsgl1) | Live attenuated recombinant | Th- and cytotoxic T cell-mediated immunity | [131] | |
| CneF (culture filtrate Ags), mannoprotein | Subunit/recombinant | Immunoglobulin (IgG, IgA)-mediated and Th1-mediated responses | [132] | |
| GXM | GXM–protein conjugate | High-titer IgG responses | [133] | |
| Laminaran | Subunit (algal βglucan based) | Passive immunity | [134] | |
| Pneumocystis | Kexin genes | Kexin-CD40 L DNA vaccine | Elevated IgG titers | [135] |
| P55 protein | Recombinant protein | Th1-Th2 responses | [136] | |
| gp120 | Recombinant protein | Immunoglobulin (IgG, IgA)-mediated and Th1-mediated responses | [137] | |
| Histoplasmosis | Ethylenediamine extract from cell wall in aqueous formulation | Inactivated filtrated antigen combined with soluble antigenic fractions | Nd | [138] |
| Ribosomes or live yeast cells of H. capsulatum | Live attenuated vaccine type | Lymphoid cell-mediated immunity | [139] | |
| Histone H2B–like protein | Live attenuated and recombinant type | Immunoglobulin (IgG, IgA)-mediated and Th1-mediated responses | [140] | |
| HSP-60 | Recombinant type vaccine | Th1 | [141] | |
| HIS-62 | Recombinant protein vaccine | Cellular immune response | [142] | |
| 80 kDa antigen | Recombinant vaccine | Immunoglobulin (IgG, IgA)-mediated and Th1-mediated responses | [143] | |
| Sec31 homologue | Recombinant | T cell-mediated | [144] | |
| H antigen | Recombinant antigen | Th1.Th2/CD8+ | [145] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabaan, A.A.; Alfaraj, A.H.; Alshengeti, A.; Alawfi, A.; Alwarthan, S.; Alhajri, M.; Al-Najjar, A.H.; Al Fares, M.A.; Najim, M.A.; Almuthree, S.A.; et al. Antibodies to Combat Fungal Infections: Development Strategies and Progress. Microorganisms 2023, 11, 671. https://doi.org/10.3390/microorganisms11030671
Rabaan AA, Alfaraj AH, Alshengeti A, Alawfi A, Alwarthan S, Alhajri M, Al-Najjar AH, Al Fares MA, Najim MA, Almuthree SA, et al. Antibodies to Combat Fungal Infections: Development Strategies and Progress. Microorganisms. 2023; 11(3):671. https://doi.org/10.3390/microorganisms11030671
Chicago/Turabian StyleRabaan, Ali A., Amal H. Alfaraj, Amer Alshengeti, Abdulsalam Alawfi, Sara Alwarthan, Mashael Alhajri, Amal H. Al-Najjar, Mona A. Al Fares, Mustafa A. Najim, Souad A. Almuthree, and et al. 2023. "Antibodies to Combat Fungal Infections: Development Strategies and Progress" Microorganisms 11, no. 3: 671. https://doi.org/10.3390/microorganisms11030671