
Contrasting Patterns of Asaia Association with Pyrethroid Resistance Escalation between the Malaria Vectors Anopheles funestus and Anopheles gambiae
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Strains and Rearing
2.2. Evaluating the Susceptibility Profile of An. funestus and An. gambiae Strains to Pyrethroids
2.3. Assessing the Contribution of Known Molecular Markers to the Escalation of Pyrethroid Resistance in An. funestus and An. gambiae
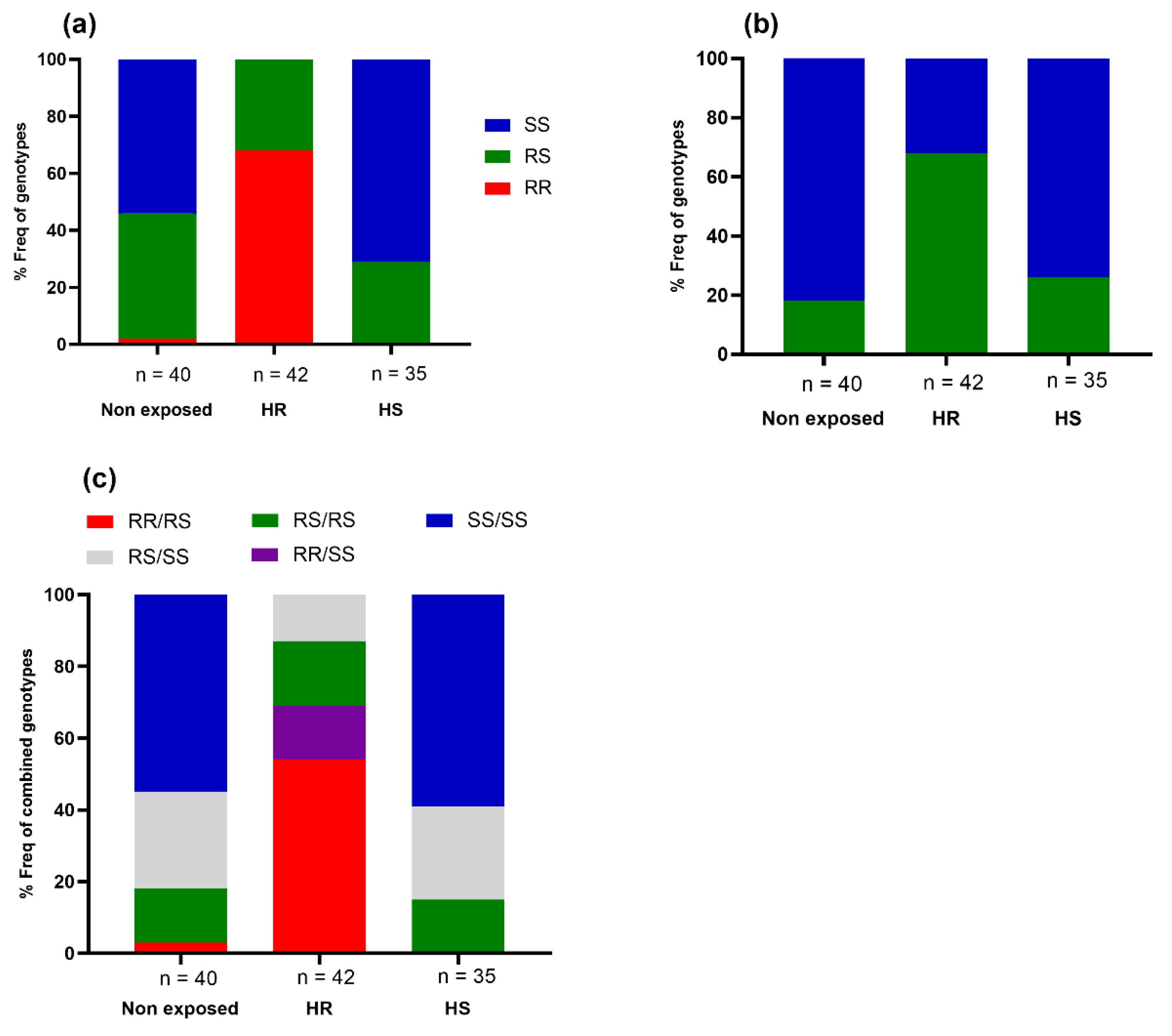
2.3.1. Contribution of CYP6P9a, CYP6P9b, and 6.5 kb-SV Mutations to the Escalation of Pyrethroid Resistance in An. funestus
2.3.2. Contribution of the Voltage-Gated Sodium Channel (VGSC) Mutations, L1014F and N1575Y, to the Escalation of Pyrethroid Resistance in An. gambiae
2.4. Assessing the Contribution of Asaia spp. in the Escalation of Pyrethroid Resistance in An. funestus and An. gambiae Mosquitoes
2.4.1. Detection of Asaia spp. Frequency in Anopheles Strains
2.4.2. Quantification of Asaia spp. in Anopheles Strain
3. Results
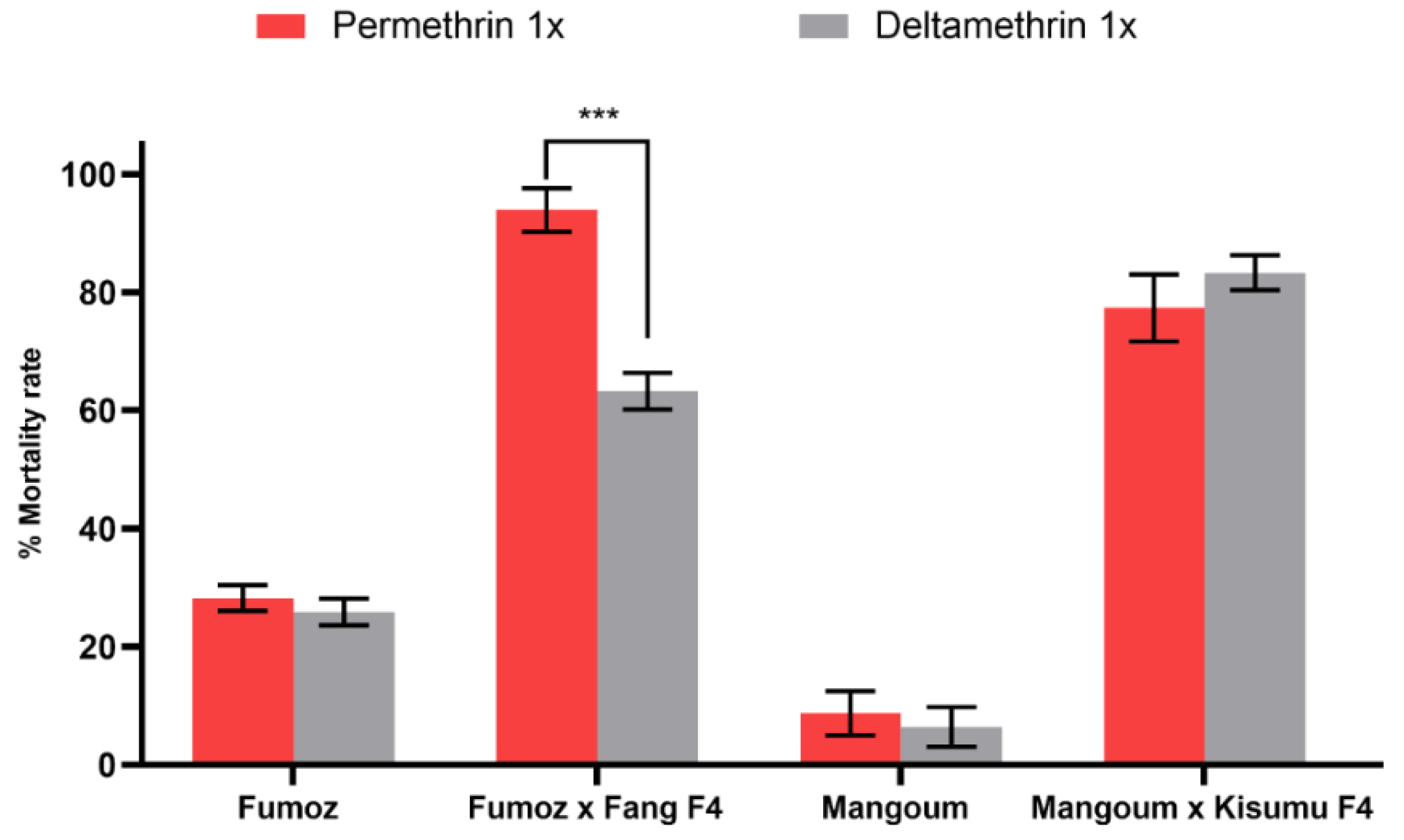
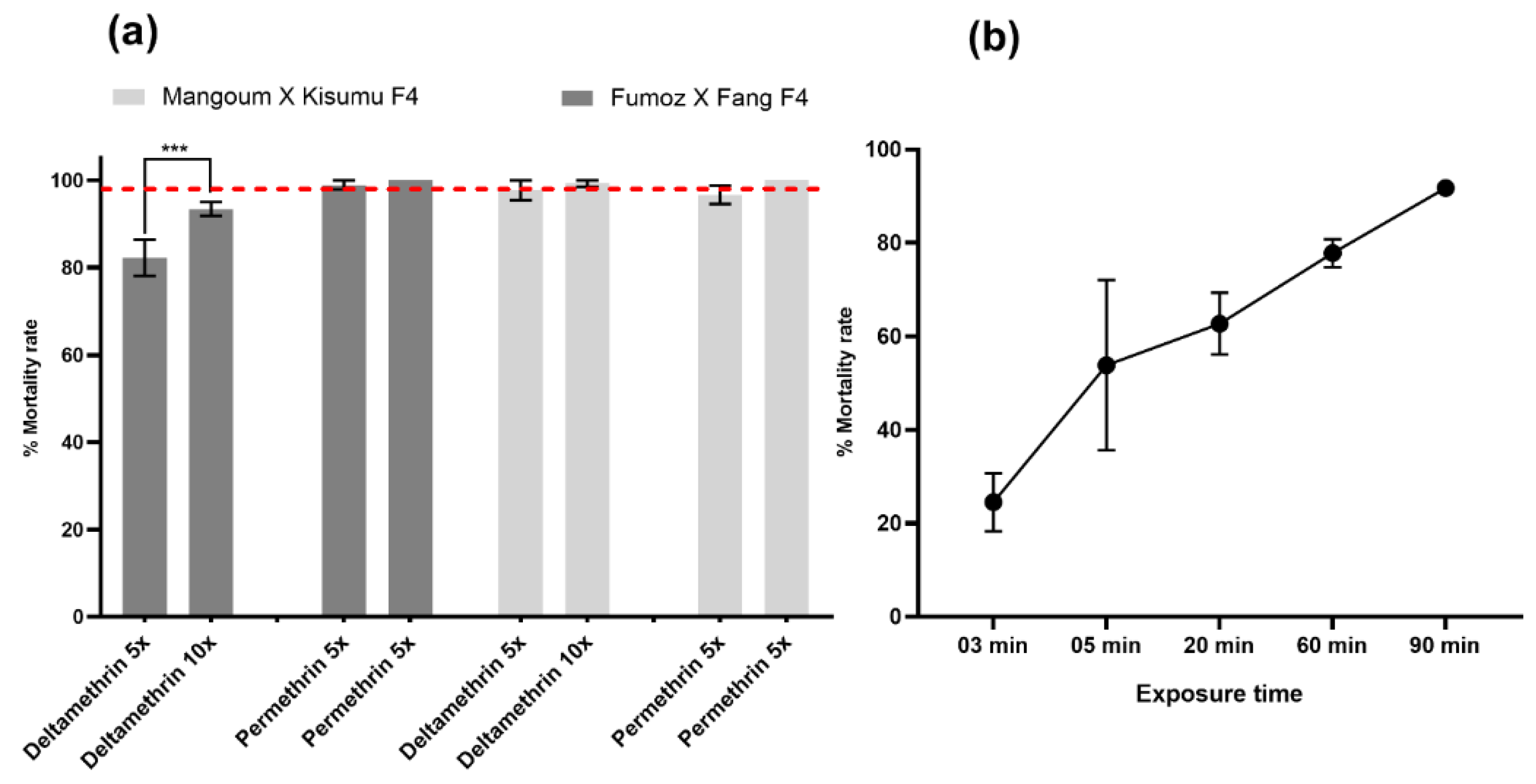
3.1. Resistance Profile of Anopheles Funestus and Anopheles gambiae Strains
3.2. Contribution of Cytochrome P450 Markers in the Escalation of Deltamethrin Resistance in An. funestus
3.3. Impact of VGSC Mutations on the Escalation of Permethrin Resistance in An. gambiae
3.4. Contribution of Asaia spp. to the Escalation of Pyrethroid Insecticide Resistance in Malaria Vectors
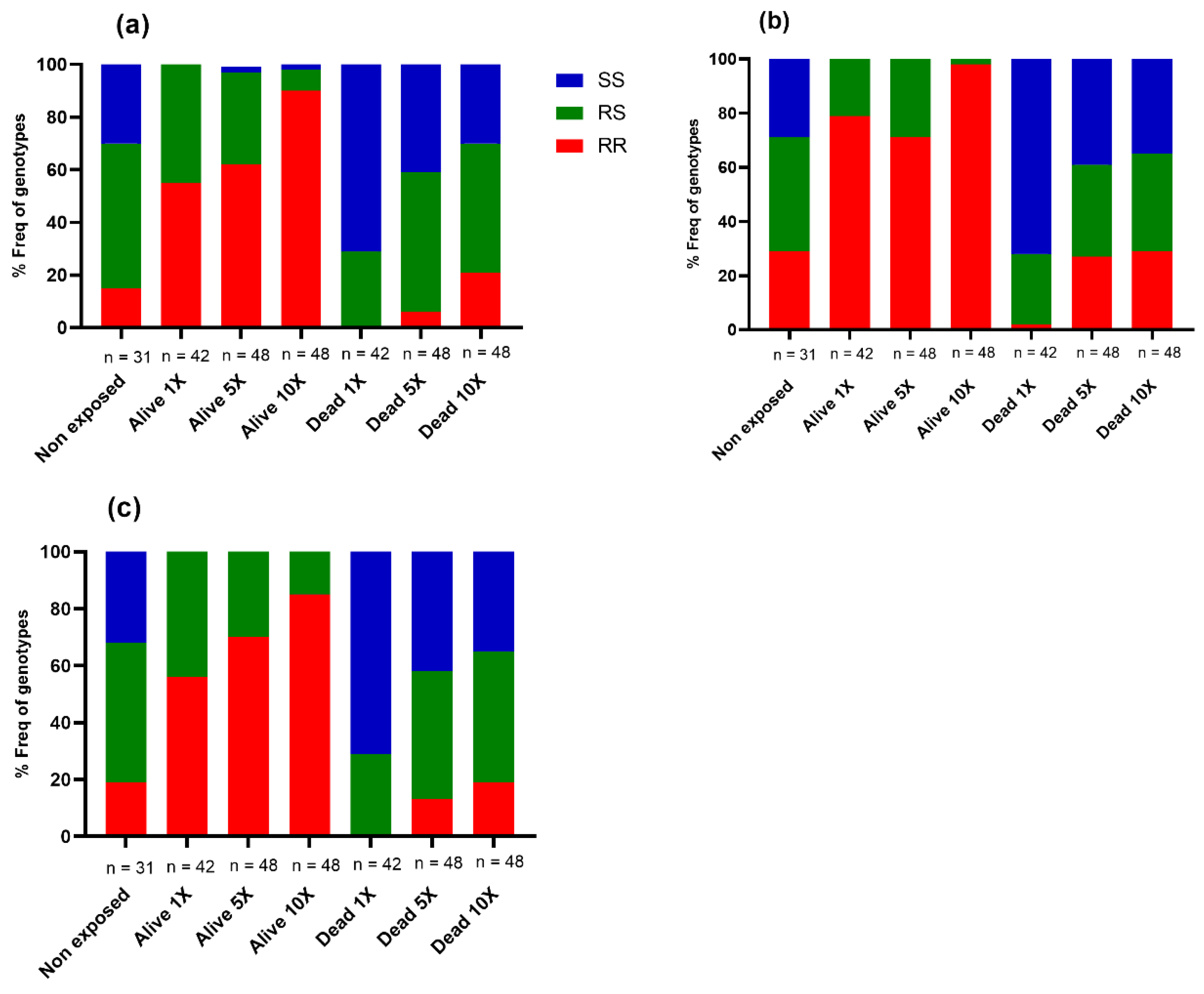
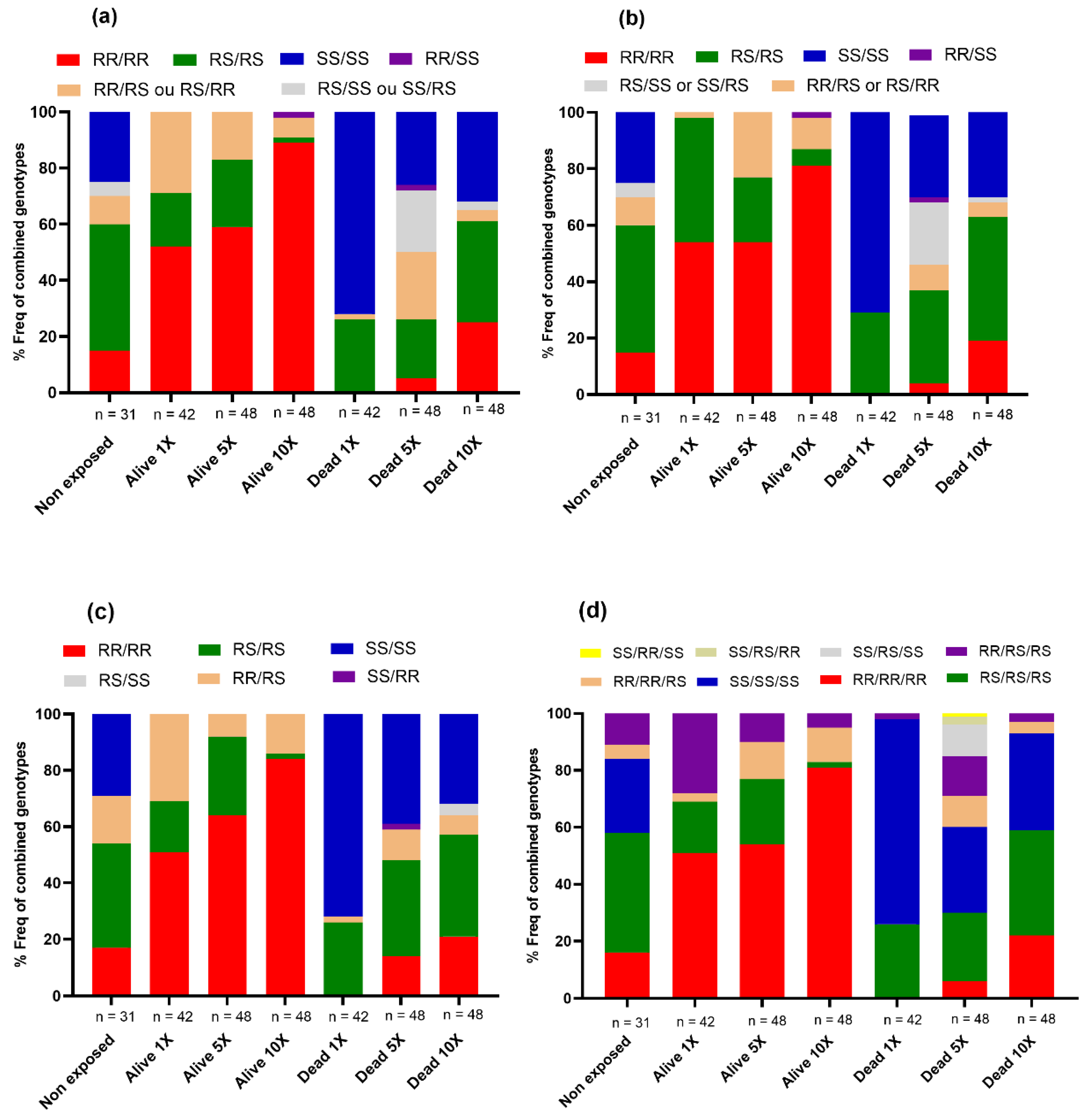
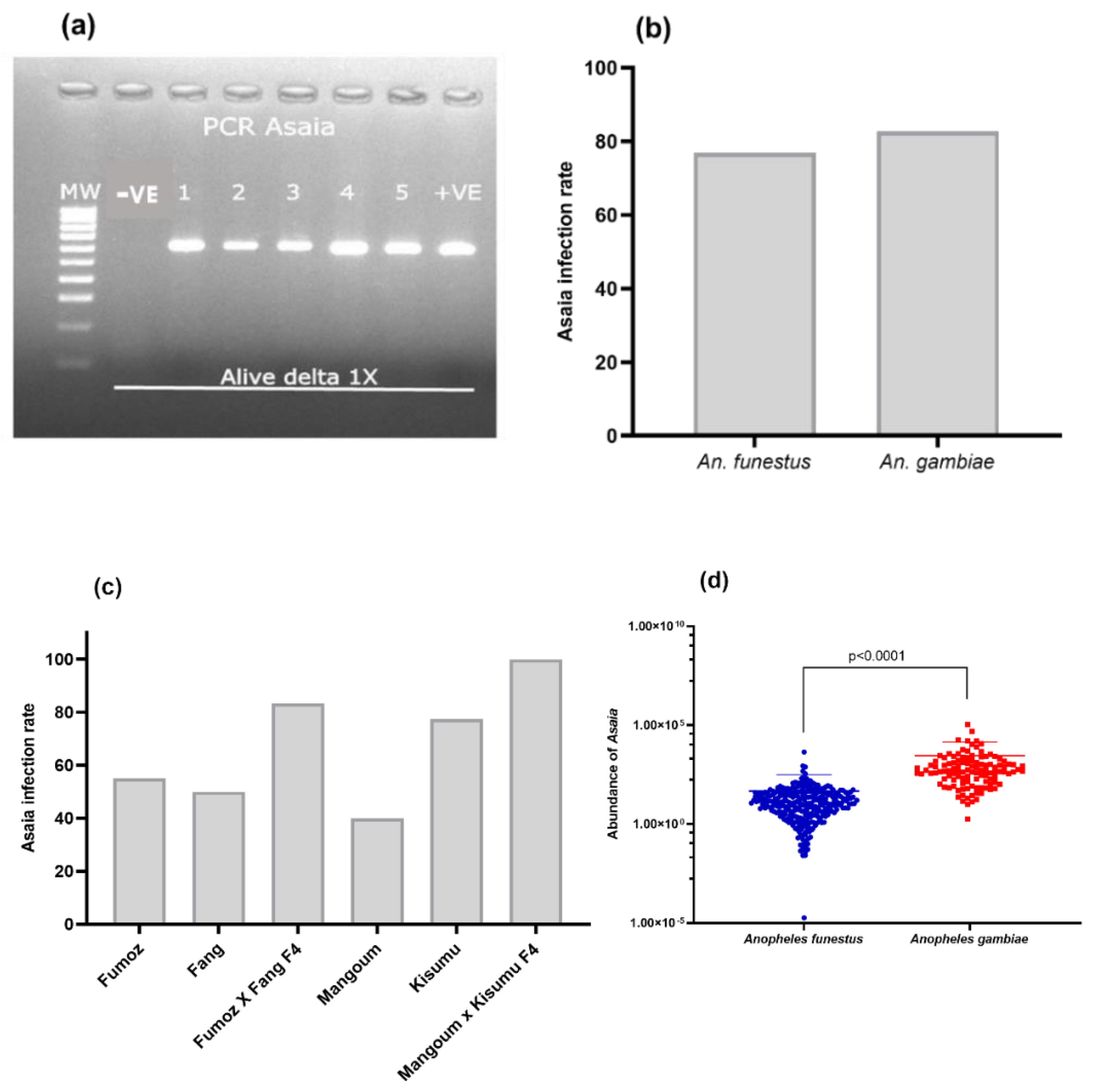
3.4.1. Prevalence of Asaia spp. in Anopheles Strains
Assessment of the Link between the Prevalence of Asaia spp. and Insecticide Resistance in Hybrids FUMOZ_X_FANG
Association between the Prevalence of Asaia spp. and Insecticide Resistance in Hybrids MANGOUM_X_KISUMU
3.4.2. Abundance of Asaia spp. in Anopheles Hybrids Strains
Assessment of the Link between the Quantity of Asaia spp. and Insecticide Resistance in Hybrids FUMOZ_X_FANG
Association between Asaia spp. Load and Insecticide Resistance in Anopheles gambiae Hybrid Strains
4. Discussion
4.1. High Level of Resistance to Pyrethroids in An. funestus and An. gambiae Strains
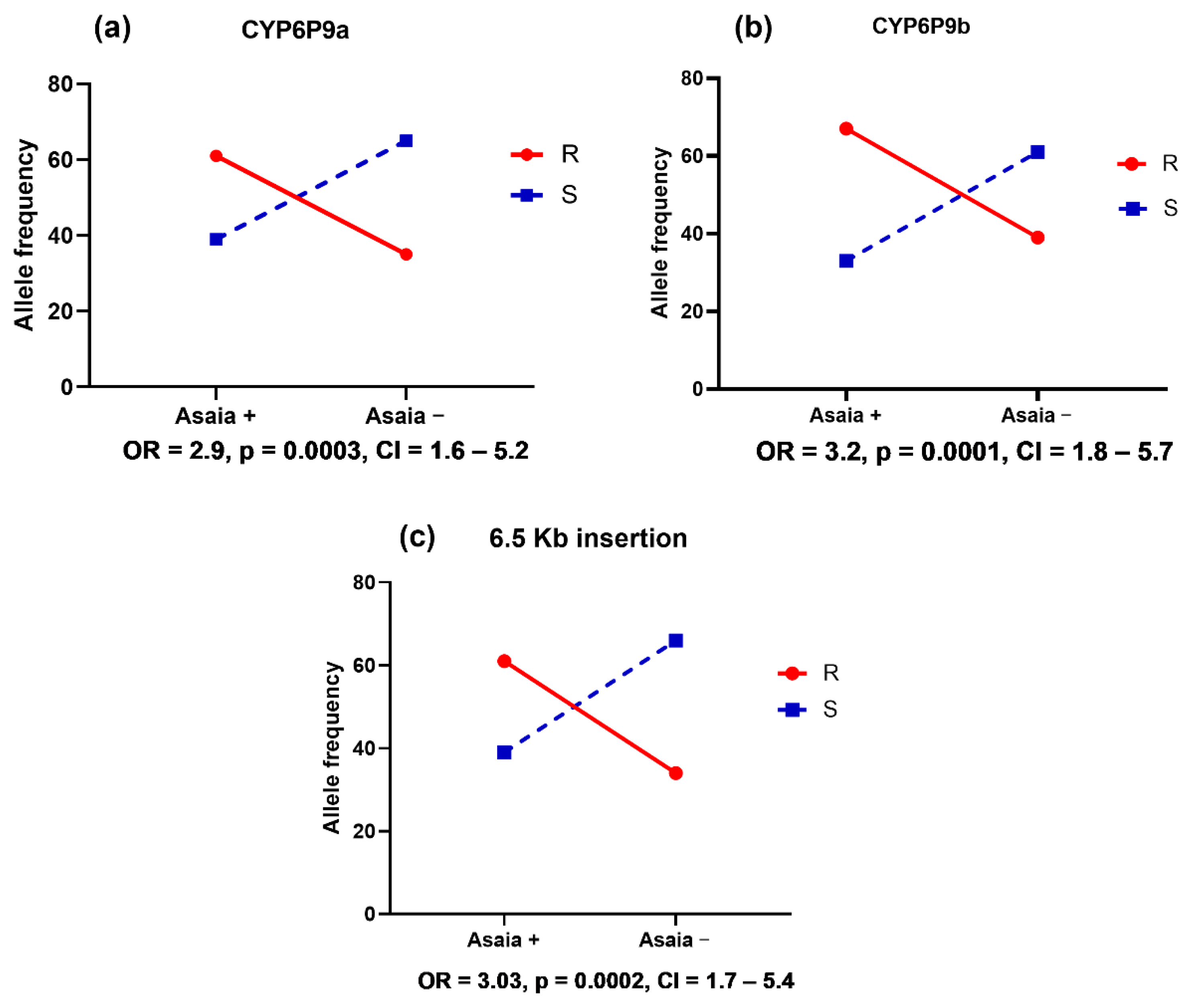
4.2. Cytochrome P450 Markers and VGSC Mutations Are Partially Associated with the Escalation of Pyrethroid Resistance in Mosquitoes
4.3. Detection of Asaia spp. in Anopheles Strains
4.4. Asaia spp. Is Associated with the Escalation of Pyrethroid Resistance in An. funestus
4.5. Asaia Load Is Negatively Associated with the Pyrethroid Resistance in Anopheles gambiae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- World Health Organization. Malaria Vaccine Implementation, P. Full Evidence Report on the RTS,S/AS01 Malaria Vaccine; WHO: Geneva, Switzerland, 2021; p. 90. [Google Scholar]
- Bhatt, S.; Weiss, D.J.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.E.; Moyes, C.L.; Henry, A.; Eckhoff, P.A.; et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Hemingway, J. The way forward for vector control. Science 2017, 358, 998–999. [Google Scholar] [CrossRef]
- Menze, B.D.; Kouamo, M.F.; Wondji, M.J.; Tchapga, W.; Tchoupo, M.; Kusimo, M.O.; Mouhamadou, C.S.; Riveron, J.M.; Wondji, C.S. An Experimental Hut Evaluation of PBO-Based and Pyrethroid-Only Nets against the Malaria Vector Anopheles funestus Reveals a Loss of Bed Nets Efficacy Associated with GSTe2 Metabolic Resistance. Genes 2020, 11, 143. [Google Scholar] [CrossRef] [Green Version]
- Tchouakui, M.; Mugenzi, L.M.J.; Menze, B.D.; Khaukha, J.N.T.; Tchapga, W.; Tchoupo, M.; Wondji, M.J.; Wondji, C.S. Pyrethroid Resistance Aggravation in Ugandan Malaria Vectors Is Reducing Bednet Efficacy. Pathogens 2021, 10, 415. [Google Scholar] [CrossRef]
- Mugenzi, L.M.; Akosah-Brempong, G.; Tchouakui, M.; Menze, B.D.; Tekoh, T.A.; Tchoupo, M.; Nkemngo, F.N.; Wondji, M.J.; Nwaefuna, E.K.; Osae, M.; et al. Escalating pyrethroid resistance in two major malaria vectors Anopheles funestus and Anopheles gambiae (s.l.) in Atatam, Southern Ghana. BMC Infect. Dis. 2022, 22, 799. [Google Scholar] [CrossRef]
- Toé, K.H.; Jones, C.M.; N’Fale, S.; Ismail, H.M.; Dabiré, R.K.; Ranson, H. Increased Pyrethroid Resistance in Malaria Vectors and Decreased Bed Net Effectiveness, Burkina Faso. Emerg. Infect. Dis. 2014, 20, 1691. [Google Scholar] [CrossRef]
- Riveron, J.M.; Huijben, S.; Tchapga, W.; Tchouakui, M.; Wondji, M.J.; Tchoupo, M.; Irving, H.; Cuamba, N.; Maquina, M.; Paaijmans, K.; et al. Escalation of Pyrethroid Resistance in the Malaria Vector Anopheles funestus Induces a Loss of Efficacy of Piperonyl Butoxide–Based Insecticide-Treated Nets in Mozambique. J. Infect. Dis. 2019, 220, 467–475. [Google Scholar] [CrossRef] [Green Version]
- Corbel, V.; N’Guessan, R. Distribution, Mechanisms, Impact and Management of Insecticide Resistance in Malaria Vectors: A Pragmatic Review. In Anopheles Mosquitoes–New Insights into Malaria Vectors; Manguin, S., Ed.; InTech: Tokyo, Japan, 2013. [Google Scholar]
- Riveron, J.M.; Tchouakui, M.; Mugenzi, L.; Menze, B.D.; Chiang, M.-C.; Wondji, C.S. Insecticide Resistance in Malaria Vectors: An Update at a Global Scale. In Towards Malaria Elimination—A Leap Forward; Manguin, S., Dev, V., Eds.; InTech: Tokyo, Japan, 2018. [Google Scholar]
- Omoke, D.; Kipsum, M.; Otieno, S.; Esalimba, E.; Sheth, M.; Lenhart, A.; Njeru, E.M.; Ochomo, E.; Dada, N. Western Kenyan Anopheles gambiae showing intense permethrin resistance harbour distinct microbiota. Malar. J. 2021, 20, 77. [Google Scholar] [CrossRef]
- Pelloquin, B.; Kristan, M.; Edi, C.; Meiwald, A.; Clark, E.; Jeffries, C.L.; Walker, T.; Dada, N.; Messenger, L.A. Overabundance of Asaia and Serratia Bacteria Is Associated with Deltamethrin Insecticide Susceptibility in Anopheles coluzzii from Agboville, Côte d’Ivoire. Microbiol. Spectr. 2021, 9, e00157-21. [Google Scholar] [CrossRef]
- Dada, N.; Lol, J.C.; Benedict, A.C.; López, F.; Sheth, M.; Dzuris, N.; Padilla, N.; Lenhart, A. Pyrethroid exposure alters internal and cuticle surface bacterial communities in Anopheles albimanus. ISME J. 2019, 13, 2447–2464. [Google Scholar] [CrossRef] [Green Version]
- Ingham, V.A.; Tennessen, J.A.; Lucas, E.R.; Elg, S.; Yates, H.C.; Carson, J.; Guelbeogo, W.M.; Sagnon, N.F.; Hughes, G.L.; Heinz, E.; et al. Integration of whole genome sequencing and transcriptomics reveals a complex picture of the reestablishment of insecticide resistance in the major malaria vector Anopheles coluzzii. PLoS Genet. 2021, 17, e1009970. [Google Scholar] [CrossRef]
- Douglas, A.E. Multiorganismal Insects: Diversity and Function of Resident Microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Gendrin, M.; Christophides, G.K. The Anopheles Mosquito Microbiota and Their Impact on Pathogen Transmission. In Anopheles Mosquitoes—New Insights into Malaria Vectors; Manguin, S., Ed.; InTech: Tokyo, Japan, 2013. [Google Scholar]
- Bahia, A.C.; Dong, Y.; Blumberg, B.J.; Mlambo, G.; Tripathi, A.; BenMarzouk-Hidalgo, O.J.; Chandra, R.; Dimopoulos, G. Exploring Anopheles gut bacteria for Plasmodium blocking activity. Environ. Microbiol. 2014, 16, 2980–2994. [Google Scholar] [CrossRef] [Green Version]
- Cirimotich, C.M.; Dong, Y.; Clayton, A.M.; Sandiford, S.L.; Souza-Neto, J.A.; Mulenga, M.; Dimopoulos, G. Natural microbe-mediated refractoriness to Plasmodium infection in Anopheles gambiae. Science 2011, 332, 855–858. [Google Scholar] [CrossRef] [Green Version]
- Meister, S.; Agianian, B.; Turlure, F.; Relógio, A.; Morlais, I.; Kafatos, F.C.; Christophides, G.K. Anopheles gambiae PGRPLC-Mediated Defense against Bacteria Modulates Infections with Malaria Parasites. PLoS Pathog. 2009, 5, e1000542. [Google Scholar] [CrossRef] [Green Version]
- Pietri, J.E.; Liang, D. The Links Between Insect Symbionts and Insecticide Resistance: Causal Relationships and Physiological Tradeoffs. Ann. Entomol. Soc. Am. 2018, 111, 92–97. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Hayatsu, M.; Hosokawa, T.; Nagayama, A.; Tago, K.; Fukatsu, T. Symbiont-mediated insecticide resistance. Proc. Natl. Acad. Sci. USA 2012, 109, 8618–8622. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Sun, B.; Gurr, G.M.; Vasseur, L.; Xue, M.; You, M. Gut Microbiota Mediate Insecticide Resistance in the Diamondback Moth, Plutella xylostella (L.). Front. Microbiol. 2018, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Dada, N.; Sheth, M.; Liebman, K.; Pinto, J.; Lenhart, A. Whole metagenome sequencing reveals links between mosquito microbiota and insecticide resistance in malaria vectors. Sci. Rep. 2018, 8, 2084. [Google Scholar] [CrossRef] [Green Version]
- Comandatore, F.; Damiani, C.; Cappelli, A.; Ribolla, P.E.M.; Gasperi, G.; Gradoni, F.; Capelli, G.; Piazza, A.; Montarsi, F.; Mancini, M.V.; et al. Phylogenomics Reveals that Asaia Symbionts from Insects Underwent Convergent Genome Reduction, Preserving an Insecticide-Degrading Gene. mBio 2021, 12, e00106-21. [Google Scholar] [CrossRef]
- Favia, G.; Ricci, I.; Marzorati, M.; Negri, I.; Alma, A.; Sacchi, L.; Bandi, C.; Daffonchio, D. Bacteria of the Genus Asaia: A Potential Paratransgenic Weapon Against Malaria. In Transgenesis and the Management of Vector-Borne Disease; Aksoy, S., Ed.; Springer: New York, NY, USA, 2008; Volume 627, pp. 49–59. [Google Scholar]
- Moran, N.A.; Dunbar, H.E. Sexual acquisition of beneficial symbionts in aphids. Proc. Natl. Acad. Sci. USA 2006, 103, 12803–12806. [Google Scholar] [CrossRef] [Green Version]
- Bassene, H.; Niang, E.H.A.; Fenollar, F.; Doucoure, S.; Faye, O.; Raoult, D.; Sokhna, C.; Mediannikov, O. Role of plants in the transmission of Asaia sp., which potentially inhibit the Plasmodium sporogenic cycle in Anopheles mosquitoes. Sci. Rep. 2020, 10, 7144. [Google Scholar] [CrossRef]
- Hunt, R.H.; Brooke, B.D.; Pillay, C.; Koekemoer, L.L.; Coetzee, M. Laboratory selection for and characteristics of pyrethroid resistance in the malaria vector Anopheles funestus. Med. Vet. Entomol. 2005, 19, 271–275. [Google Scholar] [CrossRef]
- Hargreaves, K.; Koekemoer, L.L.; Brooke, B.D.; Hunt, R.H.; Mthembu, J.; Coetzee, M. Anopheles funestus resistant to pyrethroid insecticides in South Africa. Med. Vet. Entomol. 2000, 14, 181–189. [Google Scholar] [CrossRef]
- Besansky, N.J.; Stump, A.D.; Vulule, J.M.; Atieli, F.K. Dynamics of the Pyrethroid Knockdown Resistance Allele in Western Kenyan Populations of Anopheles Gambiae in Response to Insecticide-Treated Bed Net Trials. Am. J. Trop. Med. Hyg. 2004, 70, 591–596. [Google Scholar] [CrossRef]
- Tepa, A.; Kengne-Ouafo, J.A.; Djova, V.S.; Tchouakui, M.; Mugenzi, L.M.J.; Djouaka, R.; Pieme, C.A.; Wondji, C.S. Molecular Drivers of Multiple and Elevated Resistance to Insecticides in a Population of the Malaria Vector Anopheles gambiae in Agriculture Hotspot of West Cameroon. Genes 2022, 13, 1206. [Google Scholar] [CrossRef]
- World Health Organization; Food and Agriculture Organization of the United Nations. Global Situation of Pesticide Management in Agriculture and Public Health: Report of a 2018 WHO-FAO Survey; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Livak, K.J. Organization and Mapping of A Sequence on the Drosophila Melanogaster X and Y Chromosomes That is Transcribed During Spermatogenesis. Genetics 1984, 107, 611–634. [Google Scholar] [CrossRef]
- Weedall, G.D.; Mugenzi, L.M.J.; Menze, B.D.; Tchouakui, M.; Ibrahim, S.S.; Amvongo-Adjia, N.; Irving, H.; Wondji, M.J.; Tchoupo, M.; Djouaka, R.; et al. A cytochrome P450 allele confers pyrethroid resistance on a major African malaria vector, reducing insecticide-treated bednet efficacy. Sci. Transl. Med. 2019, 11, eaat7386. [Google Scholar] [CrossRef] [Green Version]
- Mugenzi, L.M.J.; Menze, B.D.; Tchouakui, M.; Wondji, M.J.; Irving, H.; Tchoupo, M.; Hearn, J.; Weedall, G.D.; Riveron, J.M.; Wondji, C.S. Cis-regulatory CYP6P9b P450 variants associated with loss of insecticide-treated bed net efficacy against Anopheles funestus. Nat. Commun. 2019, 10, 4652. [Google Scholar] [CrossRef] [Green Version]
- Mugenzi, L.M.J.; Menze, B.D.; Tchouakui, M.; Wondji, M.J.; Irving, H.; Tchoupo, M.; Hearn, J.; Weedall, G.D.; Riveron, J.M.; Cho-Ngwa, F.; et al. A 6.5-kb intergenic structural variation enhances P450-mediated resistance to pyrethroids in malaria vectors lowering bed net efficacy. Mol. Ecol. 2020, 29, 4395–4411. [Google Scholar] [CrossRef]
- Bass, C.; Nikou, D.; Blagborough, A.M.; Vontas, J.; Sinden, R.E.; Williamson, M.S.; Field, L.M. PCR-based detection of Plasmodium in Anopheles mosquitoes: A comparison of a new high-throughput assay with existing methods. Malar. J. 2008, 7, 177. [Google Scholar] [CrossRef] [Green Version]
- Rami, A.; Raz, A.; Zakeri, S.; Dinparast Djadid, N. Isolation and identification of Asaia sp. in Anopheles spp. mosquitoes collected from Iranian malaria settings: Steps toward applying paratransgenic tools against malaria. Parasites Vectors 2018, 11, 367. [Google Scholar] [CrossRef] [Green Version]
- Jeffries, C.L.; Lawrence, G.G.; Golovko, G.; Kristan, M.; Orsborne, J.; Spence, K.; Hurn, E.; Bandibabone, J.; Tantely, M.L.; Raharimalala, F.N.; et al. Novel Wolbachia Strains in Anopheles Malaria Vectors from Sub-Saharan Africa. bioRxiv 2018. [Google Scholar] [CrossRef]
- Wondji, C.S.; Irving, H.; Morgan, J.; Lobo, N.F.; Collins, F.H.; Hunt, R.H.; Coetzee, M.; Hemingway, J.; Ranson, H. Two duplicated P450 genes are associated with pyrethroid resistance in Anopheles funestus, a major malaria vector. Genome Res. 2009, 19, 452–459. [Google Scholar] [CrossRef] [Green Version]
- Riveron, J.M.; Irving, H.; Ndula, M.; Barnes, K.G.; Ibrahim, S.S.; Paine, M.J.I.; Wondji, C.S. Directionally selected cytochrome P450 alleles are driving the spread of pyrethroid resistance in the major malaria vector Anopheles funestus. Proc. Natl. Acad. Sci. USA 2013, 110, 252–257. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.; Flood, L.; Praulins, G.; Ingham, V.A.; Morgan, J.; Lees, R.S.; Ranson, H. Characterisation of Anopheles strains used for laboratory screening of new vector control products. Parasites Vectors 2019, 12, 522. [Google Scholar] [CrossRef] [Green Version]
- Machani, M.G.; Ochomo, E.; Zhong, D.; Zhou, G.; Wang, X.; Githeko, A.K.; Yan, G.; Afrane, Y.A. Phenotypic, genotypic and biochemical changes during pyrethroid resistance selection in Anopheles gambiae mosquitoes. Sci. Rep. 2020, 10, 19063. [Google Scholar] [CrossRef]
- Tchouakui, M.; Oruni, A.; Assatse, T.; Manyaka, C.R.; Tchoupo, M.; Kayondo, J.; Wondji, C.S. Fitness cost of target-site and metabolic resistance to pyrethroids drives restoration of susceptibility in a highly resistant Anopheles gambiae population from Uganda. PLoS ONE 2022, 17, e0271347. [Google Scholar] [CrossRef]
- Brooke, B.D.; Kloke, G.; Hunt, R.H.; Koekemoer, L.L.; Tem, E.A.; Taylor, M.E.; Small, G.; Hemingway, J.; Coetzee, M. Bioassay and biochemical analyses of insecticide resistance in southern African Anopheles funestus (Diptera: Culicidae). Bull. Entomol. Res. 2001, 91, 265–272. [Google Scholar] [CrossRef]
- Amenya, D.A.; Naguran, R.; Lo, T.C.M.; Ranson, H.; Spillings, B.L.; Wood, O.R.; Brooke, B.D.; Coetzee, M.; Koekemoer, L.L. Over expression of a Cytochrome P450 (CYP6P9) in a Major African Malaria Vector, Anopheles funestus, Resistant to Pyrethroids: CYP6P9 overexpressed in resistant An. funestus. Insect Mol. Biol. 2008, 17, 19–25. [Google Scholar] [CrossRef]
- Riveron, J.M.; Ibrahim, S.S.; Chanda, E.; Mzilahowa, T.; Cuamba, N.; Irving, H.; Barnes, K.G.; Ndula, M.; Wondji, C.S. The highly polymorphic CYP6M7 cytochrome P450 gene partners with the directionally selected CYP6P9a and CYP6P9b genes to expand the pyrethroid resistance front in the malaria vector Anopheles funestus in Africa. BMC Genom. 2014, 15, 817. [Google Scholar] [CrossRef] [Green Version]
- Riveron, J.M.; Chiumia, M.; Menze, B.D.; Barnes, K.G.; Irving, H.; Ibrahim, S.S.; Weedall, G.D.; Mzilahowa, T.; Wondji, C. S. Rise of multiple insecticide resistance in Anopheles funestus in Malawi: A major concern for malaria vector control. Malar. J. 2015, 14, 344. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, S.S.; Riveron, J.M.; Bibby, J.; Irving, H.; Yunta, C.; Paine, M.J.I.; Wondji, C.S. Allelic Variation of Cytochrome P450s Drives Resistance to Bednet Insecticides in a Major Malaria Vector. PLoS Genet 2015, 11, e1005618. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Torres, D.; Chandre, F.; Williamson, M.S.; Darriet, F.; Berge, J.B.; Devonshire, A.L.; Guillet, P.; Pasteur, N.; Pauron, D. Molecular characterization of pyrethroid knockdown resistance (kdr) in the major malaria vector Anopheles gambiae s.s. Insect Mol. Biol. 1998, 7, 179–184. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly, A.O.; Khambay, B.P.S.; Williamson, M.S.; Field, L.M.; Wallace, B.A.; Davies, T.G.E. Modelling insecticide-binding sites in the voltage-gated sodium channel. Biochem. J. 2006, 396, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Verhaeghen, K.; Van Bortel, W.; Roelants, P.; Backeljau, T.; Coosemans, M. Detection of the East and West African kdr mutation in Anopheles gambiae and Anopheles arabiensis from Uganda using a new assay based on FRET/Melt Curve analysis. Malar. J. 2006, 5, 16. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.M.; Liyanapathirana, M.; Agossa, F.R.; Weetman, D.; Ranson, H.; Donnelly, M.J.; Wilding, C.S. Footprints of positive selection associated with a mutation (N1575Y) in the voltage-gated sodium channel of Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2012, 109, 6614–6619. [Google Scholar] [CrossRef] [Green Version]
- Chouaia, B.; Rossi, P.; Montagna, M.; Ricci, I.; Crotti, E.; Damiani, C.; Epis, S.; Faye, I.; Sagnon, N.F.; Alma, A.; et al. Molecular Evidence for Multiple Infections as Revealed by Typing of Asaia Bacterial Symbionts of Four Mosquito Species. Appl. Environ. Microbiol. 2010, 76, 7444–7450. [Google Scholar] [CrossRef] [Green Version]
- Damiani, C.; Ricci, I.; Crotti, E.; Rossi, P.; Rizzi, A.; Scuppa, P.; Capone, A.; Ulissi, U.; Epis, S.; Genchi, M.; et al. Mosquito-Bacteria Symbiosis: The Case of Anopheles gambiae and Asaia. Microb. Ecol. 2010, 60, 644–654. [Google Scholar] [CrossRef] [Green Version]
- Damiani, C.; Ricci, I.; Crotti, E.; Rossi, P.; Rizzi, A.; Scuppa, P.; Esposito, F.; Bandi, C.; Daffonchio, D.; Favia, G. Paternal transmission of symbiotic bacteria in malaria vectors. Curr. Biol. 2008, 18, R1087–R1088. [Google Scholar] [CrossRef] [Green Version]
- Ricci, I.; Damiani, C.; Rossi, P.; Capone, A.; Scuppa, P.; Cappelli, A.; Ulissi, U.; Mosca, M.; Valzano, M.; Epis, S.; et al. Mosquito symbioses: From basic research to the paratransgenic control of mosquito-borne diseases: Bacterial symbiosis in mosquito. J. Appl. Entomol. 2011, 135, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Soltani, A.; Vatandoost, H.; Oshaghi, M.A.; Enayati, A.A.; Chavshin, A.R. The role of midgut symbiotic bacteria in resistance of Anopheles stephensi (Diptera: Culicidae) to organophosphate insecticides. Pathog. Glob. Health 2017, 111, 289–296. [Google Scholar] [CrossRef]
- Zhang, J.; Pan, Y.; Zheng, C.; Gao, X.; Wei, X.; Xi, J.; Peng, T.; Shang, Q. Rapid evolution of symbiotic bacteria populations in spirotetramat-resistant Aphis gossypii glover revealed by pyrosequencing. Comp. Biochem. Physiol. Part D Genom. Proteom. 2016, 20, 151–158. [Google Scholar] [CrossRef]
- Berticat, C.; Rousset, F.; Raymond, M.; Berthomieu, A.; Weill, M. High Wolbachia density in insecticide-resistant mosquitoes. Proc. R. Soc. London. Ser. B Biol. Sci. 2002, 269, 1413–1416. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.; Wang, L.; Guo, W.; Zhang, X.; Peng, D.; Luo, C.; Yu, Z.; Sun, M. Bacillus thuringiensis bel protein enhances the toxicity of Cry1Ac protein to Helicoverpa armigera larvae by degrading insect intestinal mucin. Appl. Environ. Microbiol. 2009, 75, 5237–5243. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
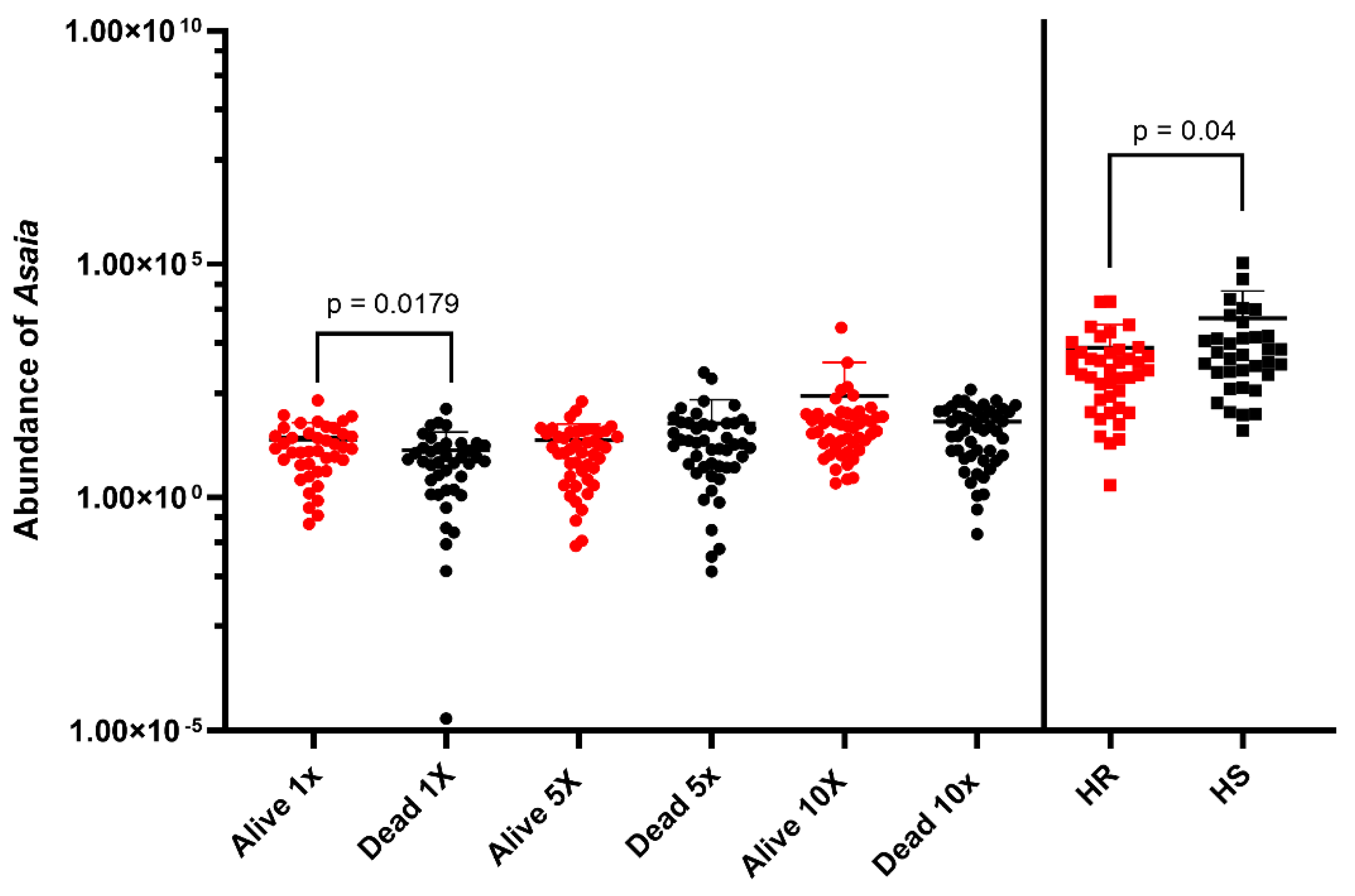
| Species | Phenotypes | Tested | Infected | Prevalence of Asaia spp. | Comparison of the Prevalence of Asaia spp. between Alive and Dead |
|---|---|---|---|---|---|
| An. funestus | Alive 1X | 42 | 41 | 97.62 | χ2 = 4.91, p = 0.02; OR = 8.2 p = 0.05 |
| Dead 1X | 42 | 35 | 83.33 | ||
| Alive 5X | 48 | 47 | 97.92 | χ2 = 16.93, p < 0.0001; OR = 25.8, p = 0.002 | |
| Dead 5X | 48 | 31 | 64.58 | ||
| Alive 10X | 48 | 48 | 100 | χ2 = 4.11, p = 0.04; OR = 9.8, p = 0.12 | |
| Dead 10X | 48 | 44 | 91.67 | ||
| An. gambiae | HR | 42 | 42 | 100 | / |
| HS | 35 | 35 | 100 |
| Markers | Genotype | Tested | Median | Mean | Percentiles (IQR) | |
|---|---|---|---|---|---|---|
| 25th | 75th | |||||
| L1014F | RR | 28 | 569.16 | 1832.8 | 142.19 | 1205.1 |
| RS | 23 | 484.32 | 1457.2 | 145.24 | 1970.4 | |
| SS | 23 | 1281.7 | 9393.1 | 432.8 | 5703.5 | |
| N1575Y | RS | 37 | 587.57 | 2452 | 276.09 | 2199.2 |
| SS | 38 | 873.93 | 5570.7 | 120.4 | 1784.2 | |
| L1014F and N1575Y | RR/RS | 21 | 550.76 | 2103.6 | 229.51 | 1074.2 |
| RS/RS | 12 | 1227.4 | 2354.1 | 265.27 | 4033 | |
| SS/SS | 18 | 1281.7 | 10,407 | 195.91 | 2887 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djondji Kamga, F.M.; Mugenzi, L.M.J.; Tchouakui, M.; Sandeu, M.M.; Maffo, C.G.T.; Nyegue, M.A.; Wondji, C.S. Contrasting Patterns of Asaia Association with Pyrethroid Resistance Escalation between the Malaria Vectors Anopheles funestus and Anopheles gambiae. Microorganisms 2023, 11, 644. https://doi.org/10.3390/microorganisms11030644
Djondji Kamga FM, Mugenzi LMJ, Tchouakui M, Sandeu MM, Maffo CGT, Nyegue MA, Wondji CS. Contrasting Patterns of Asaia Association with Pyrethroid Resistance Escalation between the Malaria Vectors Anopheles funestus and Anopheles gambiae. Microorganisms. 2023; 11(3):644. https://doi.org/10.3390/microorganisms11030644
Chicago/Turabian StyleDjondji Kamga, Fleuriane Metissa, Leon M. J. Mugenzi, Magellan Tchouakui, Maurice Marcel Sandeu, Claudine Grace Tatsinkou Maffo, Maximilienne Ascension Nyegue, and Charles S. Wondji. 2023. "Contrasting Patterns of Asaia Association with Pyrethroid Resistance Escalation between the Malaria Vectors Anopheles funestus and Anopheles gambiae" Microorganisms 11, no. 3: 644. https://doi.org/10.3390/microorganisms11030644