
Bacterial Pigments and Their Multifaceted Roles in Contemporary Biotechnology and Pharmacological Applications
 , ,
, ,  and
and
Abstract
:1. Introduction
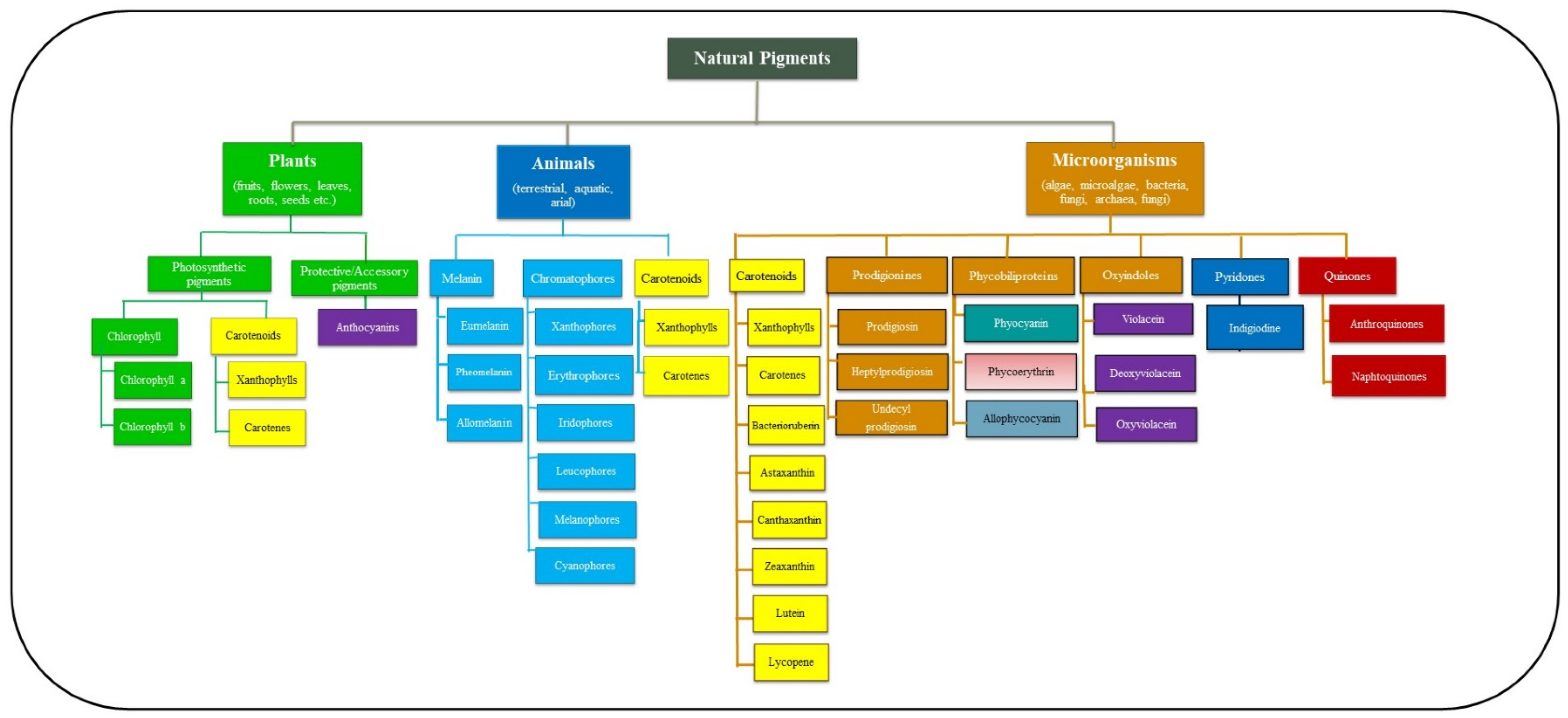
2. Overview of Natural Pigments, Plant Pigments and Microbial Pigments
3. Ecological Distribution of Microbial Pigments
4. Advantages of Microbial Pigments over Synthetic Pigments
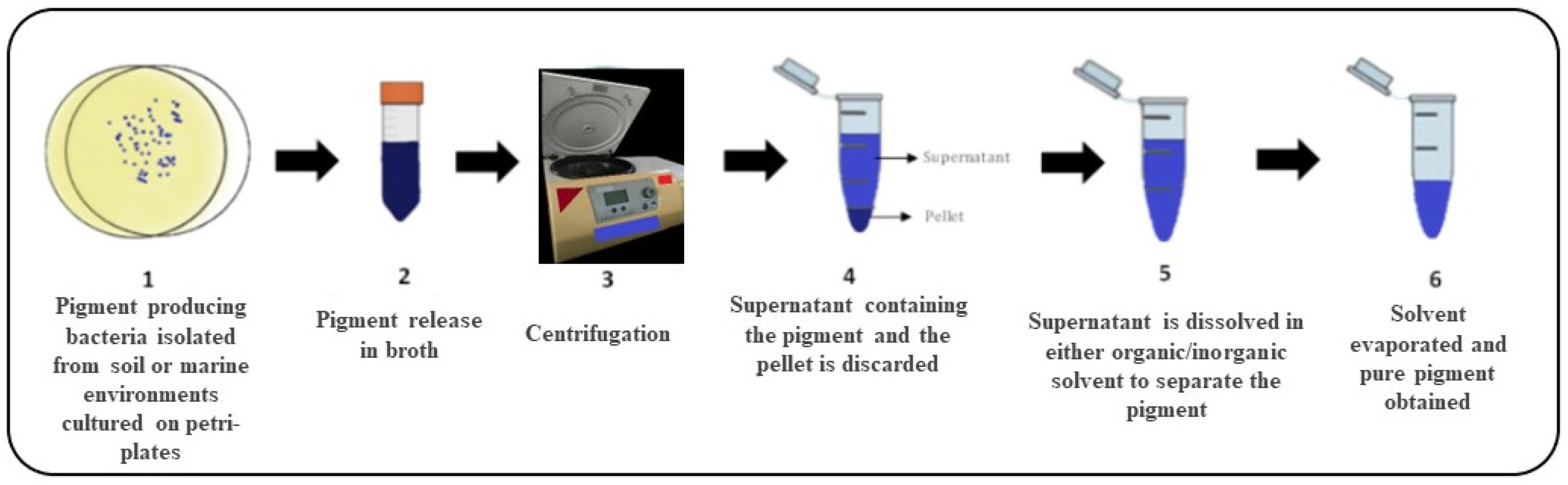
5. Pigment Production from Bacteria and Genes Involved in Pigment Production
6. The Utilization of Genome Engineering Techniques to Enhance Bacterial Pigment Production
7. Application of the Bacterial Pigments in the Pharmaceutical Industry
7.1. Anti-Tumour Effect of Bacterial Pigments
7.2. Antimicrobial Action of Bacterial Pigments
7.3. Antiviral Activity of Bacterial Pigments
7.4. Antifungal Activity of Bacterial Pigments
7.5. Antiparasitic Activity of Bacterial Pigments
7.6. Bacterial Pigments as Immunosuppressors
8. Application of Microbial Pigments in Food Industries
9. Application of Microbial Pigments in the Cosmetic Industry
10. Application of Microbial Pigments in Textile Industry
11. Structure and Bioactivity of Microbial Pigments
12. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kant, R. Textile dyeing industry an environmental hazard. Nat. Sci. 2012, 4, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Tkaczyk, A.; Mitrowska, K.; Posyniak, A. Synthetic organic dyes as contaminants of the aquatic environment and their implications for ecosystems: A review. Sci. Total. Environ. 2020, 717, 137222. [Google Scholar] [CrossRef] [PubMed]
- Dikshit, R.; Tallapragada, P. Comparative Study of Natural and Artificial Flavoring Agents and Dyes. In Natural and Artificial Flavoring Agents and Food Dyes; Academic Press: Cambridge, MA, USA, 2018; pp. 83–111. [Google Scholar] [CrossRef]
- Sutthiwong, N.; Fouillaud, M.; Valla, A.; Caro, Y.; Dufossé, L. Bacteria belonging to the extremely versatile genus Arthrobacter as novel source of natural pigments with extended hue range. Food Res. Int. 2014, 65, 156–162. [Google Scholar] [CrossRef]
- Usman, H.M.; Abdulkadir, N.; Gani, M.; Maiturare, H.M. Bacterial pigments and its significance. MOJ Bioequiv Availab. 2017, 4, 285–288. [Google Scholar] [CrossRef] [Green Version]
- AMR. 2021. Available online: https://www.alliedmarketresearch.com/carotenoids-market (accessed on 21 January 2021).
- Mussagy, C.U.; Khan, S.; Kot, A.M. Current developments on the application of microbial carotenoids as an alternative to synthetic pigments. Crit. Rev. Food Sci. Nutr. 2021, 62, 6932–6946. [Google Scholar] [CrossRef]
- Ram, S.; Mitra, M.; Shah, F.; Tirkey, S.R.; Mishra, S. Bacteria as an alternate biofactory for carotenoid production: A review of its applications, opportunities and challenges. J. Funct. Food 2020, 67, 103867. [Google Scholar] [CrossRef]
- Usmani, Z.; Sharma, M.; Sudheer, S.; Gupta, V.K.; Bhat, R. Engineered Microbes for Pigment Production Using Waste Biomass. Curr. Genom. 2020, 21, 80–95. [Google Scholar] [CrossRef]
- Jaradat, A.; Al-Akhras MA, H.; Makhadmeh, G.; Aljarrah, K.; Al-Omari, A.; Ababneh, Z.; Alshorman, M. Artificial semi-rigid sensitized with natural pigments: Effect of photon radiations. J. Pharm. BioAllied Sci. 2011, 3, 266–276. [Google Scholar] [CrossRef]
- Mordini, E.; Ashton, H. The Transparent Body: Medical Information, Physical Privacy and Respect for Body Integrity. In Second Generation Biometrics: The Ethical, Legal and Social Context. The International Library of Ethics, Law and Technology; Mordini, E., Tzovaras, D., Eds.; Springer: Dordrecht, The Netherlands, 2012; Volume 11. [Google Scholar] [CrossRef]
- Fingerman, M. Chromatophores. Physiol. Rev. 1965, 45, 296–339. [Google Scholar] [CrossRef]
- Rudh, A.; Qvarnström, A. Adaptive colouration in amphibians. Semin. Cell Dev. Biol. 2013, 24, 553–561. [Google Scholar] [CrossRef]
- Price, A.C.; Weadick, C.J.; Shim, J.; Rodd, F.H. Pigments, Patterns, and Fish Behavior. Zebrafish 2008, 5, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, J.; Grimalt, J.O.; de Wit, R.; Keely, B.J.; Maxwell, J.R. Chlorophyll and carotenoid pigments in solar saltern microbial mats. Geochim. Cosmochim. Acta 1194, 58, 4703–4715. [Google Scholar] [CrossRef]
- Rajagopal, L.; Sundari, S.C.; Balasubramanian, D.; Sonti, R. The bacterial pigment xanthomonadin offers protection against photodamage. FEBS Lett. 1997, 415, 125–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, T.; Barrow, C.J.; Deshmukh, S.K. Microbial Pigments in the Food Industry—Challenges and the Way Forward. Front. Nutr. 2019, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambati, R.R.; Siew Moi, P.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications—A review. Marine Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef]
- Galasso, C.; Corinaldesi, C.; Sansone, C. Carotenoids from Marine Organisms: Biological Functions and Industrial Applications. Antioxidants 2017, 6, 96. [Google Scholar] [CrossRef] [Green Version]
- Cappelletti, M.; Presentato, A.; Piacenza, E.; Firrincieli, A.; Turner, R.J.; Zannoni, D. Biotechnology of Rhodococcus for the production of valuable compounds. Appl. Microbiol. Biotechnol. 2020, 104, 8567–8594. [Google Scholar] [CrossRef]
- Alshamaa, D.S.; Issam, M.M. The Role of Extracted Carotenoid from Staphylococci as Antioxidant and Antibacterial. Rafidain J. Sci. 2017, 26, 17–24. [Google Scholar] [CrossRef]
- Tian, B.; Sun, Z.; Shen, S.; Wang, H.; Jiao, J.; Wang, L.; Hu, Y. Effects of carotenoids from Deinococcus radiodurans on protein oxidation. Lett. Appl. Microbiol. 2009, 49, 689–694. [Google Scholar] [CrossRef]
- Williamson, N.R. Biosynthesis of the red antibiotic, prodigiosin, in Serratia: Identification of a novel 2-methyl-3-n-amyl-pyrrole (MAP) assembly pathway, definition of the terminal condensing enzyme, and implications for undecylprodigiosin biosynthesis in Streptomyces. Mol. Microbiol. 2005, 56, 971–989. [Google Scholar] [CrossRef]
- Stankovic, N.; Senerovic, L.; Ilic-Tomic, T.; Vasiljevic, B.; Nikodinovic-Runic, J. Properties and applications of undecylprodigiosin and other bacterial prodigiosins. Appl. Microbiol. Biotechnol. 2014, 98, 3841–3858. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Liu, G.; Li, J.; Huang, H.; Zhang, X.; Zhang, H.; Ju, J. Cytotoxic and Antibacterial Angucycline- and Prodigiosin- Analogues from the Deep-Sea Derived Streptomyces sp. SCSIO 11594. Mar. Drugs 2015, 13, 1304–1316. [Google Scholar] [CrossRef] [PubMed]
- Lazaro, J.E.; Nitcheu, J.; Predicala, R.Z.; Mangalindan, G.C.; Nesslany, F.; Marzin, D.; Diquet, B. Heptyl prodigiosin, a bacterial metabolite, is antimalarial in vivo and non-mutagenic in vitro. J. Nat. Toxins 2002, 11, 367–377. [Google Scholar] [PubMed]
- Baront, S.S.; Rowe, J.J. Antibiotic Action of Pyocyanin. Antimicrob. Agents Chemother. 1981, 20, 814–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Justo, G.Z.; Durán, N.; Ferreira, C.V.; Melon, P.S.; Cordi, L.; Martins, D. Violacein: Properties and biological activities. Biotechnol. Appl. Biochem. 2007, 48, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Andrighetti-Fröhner, C.R.; Antonio, R.V.; Creczynski-Pasa, T.B.; Barardi, C.R.M.; O Simões, C.M. Cytotoxicity and potential antiviral evaluation of violacein produced by Chromobacterium violaceum. Memórias Inst. Oswaldo Cruz 2003, 98, 843–848. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, L.; Xue, Y.; Zhang, C.; Xing, X.-H.; Lou, K.; Zhang, Z.; Li, Y.; Zhang, G.; Bi, J. Production of violet pigment by a newly isolated psychrotrophic bacterium from a glacier in Xinjiang, China. Biochem. Eng. J. 2009, 43, 135–141. [Google Scholar] [CrossRef]
- Woodhams, D.C.; LaBumbard, B.C.; Barnhart, K.L.; Becker, M.H.; Bletz, M.C.; Escobar, L.A.; Flechas, S.V.; Forman, M.E.; Iannetta, A.A.; Joyce, M.D.; et al. Prodigiosin, Violacein, and Volatile Organic Compounds Produced by Widespread Cutaneous Bacteria of Amphibians Can Inhibit Two Batrachochytrium Fungal Pathogens. Microb. Ecol. 2017, 75, 1049–1062. [Google Scholar] [CrossRef]
- Rao, M.P.N.; Xiao, M.; Li, W.-J. Fungal and Bacterial Pigments: Secondary Metabolites with Wide Applications. Front. Microbiol. 2017, 8, 1113. [Google Scholar] [CrossRef] [Green Version]
- Dharmaraj, S.; Ashokkumar, B.; Dhevendaran, K. Food-grade pigments from Streptomyces sp. isolated from the marine sponge Callyspongia diffusa. Food Res. Int. 2009, 42, 487–492. [Google Scholar] [CrossRef]
- Venil, C.K.; Dufossé, L.; Renuka Devi, P. Bacterial Pigments: Sustainable Compounds with Market Potential for Pharma and Food Industry. Front. Sustain. Food Syst. 2020, 4, 100. [Google Scholar] [CrossRef]
- Hathaway, J.J.M.; Garcia, M.G.; Balasch, M.M.; Spilde, M.N.; Stone, F.D.; Dapkevicius, M.D.L.N.E.; Northup, D.E. Comparison of Bacterial Diversity in Azorean and Hawai’ian Lava Cave Microbial Mats. Geomicrobiol. J. 2014, 31, 205–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristjánsson, J.K.; Hjörleifsdóttir, S.; Marteinsson, V.T.; Alfredsson, G.A. Thermus scotoductus, sp. nov., a Pigment-Producing Thermophilic Bacterium from Hot Tap Water in Iceland and Including Thermus sp. X-1. Syst. Appl. Microbiol. 1994, 17, 44–50. [Google Scholar] [CrossRef]
- Hermansson, M.; Jones, G.W.; Kjelleberg, S. Frequency of antibiotic and heavy metal resistance, pigmentation, and plasmids in bacteria of the marine airwater interface. Appl. Environ. Microbiol. 1987, 53, 2338–2342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabbagh, P.; Namvar, A.E. The eminence status of bacterial pigments under different aspects. Microbiol. Medica 2017, 32, 122–126. [Google Scholar] [CrossRef]
- Zhao, J.; Li, Q.; Sun, T.; Zhu, X.; Xu, H.; Tang, J.; Zhang, X.; Ma, Y. Engineering central metabolic modules of Escherichia coli for improving β-carotene production. Metab. Eng. 2013, 17, 42–50. [Google Scholar] [CrossRef]
- El-Naggar, N.E.-A.; El-Ewasy, S.M. Bioproduction, characterization, anticancer and antioxidant activities of extracellular melanin pigment produced by newly isolated microbial cell factories Streptomyces glaucescens NEAE-H. Sci. Rep. 2017, 7, srep42129. [Google Scholar] [CrossRef]
- Dorina, S.; Judith, S.; Bjoern, W.; Julia, S.; Andrea, S.; Sarah, D.N.; Roland, U. A new strategy for combined isolation of EPS and pigments from cyanobacteria. J. Appl. Phycol. 2020, 32, 1729–1740. [Google Scholar] [CrossRef]
- Shetty, M.R.D.R. Screening and Extraction of Microbial Pigment from Organism Isolated from Marine Water. Int. J. Sci. Res. (IJSR) 2018, 7, 60–66. [Google Scholar] [CrossRef]
- Yang, J.; Guo, L. Biosynthesis of β-carotene in engineered E. coli using the MEP and MVA pathways. Microb. Cell Factories 2014, 13, 160. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Park, S.; Yang, Y.-H.; Choi, K.-Y. Microbial synthesis of violacein pigment and its potential applications. Crit. Rev. Biotechnol. 2021, 41, 879–901. [Google Scholar] [CrossRef] [PubMed]
- Di, L.I.; Yang, L.I.; Jiao-Yang, X.U.; Qing-Yan, L.I.; Jin-Lei, T.A.N.G.; Shi-Ru, J.I.A.; Chang-Hao, B.I.; Zhu-Bo, D.A.I.; Xin-Na, Z.H.U.; Zhang, X.L. Engineering CrtW and CrtZ for improving biosynthesis of astaxanthin in Escherichia coli. Chin. J. Nat. Med. 2020, 18, 666–676. [Google Scholar]
- Nishizaki, T.; Tsuge, K.; Itaya, M.; Doi, N.; Yanagawa, H. Metabolic engineering of carotenoid biosynthesis in Escherichia coli by ordered gene assembly in Bacillus subtilis. Appl. Environ. Microbiol. 2007, 73, 1355–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelz, A.; Wieland, K.P.; Putzbach, K.; Hentschel, P.; Albert, K.; Götz, F. Structure and Biosynthesis of Staphyloxanthin from Staphylococcus aureus. J. Biol. Chem. 2005, 280, 32493–32498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannibal, L.; Lorquin, J.; D’Ortoli, N.A.; Garcia, N.; Chaintreuil, C.; Masson-Boivin, C.; Dreyfus, B.; Giraud, E. Isolation and Characterization of Canthaxanthin Biosynthesis Genes from the Photosynthetic Bacterium Bradyrhizobium sp. Strain ORS278. J. Bacteriol. 2000, 182, 3850–3853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blankenfeldt, W.; Parsons, J.F. The structural biology of phenazine biosynthesis. Curr. Opin. Struct. Biol. 2014, 29, 26–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henke, N.A.; Wiebe, D.; Pérez-García, F.; Peters-Wendisch, P.; Wendisch, V.F. Co-production of cell-bound and secreted value-added compounds: Simultaneous production of carotenoids and amino acids by Corynebacterium glutamicum. Bioresour. Technol. 2018, 247, 744–752. [Google Scholar] [CrossRef]
- Li, X.-R.; Tian, G.-Q.; Shen, H.-J.; Liu, J.-Z. Metabolic engineering of Escherichia coli to produce zeaxanthin. J. Ind. Microbiol. Biotechnol. 2015, 42, 627–636. [Google Scholar] [CrossRef]
- Su, A.; Chi, S.; Li, Y.; Tan, S.; Qiang, S.; Chen, Z.; Meng, Y. Metabolic redesign of Rhodobacter sphaeroides for lycopene production. J. Agric. Food Chem. 2018, 66, 5879–5885. [Google Scholar] [CrossRef]
- Taniguchi, H.; Henke, N.A.; Heider, S.A.; Wendisch, V.F. Overexpression of the primary sigma factor gene sigA improved carotenoid production by Corynebacterium glutamicum: Application to production of β-carotene and the non-native linear C50 carotenoid bisanhydrobacterioruberin. Metab. Eng. Commun. 2017, 4, 1–11. [Google Scholar] [CrossRef]
- della-Cioppa, G.; Garger, S.J.; Sverlow, G.G.; Turpen, T.H.; Grill, L.K. Melanin production in Escherichia coli from a cloned tyrosinase gene. Biotechnology 1990, 8, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-H.; Kim, J.-E.; Lee, S.-H.; Park, H.-M.; Choi, M.-S.; Kim, J.-Y.; Lee, S.-H.; Shin, Y.-C.; Keasling, J.; Kim, S.-W. Engineering the lycopene synthetic pathway in E. coli by comparison of the carotenoid genes of Pantoea agglomerans and Pantoea ananatis. Appl. Microbiol. Biotechnol. 2007, 74, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Ho, C.T. Chemopreventive effects of natural dietary compounds on cancer development. Chem. Soc. Rev. 2008, 37, 2558–2574. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. CA A Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagchi, D.; Swaroop, A.; Preuss, H.G.; Bagchi, M. Free radical scavenging, antioxidant and cancer chemoprevention by grape seed proanthocyanidin: An overview. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2014, 768, 69–73. [Google Scholar] [CrossRef]
- Fernandes, C.J.; Doddavarapu, B.; Harry, A.; Dilip, S.P.S.; Ravi, L. Isolation and Identification of Pigment Producing Actinomycete Saccharomonospora azurea SJCJABS01. Biomed. Pharmacol. J. 2021, 14, 2261–2269. [Google Scholar] [CrossRef]
- Abraham, J.; Chauhan, R. Profiling of red pigment produced by Streptomyces sp. JAR6 and its bioactivity. 3 Biotech 2018, 8, 22. [Google Scholar] [CrossRef]
- Selvameenal, L.; Radhakrishnan, M.; Balagurunathan, R. Antibiotic pigment from desert soil actinomycetes; biological activity, purification and chemical screening. Indian J. Pharm. Sci. 2009, 71, 499. [Google Scholar] [CrossRef] [Green Version]
- Shaaban, M.T.; El-Sabbagh, S.M.M.; Alam, A. Studies on an actinomycete producing a melanin pigment inhibiting aflatoxin B1 production by Aspergillus flavus. Life Sci. J. 2013, 10, 1437–1448. [Google Scholar]
- Chang, Y.; Xing, L.; Sun, C.; Liang, S.; Liu, T.; Zhang, X.; Zhu, T.; Pfeifer, B.A.; Che, Q.; Zhang, G.; et al. Monacycliones G–K and ent-Gephyromycin A, Angucycline Derivatives from the Marine-Derived Streptomyces sp. HDN15129. J. Nat. Prod. 2020, 83, 2749–2755. [Google Scholar] [CrossRef]
- Ramirez-Rodriguez, L.; Stepanian-Martinez, B.; Morales-Gonzalez, M.; Diaz, L. Optimization of the Cytotoxic Activity of Three Streptomyces Strains Isolated from Guaviare River Sediments (Colombia, South America). BioMed Res. Int. 2018, 2018, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prashanthi, K.; Suryan, S.; Varalakshmi, K.N. In vitro anticancer property of yellow pigment from Streptomyces griseoaurantiacus JUACT 01. Braz. Arch. Biol. Technol. 2015, 58, 869–876. [Google Scholar] [CrossRef]
- Hegazy, G.E.; Abu-Serie, M.M.; Abo-Elela, G.M.; Ghozlan, H.; Sabry, S.A.; Soliman, N.A.; Abdel-Fattah, Y.R. In vitro dual (anticancer and antiviral) activity of the carotenoids produced by haloalkaliphilic archaeon Natrialba sp. M6. Sci. Rep. 2020, 10, 5986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moayedi, A.; Nowroozi, J.; Sepahi, A.A. Cytotoxic effect of pyocyanin on human pancreatic cancer cell line (Panc-1). Iran J. Basic Med. Sci. 2018, 21, 794–799. [Google Scholar]
- Vipin, C.; Ashwini, P.; Kavya, A.; Rekha, P. Overproduction of Pyocyanin in Pseudomonas aeruginosa by Supplementation of Pathway Precursor Shikimic acid and Evaluation of its Activity. Res. J. Pharm. Technol. 2017, 10, 533–536. [Google Scholar] [CrossRef]
- Stankovic, N.; Radulovic, V.; Petkovic, M.; Vuckovic, I.; Jadranin, M.; Vasiljevic, B.; Nikodinovic-Runic, J. Streptomyces sp. JS520 produces exceptionally high quantities of undecylprodigiosin with antibacterial, antioxidative, and UV-protective properties. Appl. Microbiol. Biotechnol. 2012, 96, 1217–1231. [Google Scholar] [CrossRef]
- Charkoudian, L.K.; Fitzgerald, J.T.; Khosla, C.; Champlin, A. In Living Color: Bacterial Pigments as an Untapped Resource in the Classroom and Beyond. PLoS Biol. 2010, 8, e1000510. [Google Scholar] [CrossRef] [Green Version]
- Teruel, M.A.; Gontier, E.; Bienaime, C.; Saucedo, J.N.; Barbotin, J.N. Response surface analysis of chlortetracycline and tetracycline production with K-carraimmobilizedbilized Streptomyces aureofaciens. Enzym. Microb. Technol. 1997, 21, 314–320. [Google Scholar] [CrossRef]
- Rutherford, S.T.; Bassler, B.L. Bacterial Quorum Sensing: Its Role in Virulence and Possibilities for Its Control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef]
- Naclerio, G.; Ricca, E.; Sacco, M.; De Felice, M. Antimicrobial activity of a newly identified bacteriocin of Bacillus cereus. Appl. Environ. Microbiol. 1993, 59, 4313–4316. [Google Scholar] [CrossRef] [Green Version]
- Baron, S.S.; Terranova, G.; Rowe, J.J. Molecular mechanism of the antimicrobial action of pyocyanin. Curr. Microbiol. 1989, 18, 223–230. [Google Scholar] [CrossRef]
- Raji El Feghali, P.A.; Nawas, T. Pyocyanin: A powerful inhibitor of bacterial growth and biofilm formation. Madridge J. Case Rep. Stud. 2018, 2, 101–107. [Google Scholar] [CrossRef]
- Akaike, T. Role of free radicals in viral pathogenesis and mutation. Rev. Med. Virol. 2001, 11, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Brunelle, M.-N.; Jacquard, A.-C.; Pichoud, C.; Durantel, D.; Carrouée-Durantel, S.; Villeneuve, J.-P.; Trépo, C.; Zoulim, F. Susceptibility to antivirals of a human HBV strain with mutations conferring resistance to both lamivudine and adefovir. Hepatology 2005, 41, 1391–1398. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Zeng, C.; Liu, R.; Chen, J.; Li, R.; Wang, X.; Bai, W.; Liu, X.; Xiang, T.; Zhang, L.; et al. Antiviral activity and specific modes of action of bacterial prodigiosin against Bombyx mori nucleopolyhedrovirus in vitro. Appl. Microbiol. Biotechnol. 2015, 100, 3979–3988. [Google Scholar] [CrossRef]
- Suryawanshi, R.K.; Koujah, L.; Patil, C.D.; Ames, J.M.; Agelidis, A.; Yadavalli, T.; Patil, S.V.; Shukla, D. Bacterial Pigment Prodigiosin Demonstrates a Unique Antiherpesvirus Activity That Is Mediated through Inhibition of Prosurvival Signal Transducers. J. Virol. 2020, 94, e00251-20. [Google Scholar] [CrossRef] [PubMed]
- Durán, M.; Faljoni-Alario, A.; Durán, N. Chromobacterium violaceum and its important metabolites—Review. Folia Microbiol. 2010, 55, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Lionakis, M.S. Genetic Susceptibility to Fungal Infections in Humans. Curr. Fungal Infect. Rep. 2012, 6, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Thompson, G.R.; Cadena, J.; Patterson, T.F. Overview of Antifungal Agents. Clin. Chest Med. 2009, 30, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Jimtha, C.J.; Jishma, P.; Sreelekha, S.; Chithra, S.; Radhakrishnan, E. Antifungal properties of prodigiosin producing rhizospheric Serratia sp. Rhizosphere 2017, 3, 105–108. [Google Scholar] [CrossRef]
- Dawoud, T.M.; Alharbi, N.S.; Theruvinthalakal, A.M.; Thekkangil, A.; Kadaikunnan, S.; Khaled, J.M.; Almanaa, T.N.; Sankar, K.; Innasimuthu, G.M.; Alanzi, K.F.; et al. Characterization and antifungal activity of the yellow pigment produced by a Bacillus sp. DBS4 isolated from the lichen Dirinaria agealita. Saudi J. Biol. Sci. 2019, 27, 1403–1411. [Google Scholar] [CrossRef] [PubMed]
- Wagh, P.; Mane, R. Identification and characterization of extracellular red pigment producing Neisseria spp. isolated from soil sample. Int. J. Innov. Knowl. Concept 2017, 5, 23–25. [Google Scholar]
- Houshaymi, B.; Awada, R.; Kedees, M.; Soayfane, Z. Pyocyanin, a Metabolite of Pseudomonas Aeruginosa, Exhibits Antifungal Drug Activity Through Inhibition of a Pleiotropic Drug Resistance Subfamily FgABC3. Drug Res. 2019, 69, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Sudhakar, T.; Karpagam, S. Antifungal efficacy of pyocyanin produced from bioindicators of nosocomial hazards. In Proceedings of the International Conference on Green technology and environmental Conservation (GTEC-2011), Chennai, India, 15–17 December 2011; pp. 224–229. [Google Scholar]
- Kerr, J.R.; Taylor, G.W.; Rutman, A.; Hoiby, N.; Cole, P.J.; Wilson, R. Pseudomonas aeruginosa pyocyanin and 1-hydroxyphenazine inhibit fungal growth. J. Clin. Pathol. 1999, 52, 385–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoud, S.Y.; Ziedan, E.S.H.; Farrag, E.S.; Kalafalla, R.S.; Ali, A.M. Antifungal activity of pyocyanin produced by Pseudomonas aeruginosa against Fusarium oxysporum Schlech phytopathogenic fungi. Int. J. PharmTech Res. 2016, 9, 43–50. [Google Scholar]
- Bisht, G.; Srivastava, S.; Kulshreshtha, R.; Sourirajan, A.; Baumler, D.J.; Dev, K. Applications of red pigments from psychrophilic Rhodonellum psychrophilum GL8 in health, food and antimicrobial finishes on textiles. Process. Biochem. 2020, 94, 15–29. [Google Scholar] [CrossRef]
- Mitra, A.K.; Mawson, A.R. Neglected Tropical Diseases: Epidemiology and Global Burden. Trop. Med. Infect. Dis. 2020, 2, 36. [Google Scholar] [CrossRef] [Green Version]
- Silva, T.R.; Duarte, A.W.F.; Passarini, M.R.Z.; Ruiz, A.L.T.G.; Franco, C.H.; Moraes, C.B.; de Melo, I.S.; Rodrigues, R.A.; Fantinatti-Garboggini, F.; Oliveira, V.M. Bacteria from Antarctic environments: Diversity and detection of antimicrobial, antiproliferative, and antiparasitic activities. Polar Biol. 2018, 41, 1505–1519. [Google Scholar] [CrossRef]
- Bilsland, E.; Tavella, T.A.; Krogh, R.; Stokes, J.E.; Roberts, A.; Ajioka, J.; Spring, D.R.; Andricopulo, A.D.; Costa, F.T.M.; Oliver, S.G. Antiplasmodial and trypanocidal activity of violacein and deoxyviolacein produced from synthetic operons. BMC Biotechnol. 2018, 18, 22. [Google Scholar] [CrossRef] [Green Version]
- Rahul, S.; Chandrashekhar, P.; Hemant, B. Natural Product Research: Formerly Natural Product Letters Nematicidal activity of microbial pigment from Serratia marcescens. Nat. Prod. Res. 2014, 28, 1399–1404. [Google Scholar] [CrossRef]
- Gutierrez-dalmau, A.; Campistol, J.M. Immunosuppressive Therapy and Malignancy in Organ A Systematic Review. Drugs 2007, 67, 1167–1198. [Google Scholar] [CrossRef] [PubMed]
- Kawauchiab, K.; Shibutaniab, K.; Yagisawaa, H.; Kamataa, H.; Nakatsuji, S.; Anzaic, H.; Yokoyamab, Y.; Ikegamib, Y.; Moriyamad, Y.; Hirata, H. A Possible Immunosuppressant, Cycloprodigiosin Hydrochloride, Obtained from Pseudoalteromonas denitrificans. Biochem. Biophys. Res. Commun. 1997, 547, 543–547. [Google Scholar] [CrossRef]
- Nakashima, T.; Kurachi, M.; Kato, Y.; Yamaguchi, K.; Oda, T. Characterization of Bacterium Isolated from the Sediment at Coastal Area of Omura Bay in Japan and Several Biological Activities of Pigment Produced by This Isolate. Microbiol. Immunol. 2005, 49, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.; Chander, R.; Sainis, K. A novel prodigiosin-like immunosuppressant from an alkalophilic Micrococcus sp. Int. Immunopharmacol. 2003, 3, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Burrows, J.A. Palette of Our Palates: A Brief History of Food Coloring and Its Regulation. Compr. Rev. Food Sci. Food Saf. 2009, 8, 394–408. [Google Scholar] [CrossRef]
- Downham, A.; Collins, P. Colouring our foods in the last and next millennium. Int. J. Food Sci. Technol. 2000, 35, 5–22. [Google Scholar] [CrossRef]
- Manikprabhu, D.; Lingappa, K. γ Actinorhodin a natural and attorney source for synthetic dye to detect acid production of fungi. Saudi J. Biol. Sci. 2013, 20, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Galaffu, N.; Bortlik, K.; Michel, M. An industry perspective on natural food colour stability. In Colour Additives for Foods and Beverages; Elsevier Ltd.: Amsterdam, The Netherlands, 2015. [Google Scholar] [CrossRef]
- Wrolstad, R.E.; Culver, C.A. Alternatives to Those Artificial FD&C Food Colorants. Annu. Rev. Food Sci. Technol. 2012, 3, 59–77. [Google Scholar] [CrossRef]
- Özkan, G.; Bilek, S.E. Microencapsulation of Natural Food Colourants. Int. J. Nutr. Food Sci. 2014, 3, 145–156. [Google Scholar] [CrossRef]
- Silva, P.I.; Stringheta, P.C.; Teófilo, R.F.; de Oliveira, I.R.N. Parameter optimization for spray-drying microencapsulation of jaboticaba (Myrciaria jaboticaba) peel extracts using simultaneous analysis of responses. J. Food Eng. 2013, 117, 538–544. [Google Scholar] [CrossRef] [Green Version]
- Rocha, G.A.; Fávaro-Trindade, C.S.; Grosso, C.R.F. Microencapsulation of lycopene by spray drying: Characterization, stability and application of microcapsules. Food Bioprod. Process. 2012, 90, 37–42. [Google Scholar] [CrossRef]
- Heer, K.; Sharma, S. Microbial pigments as a natural colour—A review. Int. J. Pharm. Sci. Res. 2017, 8, 1913–1922. [Google Scholar] [CrossRef]
- Yokoyama, A. Composition and presumed biosynthetic pathway of carotenoids in the astaxanthin-producing bacterium. FEMS Microbiol. Lett. 1995, 128, 139–144. [Google Scholar] [CrossRef]
- Surai, P.F. The antioxidant properties of canthaxanthin and its potential effects in the poultry eggs and on embryonic development of the chick. Part 2. World’s Poult. Sci. J. 2012, 68, 717–726. [Google Scholar] [CrossRef]
- Wagner-Döbler, I.; Beil, W.; Lang, S.; Meiners, M.; Laatsch, H. Integrated approach to explore the potential of marine microorganisms for the production of bioactive metabolites. In Tools and Applications of Biochemical Engineering Science; Springer: Berlin/Heidelberg, Germany, 2002; pp. 207–238. [Google Scholar] [CrossRef]
- Guillerme, J.B.; Couteau, C.; Coiffard, L. Applications for marine resources in cosmetics. Cosmetics 2017, 4, 35. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Hua, Y. Carotenoid biosynthesis in extremophilic Deinococcus-Thermus bacteria. Trends Microbiol. 2010, 18, 512–520. [Google Scholar] [CrossRef]
- Wan, M.; Hou, D.; Li, Y.; Fan, J.; Huang, J.; Liang, S.; Li, S. The effective photoinduction of Haematococcus pluvialis for accumulating astaxanthin with attached cultivation. Bioresour. Technol. 2014, 163, 26–32. [Google Scholar] [CrossRef]
- Shindo, K.; Misawa, N. New and rare carotenoids isolated from marine bacteria and their antioxidant activities. Mar. Drugs 2014, 12, 1690–1698. [Google Scholar] [CrossRef] [Green Version]
- Rittié, L.; Fisher, G.J. UV-light-induced signal cascades and skin aging. Ageing Res. Rev. 2002, 1, 705–720. [Google Scholar] [CrossRef]
- Sorrels, C.M.; Proteau, P.J.; Gerwick, W.H. Organization, evolution, and expression analysis of the biosynthetic gene cluster for scytonemin, a cyanobacterial UV-absorbing pigment. Appl. Environ. Microbiol. 2009, 75, 4861–4869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastogi, R.P.; Sinha, R.P. Incharoensakdi, Acharacterizationterization, UV-induction and photoprotective function of sunscreen pigment, scytonemin from Rivularia sp. HKAR-4. Chemosphere 2013, 93, 1874–1878. [Google Scholar] [CrossRef] [PubMed]
- Meot-Duros, L.; Cérantola, S.; Talarmin, H.; Le Meur, C.; LE Floch, G.; Magné, C. New antibacterial and cytotoxic activities of falcarindiol isolated in Crithmum maritimum L. leaf extract. Food Chem. Toxicol. 2010, 48, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Alihosseini, F.; Ju, K.S.; Lango, J.; Hammock, B.D.; Sun, G. Antibacterial colorants: Characterization of prodiginines and their applications on textile materials. Biotechnol. Prog. 2008, 24, 742–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venil, C.K.; Dufossé, L.; Velmurugan, P.; Malathi, M.; Lakshmanaperumalsamy, P. Extraction and application of pigment from Serratia marcescens SB08, an insect enteric gut bacterium, for textile dyeing. Textiles 2021, 1, 21–36. [Google Scholar] [CrossRef]
- Sajjad, W.; Ahmad, S.; Aziz, I.; Azam, S.S.; Hasan, F.; Shah, A.A. Antiproliferative, antioxidant and binding mechanism analysis of prodigiosin from newly isolated radio-resistant Streptomyces sp. Strain WMA-LM31. Mol. Biol. Rep. 2018, 45, 1787–1798. [Google Scholar] [CrossRef]
- Montaner, B.; Prez-Toms, R. The prodigiosins: A new family of anticancer drugs. Curr. Cancer Drug Targets 2003, 3, 57–65. [Google Scholar] [CrossRef]
- Cao, W.; Chen, W.; Sun, S.; Guo, P.; Song, J.; Tian, C. Investigating the antioxidant mechanism of violacein by density functional theory method. J. Mol. Struct. Theochem. 2007, 817, 1–4. [Google Scholar] [CrossRef]
- Suryawanshi, R.K.; Patil, C.D.; Borase, H.P.; Narkhede, C.P.; Stevenson, A.; Hallsworth, J.E.; Patil, S.V. Towards an understanding of bacterial metabolites prodigiosin and violacein and their potential for use in commercial sunscreens. Int. J. Cosmet. Sci. 2015, 37, 98–107. [Google Scholar] [CrossRef]
- Britton, G. Structure and properties of carotenoids in relation to function. FASEB J. 1995, 9, 1551–1558. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| S. No. | Pigment | Colour | Microorganisms | Clinical Uses | References |
|---|---|---|---|---|---|
| 1. Carotenoids | |||||
| (i) | Canthaxanthin | Orange to deep pink | Bradyrhizobium spp., Lactobacillus pluvialis | Antioxidant, Photoprotectant, Anti-cancer, Anti-inflammatory | [17] |
| (ii) | Astaxanthin | Red to orange | Agrobacterium aurantiacum, Paracoccus carotinifaciens | Antioxidant, Photo-protectant, Anti-cancer, Anti-inflammatory | [18] |
| (iii) | Zeaxanthin | Yellow | Staphylococcus aureus, Flavobacterium spp., Paracoccus zeaxanthinifaciens | Photoprotectant, Antioxidant | [19] |
| (iv) | β-Carotene | Red | Rhodococcus maris, Rhodococcus ruber | Anticancer, Antioxidant | [20] |
| (v) | Staphyloxanthin | Yellow | Staphylococcus aureus | Antioxidant | [21] |
| (vi) | Deinoxanthin | Red | Deinococcus radiodurans | Anti-cancer | [22] |
| 2. Prodiginines | |||||
| (i) | Prodigiosin | Red | Serratia marcescens | Anti-cancer, DNA cleavage, Immunosuppressant | [23] |
| (ii) | Undecylprodigiosin | Red | Streptomyces sp. | Antimalarial activity, Antibacterial, Antioxidant, Anti-cancer | [24,25] |
| (iii) | Heptyl Prodigiosin | Red | α-Proteobacteria | Antiplasmodial | [26] |
| 3. Phycobiliproteins | |||||
| (i) | Pycocyanin | Blue | Pseudomonas spp. | Cytotoxicity, Neutrophil apoptosis, Ciliary dysmotility, Proinflammatory | [27] |
| 4.Oxyindoles | |||||
| (i) | Violacein | Purple | Chromobacterium violaceum, Collimonas sp., Duganella sp., Pseudoalteromonas sp. | Antifungal, Antibacterial, Antiplasmodial, Anti-cancer | [28,29,30,31] |
| S. No. | Pigments | Genes Involved in the Biosynthesis of Pigments | References |
|---|---|---|---|
| 1. | β-carotene | crtE, crtY, crtI, crtB | [43] |
| 2. | Violacein | vioA, vioB, vioC, vioD, vioE | [44] |
| 3. | Prodigiosin | pig B, pigC, pigD, pigE, pigF, pigM, pigH, pigJ | [23] |
| 4. | Astaxanthin | crtW, crtZ | [45] |
| 5. | Zeaxanthin | crtE, crtB, crtI, crtY, crtZ | [46] |
| 6. | Staphyloxanthin | crtO, crtP, crtQ, crtM, crtN | [47] |
| 7. | Canthaxanthin | crtE, crtY, crtI, crtB, crtW | [48] |
| 8. | Pyocyanine | phzE, phzD, phzF, phzB, phzG | [49] |
| S. No. | Microorganism | Type of Modification | Gene/Pathway Modified | Genes Origin | Pigment Produced | References |
|---|---|---|---|---|---|---|
| 1. | C. glutamicum | Deletion | Sigma B factor (sigB) | - | Increase in Decaprenoxanthin β-carotene | [53] |
| Over-expression | Sigma A factor (sigA) | - | Increase in Decaprenoxanthin Lycopene | |||
| 2. | E. coli | Pathway modification | MEP pathway | - | Lycopene | [43] |
| Insertion of pathway | MVA pathway | - | ||||
| Insertion | mel and ORF438 gene | Streptomyces antibioticus | Eumelanin | [54] | ||
| crtE, crtB, and crtI | Pantoea agglomerans | Lycopene | ||||
| Introduction of intergenic regions | crtY and crtZ genes | Pantoea ananatis | Zeaxanthin | [55] | ||
| Deletion | zwf gene | - | Increase in β-carotene | [51] | ||
| ptsHIcrr operon | - | |||||
| 3. | Rhodobacter sphaeroides | Replace | crtI3 with crtI4 | Rhodospirillum rubrum | Lycopene | [52] |
| Deletion | crtC gene | |||||
| Knock out | zwf gene |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agarwal, H.; Bajpai, S.; Mishra, A.; Kohli, I.; Varma, A.; Fouillaud, M.; Dufossé, L.; Joshi, N.C. Bacterial Pigments and Their Multifaceted Roles in Contemporary Biotechnology and Pharmacological Applications. Microorganisms 2023, 11, 614. https://doi.org/10.3390/microorganisms11030614
Agarwal H, Bajpai S, Mishra A, Kohli I, Varma A, Fouillaud M, Dufossé L, Joshi NC. Bacterial Pigments and Their Multifaceted Roles in Contemporary Biotechnology and Pharmacological Applications. Microorganisms. 2023; 11(3):614. https://doi.org/10.3390/microorganisms11030614
Chicago/Turabian StyleAgarwal, Himani, Sneh Bajpai, Arti Mishra, Isha Kohli, Ajit Varma, Mireille Fouillaud, Laurent Dufossé, and Naveen Chandra Joshi. 2023. "Bacterial Pigments and Their Multifaceted Roles in Contemporary Biotechnology and Pharmacological Applications" Microorganisms 11, no. 3: 614. https://doi.org/10.3390/microorganisms11030614