
Genomic Analysis of Haloarchaea from Diverse Environments, including Permian Halite, Reveals Diversity of Ultraviolet Radiation Survival and DNA Photolyase Gene Variants

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
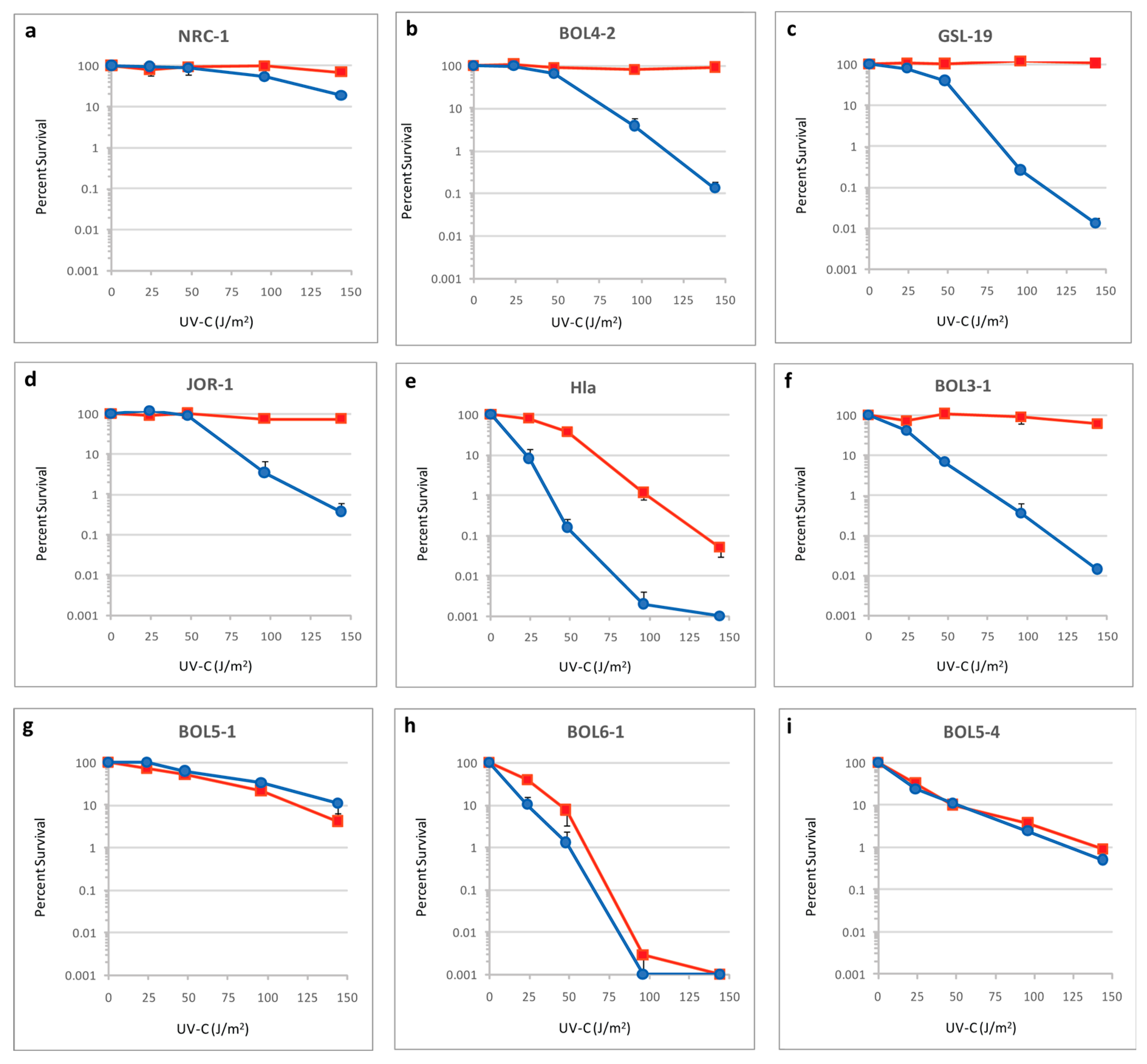
2.2. UV Survival Assays
2.3. Phylogenetic Analysis
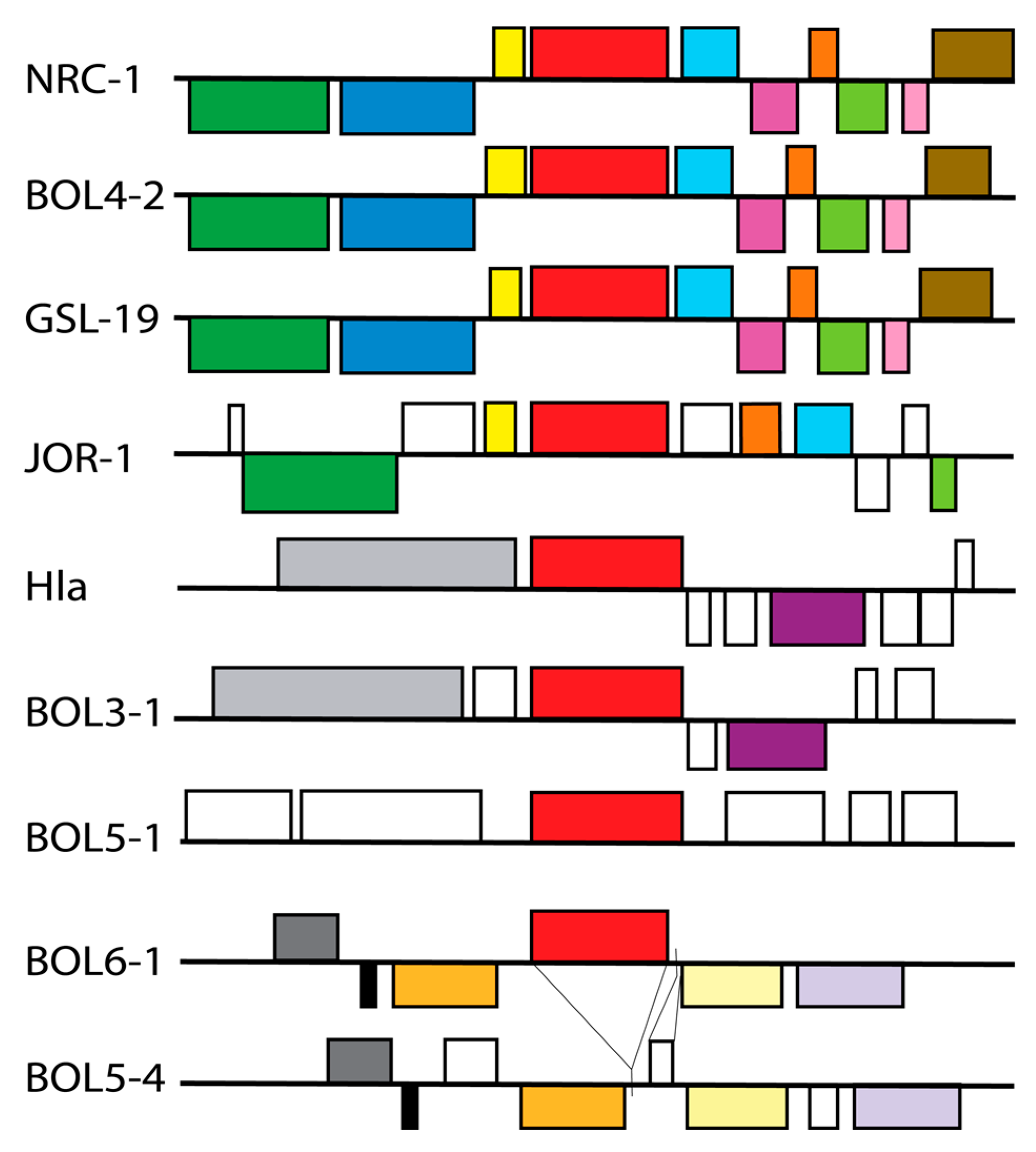
2.4. Gene Organization
2.5. Protein Structural Analysis
3. Results and Discussion
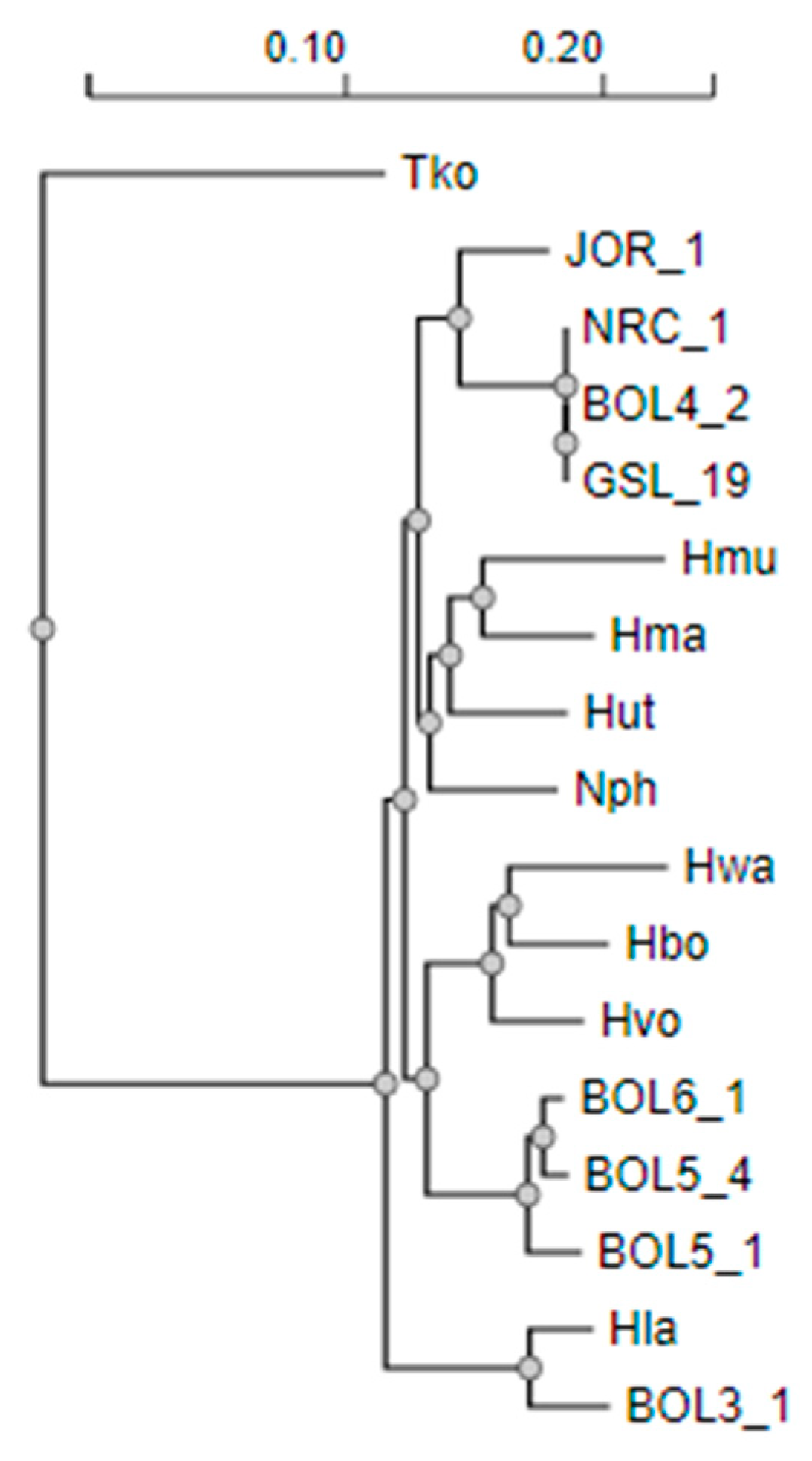
3.1. Phylogeny of Diverse Haloarchaea
3.2. Genomic Characteristics of Diverse Haloarchaea
3.3. Photolyase Genes and Gene Organization
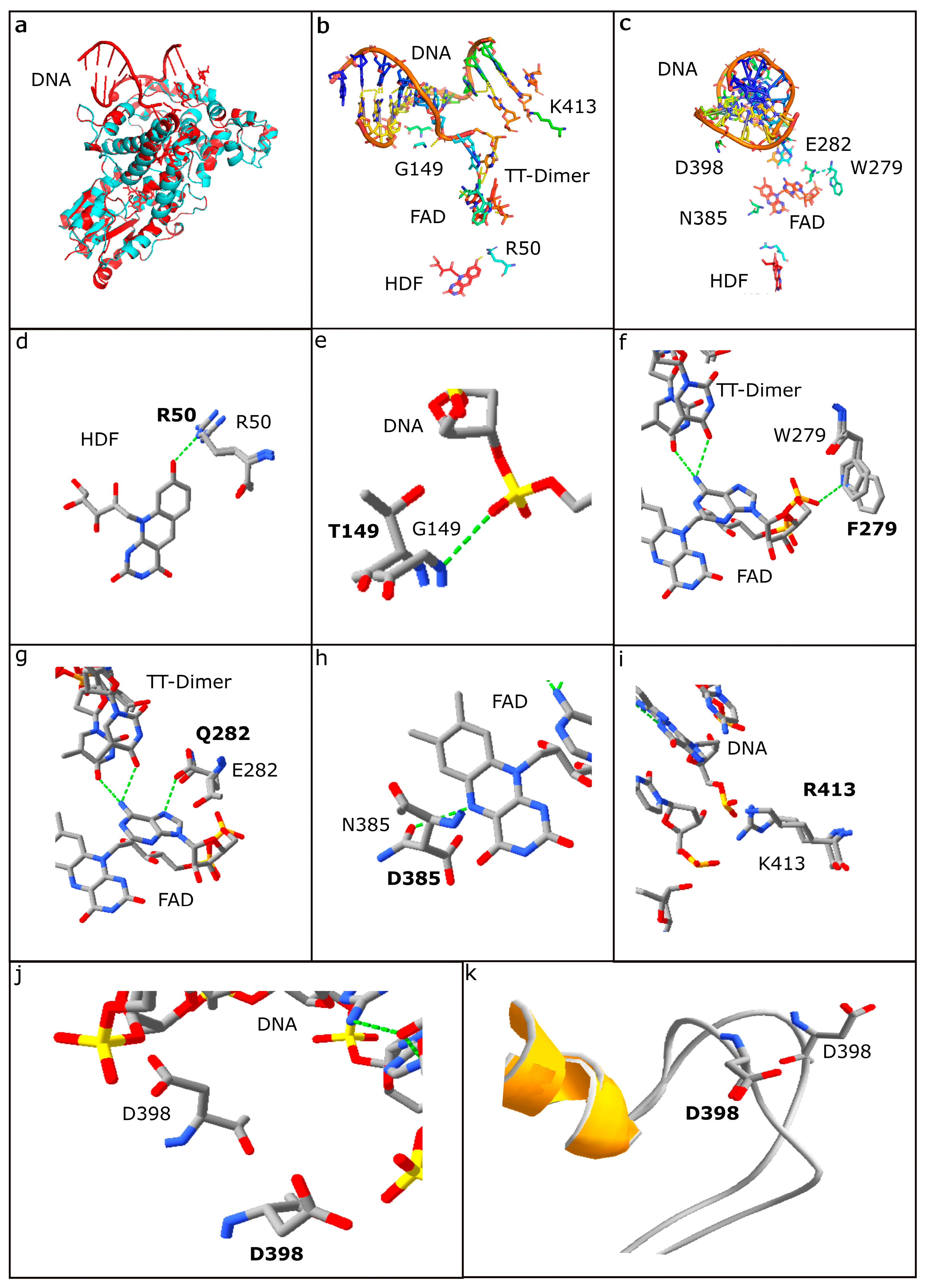
3.4. Structural Analysis of Photolyases
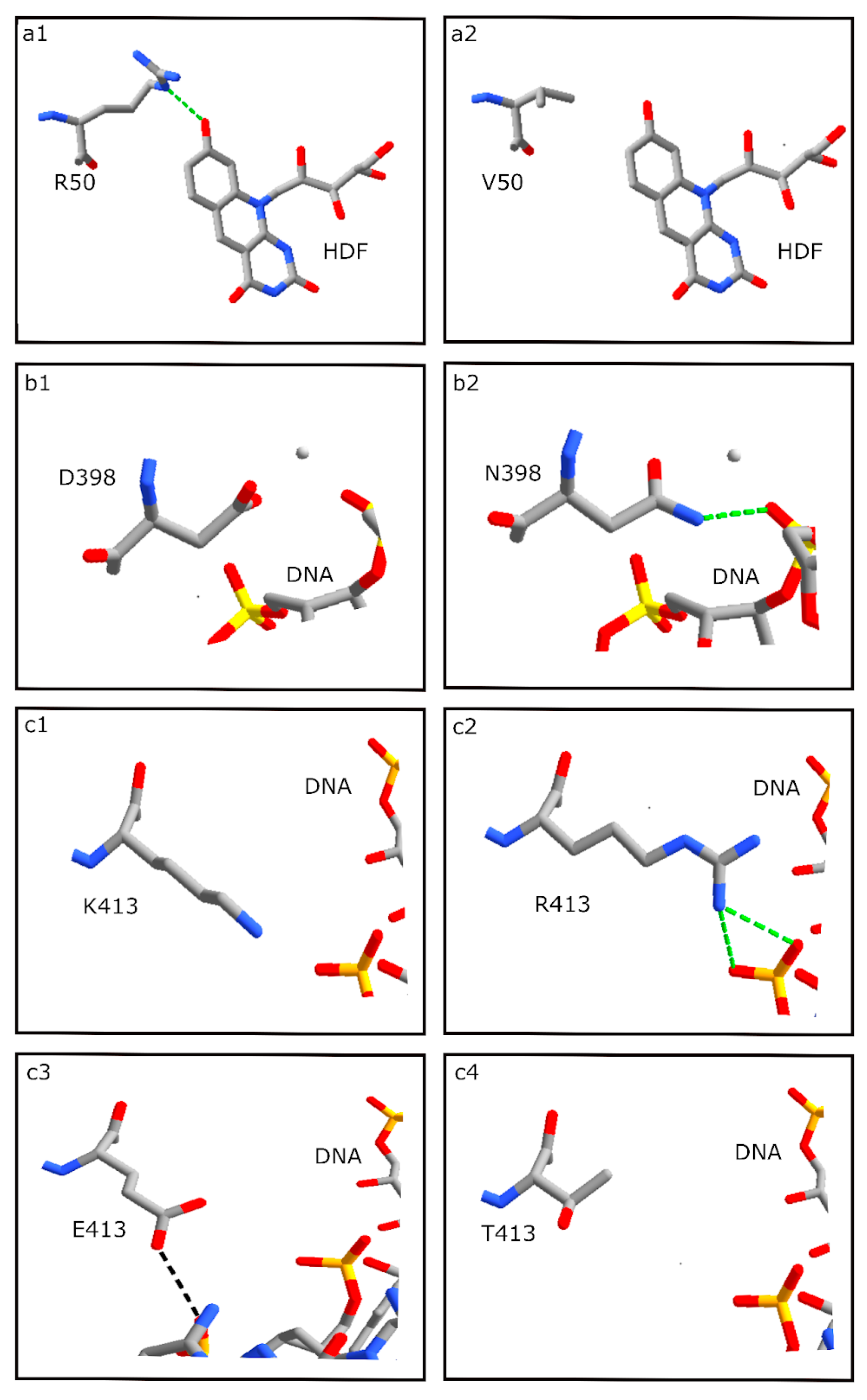
3.5. HDF Binding Residues
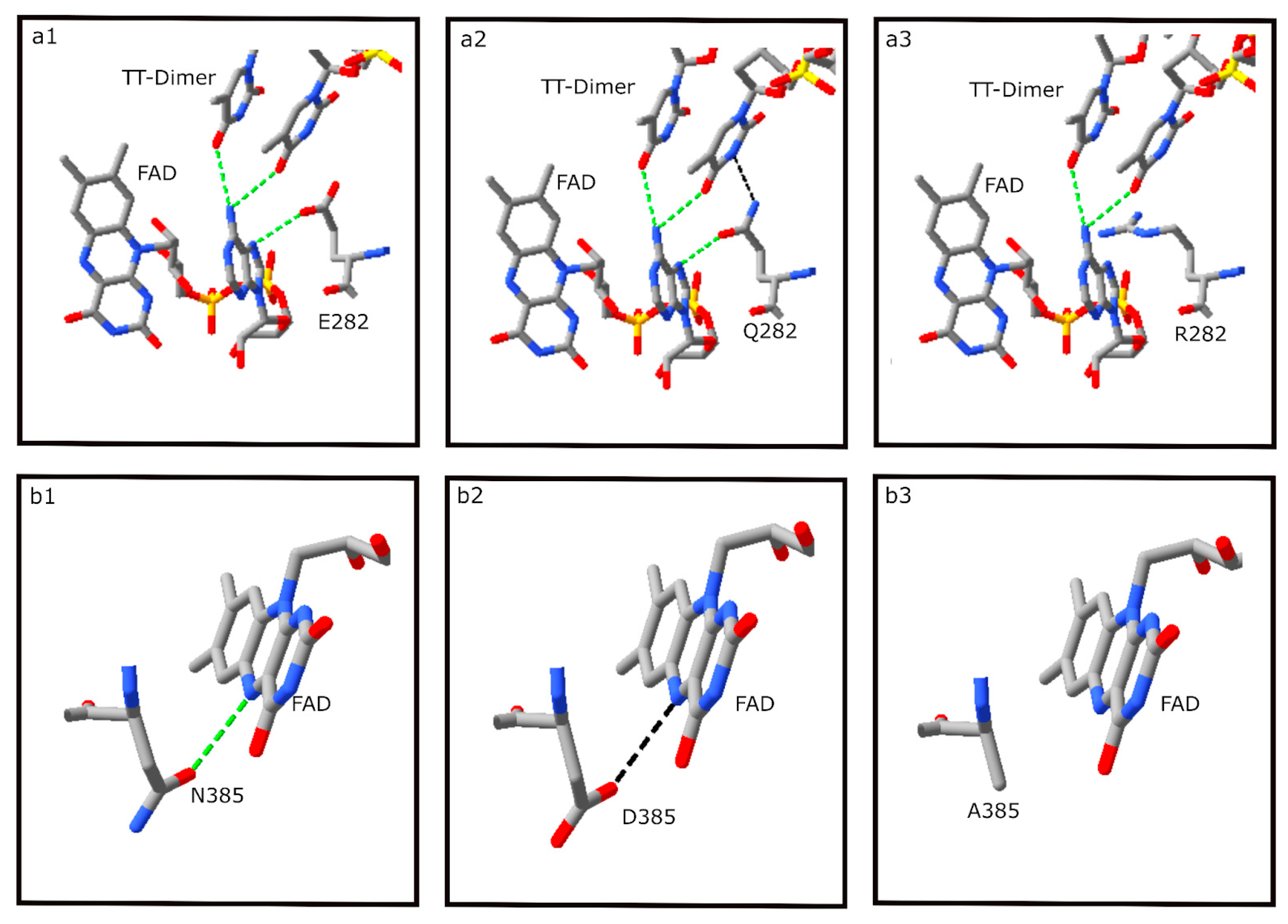
3.6. DNA Binding Residues
3.7. FAD Binding Residues
3.8. Environmental Implications
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- McGenity, T.J.; Gemmell, R.T.; Grant, W.D.; Stan-Lotter, H. Origins of halophilic microorganisms in ancient salt deposits. Environ. Microbiol. 2000, 2, 243–250. [Google Scholar] [CrossRef] [PubMed]
- DasSarma, S. Extreme Halophiles Are Models for Astrobiology. Microbe 2006, 1, 120–126. [Google Scholar] [CrossRef] [Green Version]
- DasSarma, S.; Schwieterman, E. Early evolution of purple retinal pigments on Earth and implications for exoplanet biosignatures. Int. J. Astrobiol. 2018, 20, 241–250. [Google Scholar] [CrossRef] [Green Version]
- DasSarma, S.; DasSarma, P.; Laye, V.J.; Schwieterman, E.W. Extremophilic models for astrobiology: Haloarchaeal survival strategies and pigments for remote sensing. Extremophiles 2020, 24, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Carrier, B.L.; Beaty, D.W.; Meyer, M.A.; Blank, J.G.; Chou, L.; DasSarma, S.; Des Marais, D.J.; Eigenbrode, J.L.; Grefenstette, N.; Lanza, N.L.; et al. Mars Extant Life: What’s Next? Conf. Rep. Astrobiol. 2020, 20, 785–814. [Google Scholar] [CrossRef]
- Mancinelli, R.L.; Fahlen, T.F.; Landheim, R.; Klovstad, M.R. Brines and evaporites: Analogs for Martian life. Adv. Space Res. 2004, 33, 1244–1246. [Google Scholar] [CrossRef]
- DasSarma, P.; Laye, V.J.; Harvey, J.; Reid, C.; Shultz, J.; Yarborough, A.; Lamb, A.; Koske-Phillips, A.; Herbst, A.; Molina, F.; et al. Survival of halophilic Archaea in Earth’s cold stratosphere. Int. J. Astrobiol. 2016, 16, 321–327. [Google Scholar] [CrossRef] [Green Version]
- DasSarma, P.; DasSarma, S. Survival of microbes in Earth’s stratosphere. Curr. Opin. Microbiol. 2018, 43, 24–30. [Google Scholar] [CrossRef] [PubMed]
- DasSarma, P.; Antunes, A.; Simões, M.F.; DasSarma, S. Earth’s Stratosphere and Microbial Life. Curr. Issues Mol. Biol. 2020, 38, 197–244. [Google Scholar] [CrossRef] [Green Version]
- DasSarma, S.; Berquist, B.R.; Coker, J.A.; DasSarma, P.; Müller, J.A. Post-genomics of the model haloarchaeon Halobacterium sp. NRC-1. Saline Syst. 2006, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- DeVeaux, L.C.; Müller, J.A.; Smith, J.; Petrisko, J.; Wells, D.P.; DasSarma, S. Extremely Radiation-Resistant Mutants of a Halophilic Archaeon with Increased Single-Stranded DNA-Binding Protein (RPA) Gene Expression. Radiat. Res. 2007, 168, 507–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCready, S.; Marcello, L. Repair of UV damage in Halobacterium salinarum. Biochem. Soc. Trans. 2003, 31, 694–698. [Google Scholar] [CrossRef] [Green Version]
- McCready, S.; Müller, J.A.; Boubriak, I.; Berquist, B.R.; Ng, W.L.; DasSarma, S. UV irradiation induces homologous recombination genes in the model archaeon, Halobacterium sp. NRC-1. Saline Syst. 2005, 1, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowley, D.J.; Boubriak, I.; Berquist, B.R.; Clark, M.; Richard, E.; Sullivan, L.; DasSarma, S.; McCready, S. The uvrA, uvrB, and uvrC genes are required for repair of ultraviolet light induced DNA photoproducts in Halobacterium sp. NRC-1. Saline Syst. 2006, 2, 11. [Google Scholar] [CrossRef] [Green Version]
- Boubriak, I.; Ng, W.L.; DasSarma, P.; DasSarma, S.; Crowley, D.J.; McCready, S.J. Transcriptional responses to biologically relevant doses of UV-B radiation in the model archaeon, Halobacterium sp. NRC-1. Saline Syst. 2008, 4, 13. [Google Scholar] [CrossRef] [Green Version]
- Capes, M.D.; Coker, J.A.; Gessler, R.; Grinblat-Huse, V.; DasSarma, S.L.; Jacob, C.G.; Kim, J.-M.; DasSarma, P.; DasSarma, S. The information transfer system of halophilic archaea. Plasmid 2011, 65, 77–101. [Google Scholar] [CrossRef] [PubMed]
- Capes, M.D.; DasSarma, P.; DasSarma, S. The core and unique proteins of haloarchaea. BMC Genom. 2012, 13, 39. [Google Scholar] [CrossRef] [Green Version]
- Goo, Y.A.; Roach, J.; Glusman, G.; Baliga, N.S.; Deutsch, K.; Pan, M.; Kennedy, S.; DasSarma, S.; Ng, W.V.; Hood, L. Low-pass sequencing for microbial comparative genomics. BMC Genom. 2004, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Hoff, W.D.; Jung, K.H.; Spudich, J.L. Molecular Mechanisms of Photosignaling by Archaeal Sensory Rhodopsins. Annu. Rev. Biophys. Biomol. Struct. 1997, 26, 223–258. [Google Scholar] [CrossRef] [Green Version]
- DasSarma, P.; DasSarma, S. Gas Vesicle Nanoparticles for Antigen Display. Vaccines 2021, 3, 686–702. [Google Scholar] [CrossRef] [Green Version]
- Krebs, M.P.; Khorana, H.G. Mechanism of light-dependent proton translocation by bacteriorhodopsin. J. Bacteriol. 1993, 175, 1555–1560. [Google Scholar] [CrossRef] [Green Version]
- DasSarma, P.; Zamora, R.C.; Müller, J.A.; DasSarma, S. Genome-wide responses of the model archaeon Halobacterium NRC-1 to oxygen limitation. J. Bacteriol. 2012, 194, 5520–5537. [Google Scholar] [CrossRef] [Green Version]
- Racker, E.; Stoeckenius, W. Reconstitution of purple membrane vesicles catalyzing light-driven proton uptake and adenosine triphosphate formation. J. Biol. Chem. 1974, 249, 662–663. [Google Scholar] [CrossRef] [PubMed]
- Schwieterman, E.W.; Kiang, N.Y.; Parenteau, M.N.; Harman, C.E.; DasSarma, S.; Fisher, T.M.; Arney, G.N.; Hartnett, H.E.; Reinhard, C.T.; Olson, S.L.; et al. Exoplanet Biosignatures: A Review of Remotely Detectable Signs of Life. Astrobiology 2018, 18, 663–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catling, D.C.; Zahnle, K.J. The Archean atmosphere. Sci. Adv. 2020, 6, eaax1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cnossen, I.; Sanz-Forcada, J.; Favata, F.; Witasse, O.; Zegers, T.; Arnold, N.F. Habitat of early life: Solar X-ray and UV radiation at Earth’s surface 4–3.5 billion years ago. J. Geophys. Res. 2007, 112, e02008. [Google Scholar] [CrossRef]
- Ng, W.V.; Kennedy, S.P.; Mahairas, G.G.; Berquist, B.; Pan, M.; Shukla, H.D.; Lasky, S.R.; Baliga, N.S.; Thorsson, V.; Sbrogna, J.; et al. Genome sequence of Halobacterium species NRC-1. Proc. Natl. Acad. Sci. USA 2000, 97, 12176–12181. [Google Scholar] [CrossRef] [Green Version]
- DasSarma, S. Genome Sequence of an Extremely Halophilic Archaeon. In Microbial Genomes; Fraser, C.M., Read, T., Nelson, K.E., Eds.; Humana Press, Inc.: Totowa, NJ, USA, 2004; pp. 383–399. [Google Scholar]
- Gan, R.R.; Yi, E.C.; Chiu, Y.; Lee, H.; Kao, Y.-C.P.; Wu, T.H.; Aebersold, R.; Goodlett, D.R.; Ng, W.V. Proteome Analysis of Halobacterium sp. NRC-1 Facilitated by the Biomodule Analysis Tool. Mol.Cell. Proteom. 2006, 5, 987–997. [Google Scholar] [CrossRef] [Green Version]
- Anderson, I.J.; DasSarma, P.; Lucas, S.; Copeland, A.; Lapidus, A.; Del Rio, T.G.; Tice, H.; Dalin, E.; Bruce, D.C.; Goodwin, L.; et al. Complete genome sequence of the Antarctic Halorubrum lacusprofundi type strain ACAM 34. Stand. Genom. Sci. 2016, 11, 70. [Google Scholar] [CrossRef] [Green Version]
- Mees, A.; Klar, T.; Gnau, P.; Hennecke, U.; Eker, A.P.M.; Carell, T.; Essen, L.-O. Crystal structure of a photolyase bound to a CPD-like DNA lesion after in situ repair. Science 2004, 306, 1789–1793. [Google Scholar] [CrossRef]
- Tamada, T.; Kitadokoro, K.; Higuchi, Y. Crystal structure of DMA photolyase from Anacystis nidulans. Nat. Struct. Mol. Biol. 1997, 4, 887–891. [Google Scholar] [CrossRef] [PubMed]
- DasSarma, P.; DasSarma, S. On the origin of prokaryotic “species”: The taxonomy of halophilic. Saline Syst. 2008, 4, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzmann, P.D.; Stackebrandt, E.; Sanderson, K.; Volkman, J.K.; Cameron, D.E.; Stevenson, P.L.; McMeekin, T.A.; Burton, H.R. Halobacterium lacusprofundi sp. nov., a halophilic bacterium isolated from Deep Lake, Antarctica. Syst. Appl. Microbiol. 1988, 11, 20–27. [Google Scholar] [CrossRef]
- DasSarma, P.; Anton, B.P.; von Ehrenheim, H.A.L.; Roberts, R.J.; DasSarma, S. Complete Genome Sequence of an Extremely Halophilic Archaeon from Great Salt Lake, Halobacterium sp. GSL-19. Microbiol. Resour. Announc. 2021, 10, e00520-21. [Google Scholar] [CrossRef] [PubMed]
- Anton, B.P.; DasSarma, P.; Martinez, F.L.; DasSarma, S.L.; Al Madadha, M.; Roberts, R.J.; DasSarma, S. Genome Sequence of Salarchaeum sp. Strain JOR-1, an Extremely Halophilic Archaeon from the Dead Sea. Microbiol. Resour. Announc. 2020, 9, e01505-19. [Google Scholar] [CrossRef] [Green Version]
- DasSarma, P.; Anton, B.P.; DasSarma, S.; Laye, V.J.; Guzmán, D.; Roberts, R.J.; DasSarma, S. Genome Sequence and Methylation Patterns of Halorubrum sp. Strain BOL3-1, the First Haloarchaeon Isolated and Cultured from Salar de Uyuni, Bolivia. Microbiol. Resour. Announc. 2019, 8, e00386-19. [Google Scholar] [CrossRef] [Green Version]
- DasSarma, P.; Anton, B.P.; von Ehrenheim, H.A.L.; Martinez, F.L.; Guzmán, D.; Roberts, R.J.; DasSarma, S. Genome Sequence of Halobacterium sp. Strain BOL4-2, Isolated and Cultured from Salar de Uyuni, Bolivia. Microbiol. Resour. Announc. 2021, 10, e0104521. [Google Scholar] [CrossRef]
- DasSarma, P.; Anton, B.P.; DasSarma, S.L.; von Ehrenheim, H.A.L.; Martinez, F.L.; Guzmán, D.; Roberts, R.J.; DasSarma, S. Genome Sequence and Methylation Pattern of Haloterrigena salifodinae BOL5-1, an Extremely Halophilic Archaeon from a Bolivian Salt Mine. Microbiol. Resour. Announc. 2021, 10, e00275-21. [Google Scholar] [CrossRef]
- DasSarma, P.; Anton, B.P.; DasSarma, S.L.; Martinez, F.L.; Guzmán, D.; Roberts, R.J.; DasSarma, S. Genome Sequences and Methylation Patterns of Natrinema versiforme BOL5-4 and Natrinema pallidum BOL6-1, Two Extremely Halophilic Archaea from a Bolivian salt mine. Microbiol. Resour. Announc. 2019, 8, e00810-19. [Google Scholar] [CrossRef] [Green Version]
- Berquist, B.R.; Müller, J.A.; DasSarma, S. Genetic Systems for Halophilic Archaea. In Methods in Microbiology; Oren, A., Rainey, F., Eds.; Elsevier/Academic Press: Amsterdam, The Netherlands, 2006; Volume 35, pp. 649–680. [Google Scholar]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S.; Gascuel, O. NGPhylogeny.fr: New generation phylogenetic services for non-specialists. Nucleic Acids Res. 2019, 47, W260–W265. [Google Scholar] [CrossRef] [Green Version]
- DasSarma, S.L.; Capes, M.D.; DasSarma, P. HaloWeb: The haloarchaeal genomes database. Saline Syst. 2010, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M. Linking databases and organisms: GenomeNet resources in Japan. Trends Biochem. Sci. 1997, 22, 442–444. [Google Scholar] [CrossRef]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-PdbViewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef]
- Coker, J.A.; DasSarma, P.; Kumar, J.; Müller, J.A.; DasSarma, S. Transcriptional profiling of the model Archaeon Halobacterium sp. NRC-1: Responses to changes in salinity and temperature. Saline Syst. 2007, 3, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takao, M.; Kobayashi, T.; Oikawa, A.; Yasui, A. Tandem arrangement of photolyase and superoxide dismutase genes in Halobacterium halobium. J. Bacteriol. 1989, 171, 6323–6329. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, K.; Kim, S.T.; Walsh, C.; Sancar, A. Roles of FAD and 8-hydroxy-5-deazaflavin chromophores in photoreactivation by Anacystis nidulans DNA photolyase. J. Biol. Chem. 1992, 267, 15406–15411. [Google Scholar] [CrossRef]
- Glas, A.F.; Maul, M.J.; Cryle, M.; Barends, T.R.M.; Schneider, S.; Kaya, E.; Schlichting, I.; Carell, T. The archaeal cofactor F 0 is a light-harvesting antenna chromophore in eukaryotes. Proc. Natl. Acad. Sci. USA 2009, 106, 11540–11545. [Google Scholar] [CrossRef] [Green Version]
- Iwasa, T.; Tokutomi, S.; Tokunaga, F. Photoreactivation of Halobacterium halobium: Action spectrum and role of pigmentation. Photochem. Photobiol. 1988, 47, 267–270. [Google Scholar] [CrossRef]
- Park, H.W.; Kim, S.T.; Sancar, A.; Deisenhofer, J. Crystal structure of DNA photolyase from Escherichia coli. Science 1995, 268, 1866–1872. [Google Scholar] [CrossRef]
- Weber, S. Light-driven enzymatic catalysis of DNA repair: A review of recent biophysical studies on photolyase. Biochim. Biophys. Acta 2005, 1707, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Wit, L.E.A.; Eker, A.P.M. 8-Hydroxy-5-deazaflavin-dependent electron transfer in the extreme halophile Halobacterium cutirubrum. FEMS Microbiol. Lett. 1987, 48, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Hescox, M.A.; Carlberg, D.M. Photoreactivation in Halobacterium cutirubrum. Can. J. Microbiol. 1972, 18, 981–985. [Google Scholar] [CrossRef] [PubMed]
- DasSarma, S.; Kennedy, S.P.; Berquist, B.; Ng, W.V.; Baliga, N.S.; Spudich, J.L.; Krebs, M.P.; Eisen, J.A.; Johnson, C.H.; Hood, L. Genomic perspective on the photobiology of Halobacterium species NRC-1, a phototrophic, phototactic, and UV-tolerant haloarchaeon. Photosynth. Res. 2001, 70, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, S.P.; Ng, W.V.; Salzberg, S.L.; Hood, L.; DasSarma, S. Understanding the Adaptation of Halobacterium Species NRC-1 to Its Extreme Environment through Computational Analysis of Its Genome Sequence. Genome Res. 2001, 11, 1641–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DasSarma, P.; Capes, M.D.; DasSarma, S. Comparative Genomics of Halobacterium Strains from Diverse Locations. In Microbial Diversity in the Genomic Era, 1st ed.; Das, S., Dash, H.R., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 285–322. [Google Scholar]
- Karan, R.; DasSarma, P.; Balcer-Kubiczek, E.; Weng, R.R.; Liao, C.C.; Goodlett, D.R.; Ng, W.V.; DasSarma, S. Bioengineering radioresistance by overproduction of RPA, a mammalian-type single-stranded DNA-binding protein, in a halophilic archaeon. Appl. Microbiol. Biotechnol. 2013, 98, 1737–1747. [Google Scholar] [CrossRef] [Green Version]
- Rousseau, B.J.G.; Shafei, S.; Migliore, A.; Stanley, R.J.; Beratan, D.N. Determinants of Photolyase’s DNA Repair Mechanism in Mesophiles and Extremophiles. J. Am. Chem. Soc. 2018, 140, 2853–2861. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Haloarchaea | Source | Genome Size (bp) | GC% | Ref. |
|---|---|---|---|---|
| Halobacterium sp. NRC-1 | San Francisco Bay, USA | 2,571,010 | 65.9 | [27] |
| Halobacterium sp. BOL4-2 | Salar de Uyuni, Bolivia | 2,428,492 | 66.3 | [38] |
| Halobacterium sp. GSL-19 | Great Salt Lake, USA | 2,326,224 | 66.7 | [35] |
| Salarchaeum sp. JOR-1 | Dead Sea, Jordan | 2,523,371 | 66.2 | [36] |
| Halorubrum lacusprofundi | Deep Lake, Antarctica | 3,692,576 | 64.0 | [30] |
| Halorubrum sp. BOL3-1 | Salar de Uyuni, Bolivia | 3,654,453 | 65.9 | [37] |
| Haloterrigena salifodinae BOL5-1 | Tarija mine, Bolivia | 5,087,240 | 63.4 | [39] |
| Natrinema pallidum BOL6-1 | Tarija mine, Bolivia | 3,778,093 | 64.3 | [40] |
| Natrinema versiforme BOL5-4 | Tarija mine, Bolivia | 4,674,473 | 63.4 | [40] |
| SEL AA * | Interaction with | SEL | NRC-1 † | JOR-1 | Hla | BOL3-1 | BOL5-1 | BOL6-1 |
|---|---|---|---|---|---|---|---|---|
| 50 | HDF | R | R | R | R | R | V | R |
| 139 | DNA | S | T | T | T | T | T | P |
| 144 | DNA | P | P | P | P | P | H | H |
| 147 | DNA | V | V | V | V | V | D | V |
| 148 | DNA | Y | Y | Y | Y | Y | F | F |
| 149 | DNA | G | T | T | S | T | S | S |
| 150 | DNA | P | Y | Y | Y | Y | D | Y |
| 242 | FAD | L | L | V | M | L | L | L |
| 279 | FAD | W | F | F | Y | F | F | F |
| 282 | DNA | E | Q | Q | Q | Q | R | Q |
| 384 | FAD | A | N | N | N | N | H | N |
| 385 | FAD | N | D | D | D | N | A | D |
| 398 | DNA | D | D | D | D | D | N | D |
| 413 | DNA | K | R | D | R | R | T | E |
| 460 | DNA | Q | R | R | R | R | H | R |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nag, S.; DasSarma, P.; Crowley, D.J.; Hamawi, R.; Tepper, S.; Anton, B.P.; Guzmán, D.; DasSarma, S. Genomic Analysis of Haloarchaea from Diverse Environments, including Permian Halite, Reveals Diversity of Ultraviolet Radiation Survival and DNA Photolyase Gene Variants. Microorganisms 2023, 11, 607. https://doi.org/10.3390/microorganisms11030607
Nag S, DasSarma P, Crowley DJ, Hamawi R, Tepper S, Anton BP, Guzmán D, DasSarma S. Genomic Analysis of Haloarchaea from Diverse Environments, including Permian Halite, Reveals Diversity of Ultraviolet Radiation Survival and DNA Photolyase Gene Variants. Microorganisms. 2023; 11(3):607. https://doi.org/10.3390/microorganisms11030607
Chicago/Turabian StyleNag, Sagorika, Priya DasSarma, David J. Crowley, Rafael Hamawi, Samantha Tepper, Brian P. Anton, Daniel Guzmán, and Shiladitya DasSarma. 2023. "Genomic Analysis of Haloarchaea from Diverse Environments, including Permian Halite, Reveals Diversity of Ultraviolet Radiation Survival and DNA Photolyase Gene Variants" Microorganisms 11, no. 3: 607. https://doi.org/10.3390/microorganisms11030607