
Fungal Pathogens Associated with Aerial Symptoms of Avocado (Persea americana Mill.) in Tenerife (Canary Islands, Spain) Focused on Species of the Family Botryosphaeriaceae
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Fungal Isolation
2.2. Morphological and Molecular Identification of Fungal Isolates
2.3. Pathogenicity Tests
3. Results
3.1. Sampling and Fungal Isolation
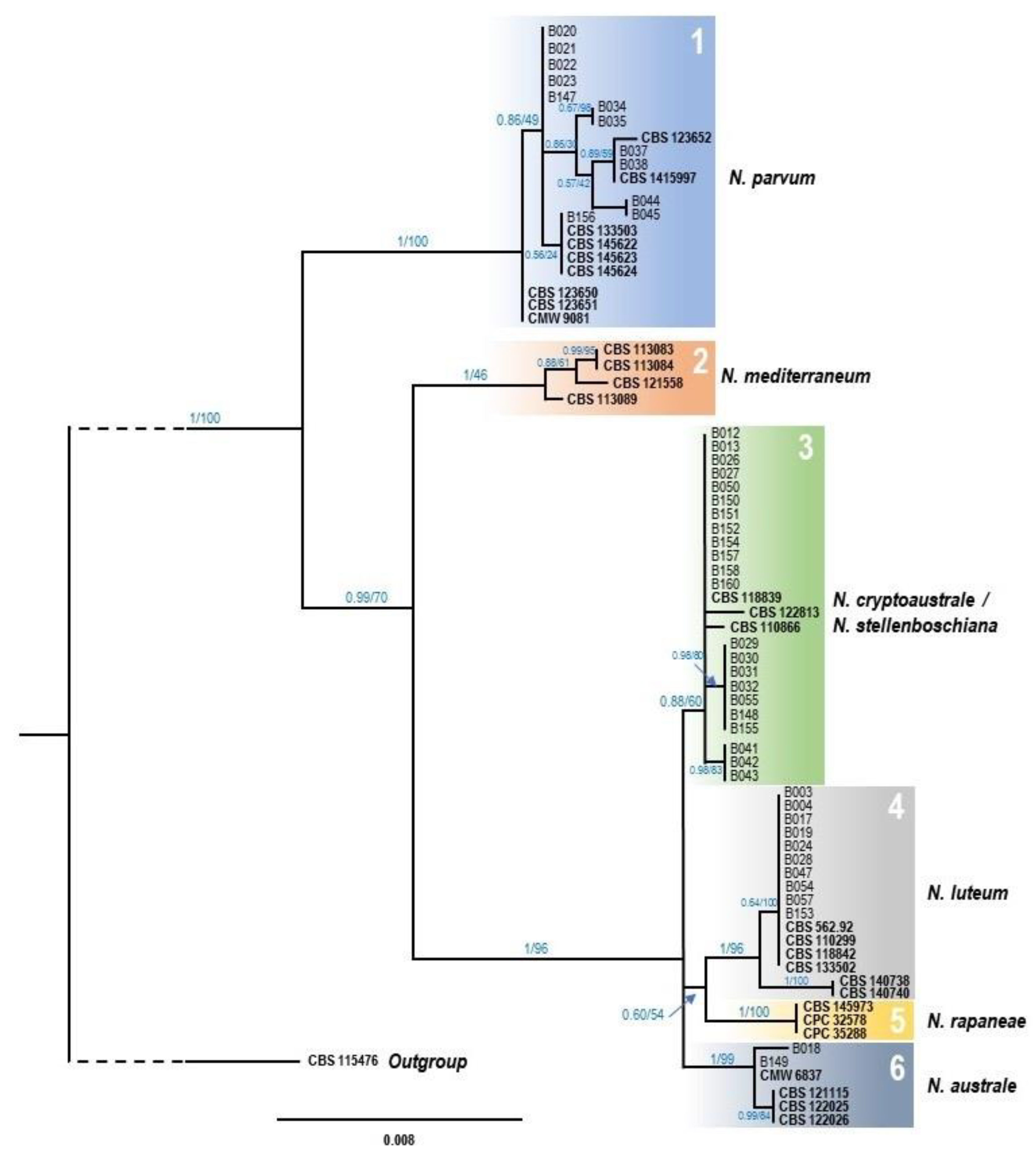
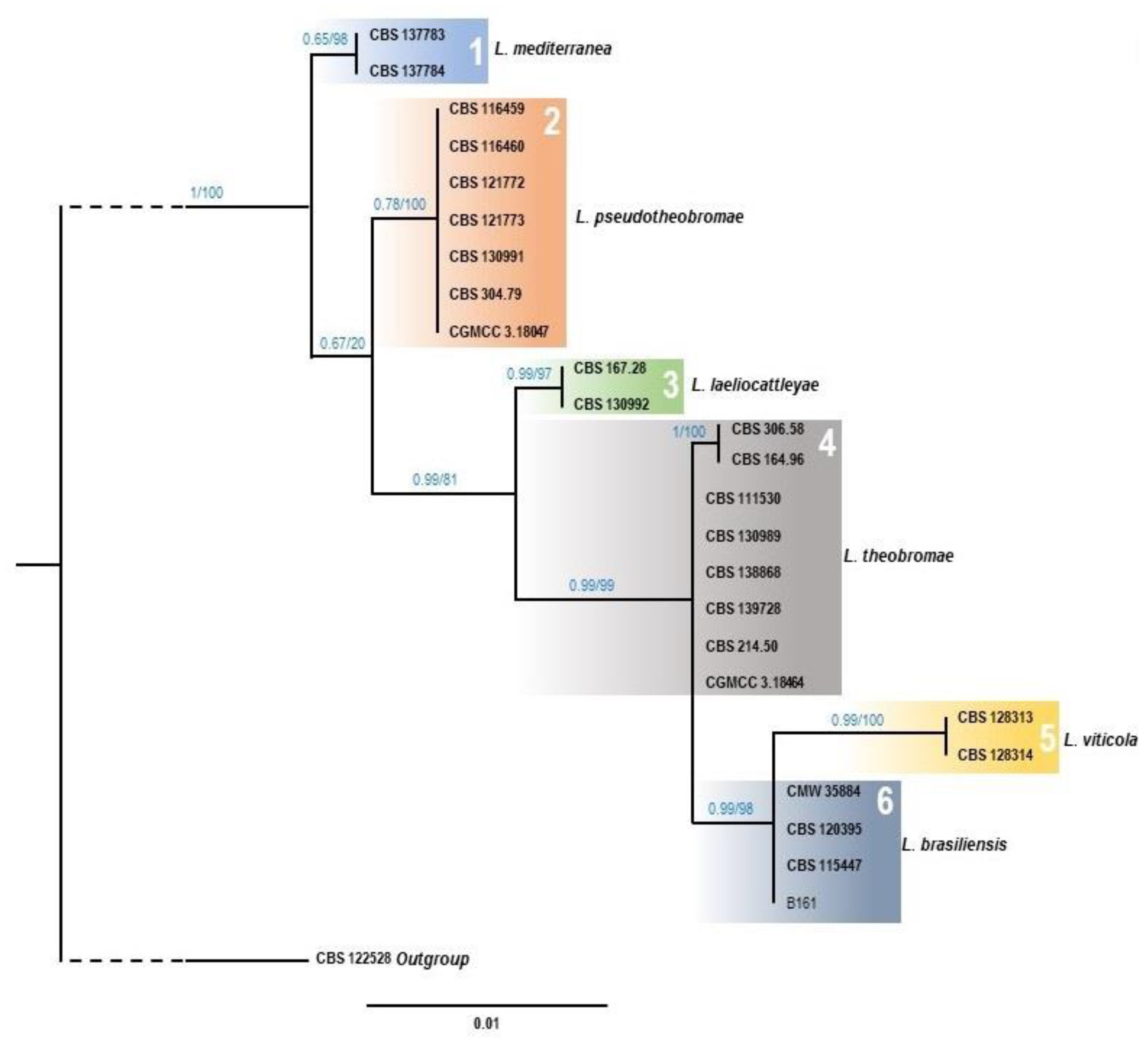
3.2. Morphological and Molecular Identification of Fungal Isolates
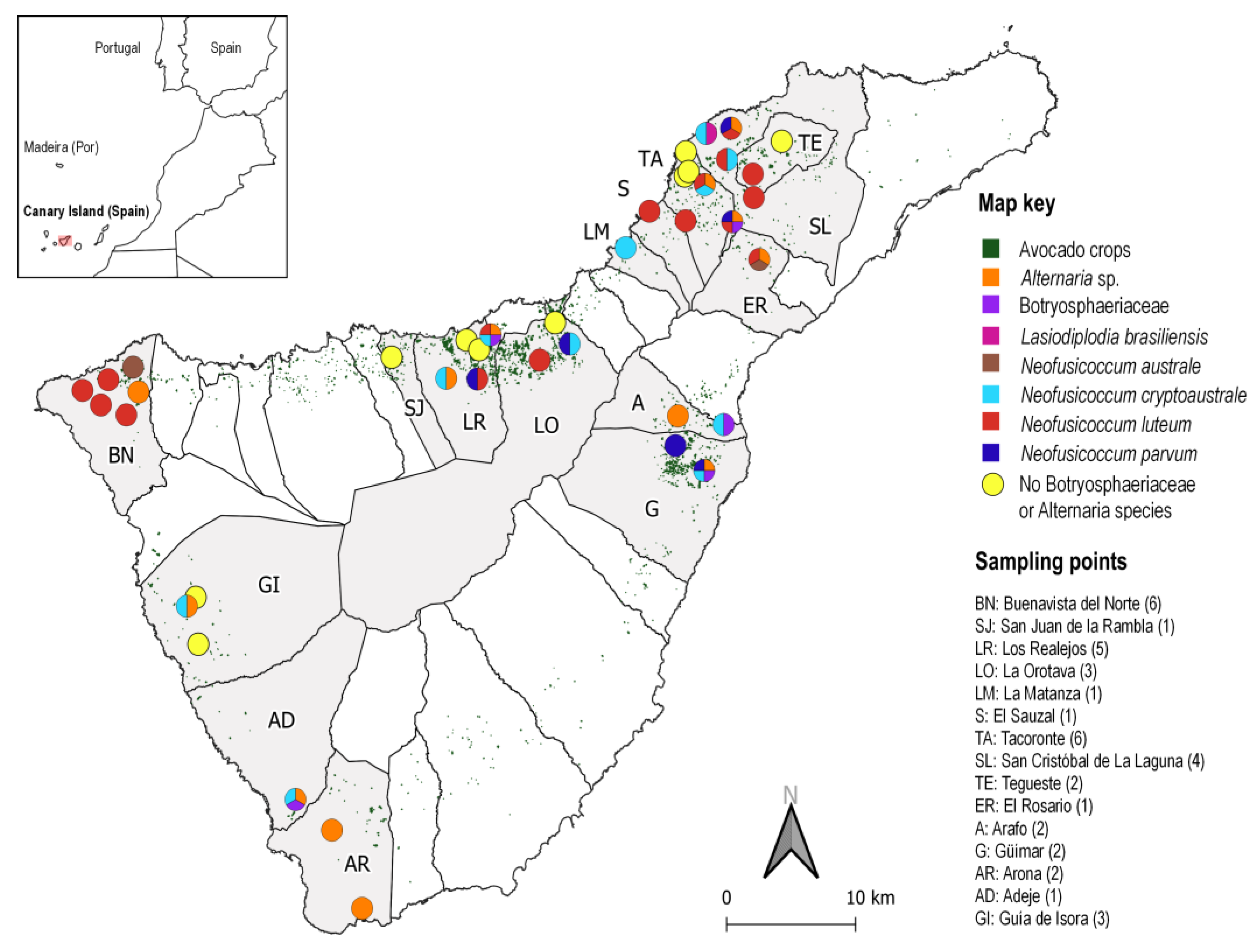
3.3. Symptoms and Distribution of Botryosphaeriaceae Species Associated to Avocado in Tenerife
3.4. Pathogenicity Tests
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ISTAC Estadística Agraria de Canarias/Series Anuales de Agricultura. Municipios, Islas y Provincias de Canarias. 1999–2021. Available online: http://www.gobiernodecanarias.org/istac/jaxi-istac/tabla.do?uripx=urn:uuid:803af13a-04f6-4789-8e78-48f5fdff64a7&uripub=urn:uuid:ef5f2e5c-e2c4-4c1d-b5ed-c20fe946ce6f%0A%0A (accessed on 30 December 2022).
- MAPA Encuesta Sobre Superficies y Rendimientos Cultivos (ESYRCE). Encuesta de Marco de Áreas de España. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/agricultura/esyrce/default.aspx (accessed on 30 December 2022).
- Auger, J.; Palma, F.; Pérez, I.; Esterio, M. First Report of Neofusicoccum australe (Botryosphaeria australis), as a Branch Dieback Pathogen of Avocado Trees in Chile. Plant Dis. 2013, 97, 842. [Google Scholar] [CrossRef]
- Zea-Bonilla, T.; González-Sánchez, M.A.; Martín-Sánchez, P.M.; Pérez-Jiménez, R.M. Avocado Dieback Caused by Neofusicoccum parvum in the Andalucia Region, Spain. Plant Dis. 2007, 91, 1052. [Google Scholar] [CrossRef] [PubMed]
- Arjona-Girona, I.; Ruano-Rosa, D.; López-Herrera, C.J. Identification, Pathogenicity and Distribution of the Causal Agents of Dieback in Avocado Orchards in Spain. Span. J. Agric. Res. 2019, 17, e1003. [Google Scholar] [CrossRef] [Green Version]
- Valencia, A.L.; Gil, P.M.; Latorre, B.A.; Rosales, I.M. Characterization and Pathogenicity of Botryosphaeriaceae Species Obtained from Avocado Trees with Branch Canker and Dieback and from Avocado Fruit with Stem End Rot in Chile. Plant Dis. 2019, 103, 996–1005. [Google Scholar] [CrossRef]
- Hartill, W.F.T.; Everett, K.R. Inoculum Sources and Infection Pathways of Pathogens Causing Stem-End Rots of ‘Hass’ Avocado (Persea americana). N. Zeal. J. Crop Hortic. Sci. 2002, 30, 249–260. [Google Scholar] [CrossRef]
- Twizeyimana, M.; McDonald, V.; Mayorquin, J.S.; Wang, D.H.; Na, F.; Akgül, D.S.; Eskalen, A. Effect of Fungicide Application on the Management of Avocado Branch Canker (Formerly Dothiorella Canker) in California. Plant Dis. 2013, 97, 897–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, V.; Lynch, S.; Eskalen, A. First Report of Neofusicoccum australe, N. luteum, and N. parvum Associated with Avocado Branch Canker in California. Plant Dis. 2009, 93, 967. [Google Scholar] [CrossRef]
- McDonald, V.; Eskalen, A. Botryosphaeriaceae Species Associated with Avocado Branch Cankers in California. Plant Dis. 2011, 95, 1465–1473. [Google Scholar] [CrossRef] [Green Version]
- Alama, I.; Maldonado, E.; Rodríguez-Gálvez, E. Lasiodiplodia theobromae Affect the Cultivation of Palto (Persea americana) under the Conditions of Piura, Peru. Universalia 2006, 11, 4–13. [Google Scholar]
- Hartill, W.F.T. Post-Harvest Diseases of Avocado Fruits in New Zealand. N. Zeal. J. Crop Hortic. Sci. 1991, 19, 297–304. [Google Scholar] [CrossRef]
- Darvas, J.; Kotze, J. Fungi Associated with Pre-and Postharvest Diseases of Avocado Fruit at Westfalia Estate, South Africa. Phytophylactica 1987, 19, 83–85. [Google Scholar]
- Twizeyimana, M.; Förster, H.; McDonald, V.; Wang, D.H.; Adaskaveg, J.E.; Eskalen, A. Identification and Pathogenicity of Fungal Pathogens Associated with Stem-End Rot of Avocado in California. Plant Dis. 2013, 97, 1580–1584. [Google Scholar] [CrossRef] [PubMed]
- Denman, S.; Crous, P.W.; Taylor, J.E.; Kang, J.C.; Pascoe, I.; Wingfield, M.J. An Overview of the Taxonomic History of Botryosphaeria, and a Re-Evaluation of Its Anamorphs Based on Morphology and ITS RDNa Phylogeny. Stud. Mycol. 2000, 2000, 129–140. [Google Scholar]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and Species Known from Culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A.J.; Phillips, A.J.L.; Li, X.H.; Hyde, K.D. Botryosphaeriaceae: Current Status of Genera and Species. Mycosphere 2016, 7, 1001–1073. [Google Scholar] [CrossRef]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as Endophytes and Latent Pathogens of Woody Plants: Diversity, Ecology and Impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Dann, E.; Prabhakaran, A.; Bransgrove, K. Panicle Blight (Flower Dieback). Available online: https://avocado.org.au/public-articles/ta31v3panicle/ (accessed on 30 December 2022).
- Burgess, T.I.; Tan, Y.P.; Garnas, J.; Edwards, J.; Scarlett, K.A.; Shuttleworth, L.A.; Daniel, R.; Dann, E.K.; Parkinson, L.E.; Dinh, Q. Current Status of the Botryosphaeriaceae in Australia. Australas. Plant Pathol. 2019, 48, 35–44. [Google Scholar] [CrossRef]
- Peterson, R.A. Susceptibility of Fuerte Avocado Fruit at Various Stages of Growth, to Infection by Anthracnose and Stem End Rot Fungi. Aust. J. Exp. Agric. 1978, 18, 158–160. [Google Scholar] [CrossRef] [Green Version]
- Firmino, A.C.; Fischer, I.H.; Tozze Júnior, H.J.; Dias Rosa, D.; Luiz Furtado, E. Identificação de Espécies de Fusicoccum Causadoras de Podridão Em Frutos de Abacate. Summa Phytopathol. 2016, 42, 100–102. [Google Scholar] [CrossRef] [Green Version]
- Montealegre, J.R.; Ramírez, M.; Riquelme, D.; Armengol, J.; León, M.; Pérez, L.M. First Report of Neofusicoccum australe in Chile Causing Avocado Stem-End Rot. Plant Dis. 2016, 100, 2532. [Google Scholar] [CrossRef]
- Tapia, L.; Larach, A.; Riquelme, N.; Guajardo, J.; Besoain, X. First Report of Neofusicoccum luteum Causing Stem-End Rot Disease on Avocado Fruits in Chile. Plant Dis. 2020, 104, 2027. [Google Scholar] [CrossRef]
- Qiu, F.; Xu, G.; Zhou, J.; Zheng, F.Q.; Zheng, L.; Miao, W.G.; Wang, W.L.; Xie, C.P. First Report of Botryosphaeria dothidea Causing Stem-End Rot in Avocado (Persea americana) in China. Plant Dis. 2019, 104, 286. [Google Scholar] [CrossRef]
- Ramirez-Gil, J.G. Avocado Wilt Complex Disease, Implications and Management in Colombia. Rev. Fac. Nac. Agron. Colomb. 2018, 71, 8525–8541. [Google Scholar] [CrossRef]
- Ramírez-Gil, J.G.; López, J.H.; Henao-Rojas, J.C. Causes of Hass Avocado Fruit Rejection in Preharvest, Harvest, and Packinghouse: Economic Losses and Associated Variables. Agronomy 2019, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Kebede, M.; Belay, A. Fungi Associated With Post-Harvest Avocado Fruit Rot at Jimma Town, Southwestern Ethiopia. J. Plant Pathol. Microbiol. 2019, 10, 2. [Google Scholar] [CrossRef] [Green Version]
- Guarnaccia, V.; Aiello, D.; Cirvilleri, G.; Polizzi, G.; Susca, A.; Epifani, F.; Perrone, G. Characterisation of Fungal Pathogens Associated with Stem-End Rot of Avocado Fruit in Italy. Acta Hortic. 2016, 1144, 133–139. [Google Scholar] [CrossRef]
- Molina-Gayosso, E.; Silva-Rojas, H.V.; García-Morales, S.; Ávila-Quezada, G. First Report of Black Spots on Avocado Fruit Caused by Neofusicoccum parvum in Mexico. Plant Dis. 2011, 96, 287. [Google Scholar] [CrossRef]
- Trakunyingcharoen, T.; Cheewangkoon, R.; To-anun, C. Phylogenetic Study of the Botryosphaeriaceae Species Associated with Avocado and Para Rubber in Thailand. Chiang Mai J. Sci. 2015, 42, 104–116. [Google Scholar]
- Ni, H.F.; Liou, R.F.; Hung, T.H.; Chen, R.S.; Yang, H.R. First Report of a Fruit Rot Disease of Avocado Caused by Neofusicoccum mangiferae. Plant Dis. 2009, 93, 760. [Google Scholar] [CrossRef]
- Ni, H.; Chuang, M.; Hsu, S.; Lai, S.; Yang, H. Survey of Botryosphaeria spp., Causal Agents of Postharvest Disease of Avocado, in Taiwan. J. Taiwan Agric. Res. 2011, 60, 157–166. [Google Scholar]
- Truett, G.E.; Heeger, P.; Mynatt, R.L.; Truett, A.A.; Walker, J.A.; Warman, M.L. Preparation of PCR-Quality Mouse Genomic Dna with Hot Sodium Hydroxide and Tris (HotSHOT). Biotechniques 2000, 29, 52–54. [Google Scholar] [CrossRef] [PubMed]
- Collado-Romero, M.; Mercado-Blanco, J.; Olivares-García, C.; Valverde-Corredor, A.; Jiménez-Díaz, R.M. Molecular Variability within and among Verticillium dahliae Vegetative Compatibility Groups Determined by Fluorescent Amplified Fragment Length Polymorphism and Polymerase Chain Reaction Markers. Phytopathology 2006, 96, 485–495. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. PCR Protoc. 1990, 315–322. [Google Scholar] [CrossRef]
- Alves, A.; Crous, P.W.; Correia, A.; Phillips, A.J.L. Morphological and Molecular Data Reveal Cryptic Speciation in Lasiodiplodia theobromae. Fungal Divers. 2008, 28, 1–13. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of Primer Sets Designed for Use with the PCR to Amplify Conserved Genes from Filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakalidis, M.L.; Hardy, G.E.S.J.; Burgess, T.I. Use of the Genealogical Sorting Index (GSI) to Delineate Species Boundaries in the Neofusicoccum Parvum-neofusicoccum Ribis Species Complex. Mol. Phylogenet. Evol. 2011, 60, 333–344. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef]
- Mussi-Dias, V.; Valmir dos Santos, A.; das Graças Machado Freire, M. Pycnidia and Conidia Quantification of Lasiodiplodia Using a Culture Medium Enriched with Sugarcane Bagasse. J. Agric. Res. 2016, 2, 1–16. [Google Scholar]
- Marques, M.W.; Lima, N.B.; De Morais, M.A.; Barbosa, M.A.G.; Souza, B.O.; Michereff, S.J.; Phillips, A.J.L.; Câmara, M.P.S. Species of Lasiodiplodia Associated with Mango in Brazil. Fungal Divers. 2013, 61, 181–193. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Cheewangkoon, R.; van der Bank, M.; Swart, W.J.; Stchigel, A.M.; Cano-Lira, J.F.; Roux, J.; Madrid, H.; et al. Fungal Planet Description Sheets: 154–213. Persoonia-Mol. Phylogeny Evol. Fungi 2013, 31, 188–296. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Groenewald, J.Z.; Cheewangkoon, R.; Jami, F.; Abdollahzadeh, J.; Lombard, L.; Crous, P.W. Families, Genera, and Species of Botryosphaeriales. Fungal Biol. 2017, 121, 322–346. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES Science Gateway. In Proceedings of the 1st Conference of the Extreme Science and Engineering Discovery Environment on Bridging from the eXtreme to the Campus and beyond—XSEDE ’12, Chicago, IL, USA, 16–20 July 2012; ACM Press: New York, NY, USA, 2012; p. 1. [Google Scholar]
- Tavaré, S. Some Probabilistic and Statistical Problems in the Analysis of DNA Sequences. Am. Math. Soc. Lect. Math. Life Sci. 1986, 17, 57–86. [Google Scholar]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Guarnaccia, V.; Polizzi, G.; Papadantonakis, N.; Gullino, M.L. Neofusicoccum Species Causing Branch Cankers on Avocado in Crete (Greece). J. Plant Pathol. 2020, 102, 1251–1255. [Google Scholar] [CrossRef]
- Eskalen, A.; McDonald, V. Geographical Distribution of Botryosphaeriaceae and Phomopsis/Diaporthe Canker Pathogens of Avocado in California. Calif. Avocado Soc. 2010, 93, 87–98. [Google Scholar]
- Gallo Llobet, L.; Hernández Hernández, J.M.; del Jaizme Vega, M.C.; Sala Mayato, L. Especies Fúngicas Enontradas En Los Cultivos de Canarias En El Período 1974–1984. In Proceedings of the III Congreso Nacional de Fitopatología; Sociedad Española de Fitopatología, Ed.; The Canary Islands: Tenerife, Spain, 1984; pp. 43–50. [Google Scholar]
- Márquez, M.P.; Santana, M.L.; Albalat, A.R.; Armengol, J. Muestreo e Identificación de Hongos Asociados a Las Enfermedades de La Madera de La Vid En Lanzarote (Canarias). Enoviticultura 2020, 65, 16–26. [Google Scholar]
- Benito Hernández, P. Botryosphaeriaceae Asociadas al Decaimiento de Ficus microcarpa en Gran Canaria. Master Thesis, Universitat Politècnica de València, Valencia, Spain, 2018. [Google Scholar]
- Mojeremane, K.; Lebenya, P.; Plessis, I.L.D.; Van Der Rijst, M.; Mostert, L.; Armengol, J.; Halleen, F. Cross Pathogenicity of Neofusicoccum Australe and Neofusicoccum stellenboschiana on Grapevine and Selected Fruit and Ornamental Trees. Phytopathol. Mediterr. 2020, 59, 581–593. [Google Scholar] [CrossRef]
- Sessa, L.; Abreo, E.; Bettucci, L.; Lupo, S. Botryosphaeriaceae Species Associated with Wood Diseases of Stone and Pome Fruits Trees: Symptoms and Virulence across Different Hosts in Uruguay. Eur. J. Plant Pathol. 2016, 146, 519–530. [Google Scholar] [CrossRef]
- Van Dyk, M.; Spies, C.F.J.; Mostert, L.; Halleen, F. Survey of Trunk Pathogens in South African Olive Nurseries. Plant Dis. 2021, 105, 1630–1639. [Google Scholar] [CrossRef] [PubMed]
- Spies, C.F.J.; Mostert, L.; Carlucci, A.; Moyo, P.; van Jaarsveld, W.J.; du Plessis, I.L.; van Dyk, M.; Halleen, F. Dieback and Decline Pathogens of Olive Trees in South Africa. Persoonia Mol. Phylogeny Evol. Fungi 2020, 45, 196–220. [Google Scholar] [CrossRef] [PubMed]
- Linaldeddu, B.T.; Maddau, L.; Franceschini, A.; Alves, A.; Phillips, A.J.L. Botryosphaeriaceae Species Associated with Lentisk Dieback in Italy and Description of Diplodia insularis sp. Nov. Mycosphere 2016, 7, 962–977. [Google Scholar] [CrossRef]
- Osorio, J.A.; Crous, C.J.; de Beer, Z.W.; Wingfield, M.J.; Roux, J. Endophytic Botryosphaeriaceae, Including Five New Species, Associated with Mangrove Trees in South Africa. Fungal Biol. 2017, 121, 361–393. [Google Scholar] [CrossRef] [Green Version]
- Mahamedi, A.E.; Phillips, A.J.L.; Lopes, A.; Djellid, Y.; Arkam, M.; Eichmeier, A.; Zitouni, A.; Alves, A.; Berraf-Tebbal, A. Diversity, Distribution and Host Association of Botryosphaeriaceae Species Causing Oak Decline across Different Forest Ecosystems in Algeria. Eur. J. Plant Pathol. 2020, 158, 745–765. [Google Scholar] [CrossRef]
- Netto, M.S.B.; Assunção, I.P.; Lima, G.S.A.; Marques, M.W.; Lima, W.G.; Monteiro, J.H.A.; de Queiroz Balbino, V.; Michereff, S.J.; Phillips, A.J.L.; Câmara, M.P.S. Species of Lasiodiplodia Associated with Papaya Stem-End Rot in Brazil. Fungal Divers. 2014, 67, 127–141. [Google Scholar] [CrossRef]
- Correia, K.C.; Silva, M.A.; de Morais, M.A.; Armengol, J.; Phillips, A.J.L.; Câmara, M.P.S.; Michereff, S.J. Phylogeny, Distribution and Pathogenicity of Lasiodiplodia Species Associated with Dieback of Table Grape in the Main Brazilian Exporting Region. Plant Pathol. 2016, 65, 92–103. [Google Scholar] [CrossRef]
- Cardoso, J.E.; Lima, J.S.; Viana, F.M.P.; Ootani, M.A.; Araújo, F.S.A.; Fonseca, W.L.; Lima, C.S.; Martins, M.V.V. First Report of Lasiodiplodia brasiliense Causing Postharvest Fruit Rot of Custard Apple (Annona squamosa) in Brazil. Plant Dis. 2017, 101, 1542. [Google Scholar] [CrossRef]
- Martins, M.V.V.; Lima, J.S.; Hawerroth, F.J.; Ootani, M.A.; Araujo, F.S.A.; Cardoso, J.E.; Serrano, L.A.L.; Viana, F.M.P. First Report of Lasiodiplodia brasiliense Causing Disease in Apple Trees in Brazil. Plant Dis. 2018, 102, 1027. [Google Scholar] [CrossRef]
- Zhang, H.; Wei, Y.X.; Qi, Y.X.; Pu, J.J.; Liu, X.M. First Report of Lasiodiplodia brasiliense Associated with Stem-End Rot of Mango in China. Plant Dis. 2018, 102, 679. [Google Scholar] [CrossRef]
- Urbez-Torres, J.R.; Peduto, F.; Striegler, R.K.; Urrea-Romero, K.E.; Rupe, J.C.; Cartwright, R.D.; Gubler, W.D. Characterization of Fungal Pathogens Associated with Grapevine Trunk Diseases in Arkansas and Missouri. Fungal Divers. 2012, 52, 169–189. [Google Scholar] [CrossRef]
- Menge, J.A.; Ploetz, R.C. Diseases of Avocado. In Diseases of Tropical Fruit Crops; CABI: Wallingford, UK, 2003; pp. 35–71. [Google Scholar]
- Rodríguez-Gálvez, E.; Hilário, S.; Lopes, A.; Alves, A. Diversity and Pathogenicity of Lasiodiplodia and Neopestalotiopsis Species Associated with Stem Blight and Dieback of Blueberry Plants in Peru. Eur. J. Plant Pathol. 2020, 157, 89–102. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Reference | Symptoms | Botryosphaeriaceae Species | Non-Botryosphaeriaceae Species | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant | Fruit | ||||||||||||||||||||||
| Branch or Trunk Canker | Dieback | Stem-End Rot | Black Spots | Botryosphaeria dothidea | Diplodia mutila | Diplodia pseudoseriata | Diplodia seriata | Dothiorella aromatica | Dothiorella dominicana | Dothiorella iberica | Dothiorella sp. | Lasiodiplodia pseudotheobromae | Lasiodiplodia theobromae | Neofusiccocum australe | Neofusiccocum luteum | Neofusiccocum mangiferae | Neofusiccocum mediterraneum | Neofusiccocum nonquaesitum | Neofusiccocum parvum | Neofusiccocum sp. | |||
| Australia | [20] | X | X | X | X | X | |||||||||||||||||
| Australia | [12] | X | X | X | X | X | |||||||||||||||||
| Australia | [21] | X | X | X | Cg | ||||||||||||||||||
| Brasil | [22] | X | X | X | X | ||||||||||||||||||
| Chile | [3] | X | X | ||||||||||||||||||||
| Chile | [23] | X | X | ||||||||||||||||||||
| Chile | [6] | X | X | X | X | X | X | X | X | X | X | X | |||||||||||
| Chile | [24] | X | X | X | |||||||||||||||||||
| China | [25] | X | X | ||||||||||||||||||||
| Colombia | [26] | X | X | X | |||||||||||||||||||
| Colombia | [27] | X | X | X | X | Cg, Phs, Pp, Ps | |||||||||||||||||
| Ethiopia | [28] | X | X | Af, An, Cg, Cls, Fs, Pes | |||||||||||||||||||
| Italy | [29] | X | X | Cf, Cg, | |||||||||||||||||||
| Mexico | [30] | X | X | ||||||||||||||||||||
| New Zealand | [12] | X | X | X | X | Ca, Cg, Phs | |||||||||||||||||
| New Zealand | [7] | X | X | X | X | X | Ca, Cg, Phs | ||||||||||||||||
| Peru | [11] | X | X | X | X | As, Cos, Fs | |||||||||||||||||
| South Africa | [13] | X | X | X | Cg, Cls, Np, Phs, Pms, Pp, Ps | ||||||||||||||||||
| Spain | [4] | X | X | ||||||||||||||||||||
| Spain | [5] | X | X | X | X | X | X | X | Cg | ||||||||||||||
| Thailandia | [31] | X | X | X | X | ||||||||||||||||||
| Taiwan | [32] | X | X | ||||||||||||||||||||
| Taiwan | [33] | X | X | X | X | X | X | ||||||||||||||||
| USA | [9] | X | X | X | X | X | |||||||||||||||||
| USA | [10] | X | X | X | X | X | X | X | Cos, Fs, Phs | ||||||||||||||
| USA | [8] | X | X | X | X | X | Phs | ||||||||||||||||
| USA | [14] | X | X | Cg, Phs | |||||||||||||||||||
| Genbank Accessions | |||||
|---|---|---|---|---|---|
| Species | Nº | Isolates 1 | ITS | tef1 | tub2 |
| N. australe | 2 | B018, B149 | OP788200 | OQ236719 | OQ181384 |
| N. cryptoaustrale/stellenboschiana | 12 | B012, B013, B026, B027, B050, B150, B151, B152, B154, B157, B158, B160 | OQ176234 | OQ236715 | OQ181395 |
| 7 | B029, B030, B031, B032, B055, B148, B155 | OP788375 | OQ236717 | OQ181394 | |
| 3 | B041, B042, B043 | OP788376 | OQ236716 | OQ181393 | |
| N. luteum | 9 | B003, B004, B017, B019, B028, B047, B054, B057, B153 | OP788373 | OQ236713 | OQ181386 |
| 1 | B024 | OP788372 | OQ236714 | OQ181387 | |
| N. parvum | 5 | B020, B021, B022, B023, B147 | OP788194 | OQ236708 | OQ181388 |
| 2 | B034, B035 | OP788193 | OQ236709 | OQ181389 | |
| 2 | B037, B038 | OP788192 | OQ236710 | OQ181390 | |
| 2 | B044, B045 | OP788176 | OQ236711 | OQ181391 | |
| 1 | B156 | OQ176235 | OQ236712 | OQ181392 | |
| L. brasiliensis | 1 | B161 | OP788199 | OQ236718 | OQ181385 |
| Species | Isolate | Apparition of Pycnidia/Conidia (Days) | ||
|---|---|---|---|---|
| SC-P | SC | WA | ||
| N. australe | B018 | 14/NP | 21/NP | 14/NP |
| N. cryptoaustrale/stellenboschiana | B012 | 14/NP | 14/NP | ND |
| B026 | 14/NP | 14/NP | 14/21 | |
| B030 | 21/NP | 14/NP | ND | |
| N. luteum | B003 | 21/NP | 21/42 | 14/21 |
| B017 | 14/NP | 14/14 | ND | |
| B024 | 14/NP | 14/28 | ND | |
| N. parvum | B020 | 28/NP | 28/42 | 14/NP |
| B022 | NP/NP | NP/NP | 21/21 | |
| Species | Isolates | Values Obtained in This Study | References | |||
|---|---|---|---|---|---|---|
| Length (L) (µm) | Width (W) (µm) | L/W Ratio | Length (L) (µm) | Width (W) (µm) | ||
| L. brasiliensis | B161 | 23.49 ± 1.42 | 14.95 ± 1.18 | 1.57 | 25.1–27.3 1 | 13.3–14.79 1 |
| N. cryptoaustrale/stellenboschiana | B026 | 20.44 ± 1.41 | 6.02 ± 0.51 | 3.4 | (18–) 20.5–21 (–26.5) 2 (17–) 19–21 (−22) 3 | 5–6 (–6.5) 2 (4.5–) 5.5–6 3 |
| N. luteum | B003 | 18.60 ± 2.72 | 5.93 ± 0.60 | 3.14 | (15–) 16.5–22.5 (–24) 4 | 4.5–6 (–7.5) 4 |
| B017 | 21.63 ± 1.55 | 6.34 ± 0.69 | 3.41 | |||
| B024 | 18.34 ± 1.56 | 5.50 ± 0.38 | 3.33 | |||
| N. parvum | B020 | 14.97 ± 1.05 | 5.70 ± 0.65 | 2.63 | (12–) 13.5–21 (–24) 4 | 4–6 (–10) 4 |
| Species | Isolate | Mean Lesion Length (cm) | Standard Deviation (cm) |
|---|---|---|---|
| Control | - | 0.0 | 0.0 |
| N. australe | B018 | 4.8 | 0.2 |
| N. cryptoaustrale/stellenboschiana | B043 1 | 3.7/2.5 | 3.5/1.3 |
| B050 | 2.4 | 0.3 | |
| N. luteum | B004 | 4.3 | 0.3 |
| B047 | 5.1 | 1.2 | |
| N. parvum | B034 | 5.6 | 1.2 |
| B045 | 1.4 | 0.7 | |
| B113 1 | 3.4/3.9 | 0.8/2.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández, D.; García-Pérez, O.; Perera, S.; González-Carracedo, M.A.; Rodríguez-Pérez, A.; Siverio, F. Fungal Pathogens Associated with Aerial Symptoms of Avocado (Persea americana Mill.) in Tenerife (Canary Islands, Spain) Focused on Species of the Family Botryosphaeriaceae. Microorganisms 2023, 11, 585. https://doi.org/10.3390/microorganisms11030585
Hernández D, García-Pérez O, Perera S, González-Carracedo MA, Rodríguez-Pérez A, Siverio F. Fungal Pathogens Associated with Aerial Symptoms of Avocado (Persea americana Mill.) in Tenerife (Canary Islands, Spain) Focused on Species of the Family Botryosphaeriaceae. Microorganisms. 2023; 11(3):585. https://doi.org/10.3390/microorganisms11030585
Chicago/Turabian StyleHernández, David, Omar García-Pérez, Santiago Perera, Mario A. González-Carracedo, Ana Rodríguez-Pérez, and Felipe Siverio. 2023. "Fungal Pathogens Associated with Aerial Symptoms of Avocado (Persea americana Mill.) in Tenerife (Canary Islands, Spain) Focused on Species of the Family Botryosphaeriaceae" Microorganisms 11, no. 3: 585. https://doi.org/10.3390/microorganisms11030585