
Dietary Effects of a Short-Term Administration of Microalgae Blend on Growth Performance, Tissue Fatty Acids, and Predominant Intestinal Microbiota in Sparus aurata
 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Microalgae Biomass
2.3. Experimental Feeds and Feeding Trial
2.4. Fish Sampling
2.5. Growth Performance, Nutrient Utilization, and Somatic Indices
2.6. Chemical Composition of Algae and Fish
2.7. Fatty Acid Composition of the Algae, Experimental Diets, and Fish
2.8. Evaluation of the Intestinal Microbiota
2.8.1. DNA Extraction, PCR Amplification, and Analysis of DGGE Patterns
2.8.2. Sequencing of DGGE Bands
2.9. Statistical Analysis
3. Results
3.1. Growth Performance, Nutrient Utilization, and Proximate Composition
3.2. Fatty Acid Composition of Fish
3.3. Evaluation of Intestinal Microbiota
4. Discussion
4.1. Growth Performance, Nutrient Utilization, and Proximate Composition
4.2. Fatty Acid Composition of Fish
4.3. Evaluation of Predominant Intestinal Microbiota
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hunter, M.C.; Smith, R.G.; Schipanski, M.E.; Atwood, L.W.; Mortensen, D.A. Agriculture in 2050: Recalibrating targets for sustainable intensification. BioScience 2017, 67, 386–391. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Kiron, V.; Sørensen, M.; Huntley, M.; Vasanth, G.K.; Gong, Y.; Dahle, D.; Palihawadana, A.M. Defatted biomass of the microalga, Desmodesmus sp., can replace fishmeal in the feeds for Atlantic salmon. Front. Mar. Sci. 2016, 3, 67. [Google Scholar] [CrossRef]
- Sarker, P.K.; Kapuscinski, A.R.; Lanois, A.J.; Livesey, E.D.; Bernhard, K.P.; Coley, M.L. Towards sustainable aquafeeds: Complete substitution of fish oil with marine microalga Schizochytrium sp. improves growth and fatty acid deposition in juvenile Nile tilapia (Oreochromis niloticus). PLoS ONE 2016, 11, e0156684. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Leng, Y.; Lu, Q.; Zhou, W. The application of microalgae biomass and bio-products as aquafeed for aquaculture. Algal Res. 2021, 60, 102541. [Google Scholar] [CrossRef]
- Ravindran, B.; Gupta, S.; Cho, W.-M.; Kim, J.; Lee, S.; Jeong, K.-H.; Lee, D.; Choi, H.-C. Microalgae Potential and Multiple Roles—Current Progress and Future Prospects—An Overview. Sustainability 2016, 8, 1215. [Google Scholar] [CrossRef]
- Shah, M.R.; Lutzu, G.A.; Alam, A.; Sarker, P.; Kabir Chowdhury, M.A.; Parsaeimehr, A.; Liang, Y.; Daroch, M. Microalgae in aquafeeds for a sustainable aquaculture industry. J. Appl. Phycol. 2018, 30, 197–213. [Google Scholar] [CrossRef]
- Tibbetts, S.M. The Potential for ‘Next-Generation’, Microalgae-Based Feed Ingredients for Salmonid Aquaculture in Context of the Blue Revolution. In Microalgal Biotechnology; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Sarker, P.K.; Kapuscinski, A.R.; Bae, A.Y.; Donaldson, E.; Sitek, A.J.; Fitzgerald, D.S.; Edelson, O.F. Towards sustainable aquafeeds: Evaluating substitution of fishmeal with lipid-extracted microalgal co-product (Nannochloropsis oculata) in diets of juvenile Nile tilapia (Oreochromis niloticus). PLoS ONE 2018, 13, e0201315. [Google Scholar] [CrossRef]
- Vizcaíno, A.J.; Rodiles, A.; López, G.; Sáez, M.I.; Herrera, M.; Hachero, I.; Martínez, T.F.; Cerón-García, M.C.; Alarcón, F.J. Growth performance, body composition, and digestive functionality of Senegalese sole (Solea senegalensis Kaup, 1858) juveniles fed diets including microalgae freeze-dried biomass. Fish Physiol. Biochem. 2018, 44, 661–677. [Google Scholar] [CrossRef]
- Perera, E.; Sánchez-Ruiz, D.; Sáez, M.I.; Galafat, A.; Barany, A.; Fernández-Castro, M.; Vizcaíno, A.J.; Fuentes, J.; Martínez, T.F.; Mancera, J.M.; et al. Low dietary inclusion of nutraceuticals from microalgae improves feed efficiency and modifies intermediary metabolisms in gilthead sea bream (Sparus aurata). Sci. Rep. 2020, 10, 18676. [Google Scholar] [CrossRef]
- Oostlander, P.C.; van Houcke, J.; Wijffels, R.H.; Barbosa, M.J. Microalgae production cost in aquaculture hatcheries. Aquaculture 2020, 525, 735310. [Google Scholar] [CrossRef]
- Vizcaíno, A.J.; Saéz, M.I.; López, G.; Arizcun, M.; Abellán, E.; Martínez, T.F.; Cerón-García, M.C.; Alarcón, F.J. Tetraselmis suecia and Tisochrysis lutea meal as dietary ingredients for gilthead sea bream (Sparus aurata L.) fry. J. Appl. Phycol. 2016, 28, 2843–2855. [Google Scholar] [CrossRef]
- García-Márquez, J.; Rico, R.M.; Sánchez-Saavedra, M.P.; Gómez-Pinchetti, J.L.; Acién, F.G.; Figueroa, F.L.; Alarcón, F.J.; Moriñigo, M.Á.; Abdala-Díaz, R.T. A short pulse of dietary algae boosts immune response and modulates fatty acid composition in juvenile Oreochromis niloticus. Aquac. Res. 2020, 51, 4397–4409. [Google Scholar] [CrossRef]
- Vizcaíno, A.J.; López, G.; Sáez, M.I.; Jiménez, J.A.; Barros, A.; Hidalgo, L.; Camacho-Rodríguez, J.; Martínez, T.F.; Cerón-García, M.C.; Alarcón, F.J. Effects of the microalga Scenedesmus almeriensis as fishmeal alternative in diets for gilthead sea bream, Sparus aurata, juveniles. Aquaculture 2014, 431, 34–43. [Google Scholar] [CrossRef]
- Gao, F.; Dominguez-Cabanelas, I.T.; Wijffels, R.H.; Barbosa, M.J. Fucoxanthin and docosahexaenoic acid production by cold-adapted Tisochrysis lutea. New Biotechnol. 2022, 66, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Chua, E.T.; Schenk, P.M. A biorefinery for Nannochloropsis: Induction, harvesting, and extraction of EPA-rich oil and high-value protein. Bioresour. Technol. 2017, 244, 1416–1424. [Google Scholar] [CrossRef]
- Brown, M.R.; Jeffrey, S.W.; Volkman, J.K.; Dunstan, G.A. Nutritional properties of microalgae for mariculture. Aquaculture 1997, 151, 315–331. [Google Scholar] [CrossRef]
- Sánchez, J.F.; Fernández, J.M.; Acién, F.G.; Rueda, A.; Pérez-Parra, J.; Molina, E. Influence of culture conditions on the productivity and lutein content of the new strain Scenedesmus almeriensis. Process Biochem. 2008, 43, 398–405. [Google Scholar] [CrossRef]
- Mann, J.E.; Myers, J. On pigments, growth, and photosynthesis of Phaeodactylum tricornutum. J. Phycol. 1968, 4, 349–355. [Google Scholar] [CrossRef]
- Lourenço, S.O.; Barbarino, E.; De-Paula, J.C.; Pereira, L.O.D.S.; Lanfer Marquez, U.M. Amino acid composition, protein content and calculation of nitrogen-to-protein conversion factors for 19 tropical seaweeds. Phycol. Res. 2002, 50, 233–241. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Christie, W.W. Lipid Analysis, 3rd ed.; Pergamon Press: Oxford, UK, 2003. [Google Scholar]
- Martínez, G.; Shaw, E.M.; Carrillo, M.; Zanuy, S. Protein salting-out method applied to genomic DNA isolation from fish whole blood. BioTechniques 1998, 24, 238–239. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Paniagua, S.T.; Chabrillón, M.; Díaz-Rosales, P.; de la Banda, I.G.; Lobo, C.; Balebona, M.C.; Moriñigo, M.A. Intestinal Microbiota Diversity of the Flat Fish Solea senegalensis (Kaup, 1858) Following Probiotic Administration. Microb. Ecol. 2010, 60, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Konstantinov, S.R.; Zhu, W.-Y.; Williams, B.A.; Tamminga, S.; Vos, W.M.; Akkermans, A.D.L. Effect of fermentable carbohydrates on piglet faecal bacterial communities as revealed by denaturing gradient gel electrophoresis analysis of 16S ribosomal DNA. FEMS Microbiol. Ecol. 2003, 43, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Rico, R.M.; Tejedor-Junco, M.T.; Tapia-Paniagua, S.T.; Alarcón, F.J.; Mancera, J.M.; López-Figueroa, F.; Balebona, M.C.; Abdala-Díaz, R.T.; Moriñigo, M.A. Influence of the dietary inclusion of Gracilaria cornea and Ulva rigida on the biodiversity of the intestinal microbiota of Sparus aurata juveniles. Aquac. Int. 2016, 24, 965–984. [Google Scholar] [CrossRef]
- Sanguinetti, C.J.; Neto, E.D.; Simpson, A.J.G. Rapid silver staining and recovery of PCR products separated on polyacrylamide gels. BioTechniques 1994, 17, 914–921. [Google Scholar] [PubMed]
- Häne, B.G.; Jäger, K.; Drexler, H.G. The Pearson product-moment correlation coefficient is better suited for identification of DNA fingerprint profiles than band matching algorithms. Electrophoresis 1993, 14, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Abdala-Díaz, R.T.; García-Márquez, J.; Rico, R.M.; Gómez-Pinchetti, J.L.; Mancera, J.M.; Figueroa, F.L.; Alarcón, F.J.; Martínez-Manzanares, E.; Moriñigo, M.Á. Effects of a short pulse administration of Ulva rigida on innate immune response and intestinal microbiota in Sparus aurata juveniles. Aquac. Res. 2021, 52, 3038–3051. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef]
- Maidak, B.L.; Cole, J.R.; Parker, C.T.; Garrity, G.M.; Larsen, N.; Li, B.; Lilburn, T.G.; McCaughey, M.J.; Olsen, G.J.; Overbeek, R.; et al. A new version of the RDP (Ribosomal Database Project). Nucleic Acids Res. 1999, 27, 171–173. [Google Scholar] [CrossRef]
- Ludwig, W.; Strunk, O.; Westram, R.; Richter, L.; Meier, H.; Yadhukumar, A.; Buchner, A.; Lai, T.; Steppi, S.; Jacob, G.; et al. ARB: A software environment for sequence data. Nucleic Acids Res. 2004, 32, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Younis, E.S.M.; Al-Quffail, A.S.; Al-Asgah, N.A.; Abdel-Warith, A.W.A.; Al-Hafedh, Y.S. Effect of dietary fish meal replacement by red algae, Gracilaria arcuata, on growth performance and body composition of Nile tilapia Oreochromis niloticus. Saudi J. Biol. Sci. 2018, 25, 198–203. [Google Scholar] [CrossRef] [PubMed]
- González-Félix, M.L.; Maldonado-Othón, C.A.; Perez-Velazquez, M. Effect of dietary lipid level and replacement of fish oil by soybean oil in compound feeds for the shortfin corvina (Cynoscion parvipinnis). Aquaculture 2016, 454, 217–228. [Google Scholar] [CrossRef]
- Kissinger, K.R.; García-Ortega, A.; Trushenski, J.T. Partial fish meal replacement by soy protein concentrate, squid and algal meals in low fish-oil diets containing Schizochytrium limacinum for longfin yellowtail Seriola rivoliana. Aquaculture 2016, 452, 37–44. [Google Scholar] [CrossRef]
- Tibaldi, E.; Chini Zittelli, G.; Parisi, G.; Bruno, M.; Giorgi, G.; Tulli, F.; Venturini, S.; Tredici, M.R.; Poli, B.M. Growth performance and quality traits of European sea bass (D. labrax) fed diets including increasing levels of freeze-dried Isochrysis sp. (T-ISO) biomass as a source of protein and n-3 long chain PUFA in partial substitution of fish derivatives. Aquaculture 2015, 440, 60–68. [Google Scholar] [CrossRef]
- Sørensen, M.; Berge, G.M.; Reitan, K.I.; Ruyter, B. Microalga Phaeodactylum tricornutum in feed for Atlantic salmon (Salmo salar) -Effect on nutrient digestibility, growth and utilization of feed. Aquaculture 2016, 460, 116–123. [Google Scholar] [CrossRef]
- Nakagawa, H. Effect of dietary algae on improvement of lipid metabolism in fish. Biomed. Pharmacother. 1997, 51, 345–348. [Google Scholar] [CrossRef]
- Ergün, S.; Soyutürk, M.; Güroy, B.; Güroy, D.; Merrifield, D. Influence of Ulva meal on growth, feed utilization, and body composition of juvenile Nile tilapia (Oreochromis niloticus) at two levels of dietary lipid. Aquac. Int. 2009, 17, 355–361. [Google Scholar] [CrossRef]
- Ji, H.; Om, A.D.; Yoshimatsu, T.; Hayashi, M.; Umino, T.; Nakagawas, H.; Asano, M.; Nakagawa, A. Effect of dietary vitamins C and E fortification on lipid metabolism in red sea bream Pagrus major and black sea bream Acanthopagrus schlegeli. Fish. Sci. 2003, 69, 1001–1009. [Google Scholar] [CrossRef]
- Love, R.M. The Chemical Biology of Fishes; Academic Press: London, UK, 1980; Volume 2. [Google Scholar]
- Saliu, J.K.; Joy, O.; Catherine, O. Condition factor, fat and protein content of five fish species in Lekki Lagoon, Nigeria. Life Sci. J. 2007, 4, 54–57. [Google Scholar]
- Fernandes, T.J.R.; Alves, R.C.; Souza, T.; Silva, J.M.G.; Castro-Cunha, M.; Valente, L.M.P.; Oliveira, M.B.P.P. Lipid content and fatty acid profile of Senegalese sole (Solea senegalensis Kaup, 1858) juveniles as affected by feed containing different amounts of plant protein sources. Food Chem. 2012, 134, 1337–1342. [Google Scholar] [CrossRef] [PubMed]
- Fountoulaki, E.; Vasilaki, A.; Hurtado, R.; Grigorakis, K.; Karacostas, I.; Nengas, I.; Rigos, G.; Kotzamanis, Y.; Venou, B.; Alexis, M.N. Fish oil substitution by vegetable oils in commercial diets for gilthead sea bream (Sparus aurata L.); effects on growth performance, flesh quality and fillet fatty acid profile: Recovery of fatty acid profiles by a fish oil finishing diet under fluctuating water temperatures. Aquaculture 2009, 289, 317–326. [Google Scholar]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Cardinaletti, G.; Messina, M.; Bruno, M.; Tulli, F.; Poli, B.M.; Giorgi, G.; Chini-Zittelli, G.; Tredici, M.; Tibaldi, E. Effects of graded levels of a blend of Tisochrysis lutea and Tetraselmis suecica dried biomass on growth and muscle tissue composition of European sea bass (Dicentrarchus labrax) fed diets low in fish meal and oil. Aquaculture 2018, 485, 173–182. [Google Scholar] [CrossRef]
- García-Márquez, J.; Galafat, A.; Vizcaíno, A.J.; Barany, A.; Martos-Sitcha, J.A.; Mancera, J.M.; Acién, G.; Figueroa, F.L.; Alarcón, F.J.; Arijo, S.; et al. Dietary Use of the Microalga Chlorella fusca Improves Growth, Metabolism, and Digestive Functionality in Thick-Lipped Grey Mullet (Chelon labrosus, Risso 1827) Juveniles. Front. Mar. Sci. 2022, 9, 902203. [Google Scholar] [CrossRef]
- Emery, J.A.; Norambuena, F.; Trushenski, J.; Turchini, G.M. Uncoupling EPA and DHA in fish nutrition: Dietary demand is limited in Atlantic salmon and effectively met by DHA alone. Lipids 2016, 51, 399–412. [Google Scholar] [CrossRef]
- Bell, J.G.; Henderson, R.J.; Tocher, D.R.; McGhee, F.; Dick, J.R.; Porter, A.; Smullen, R.P.; Sargent, J.R. Substituting fish oil with crude palm oil in the diet of Atlantic salmon (Salmo salar) affects muscle fatty acid composition and hepatic fatty acid metabolism. J. Nutr. 2002, 132, 222–230. [Google Scholar] [CrossRef]
- Tocher, D.R. Metabolism and functions of lipids and fatty acids in teleost fish. Rev. Fish. Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Peng, X.; Li, F.; Lin, S.; Chen, Y. Effects of total replacement of fish oil on growth performance, lipid metabolism and antioxidant capacity in tilapia (Oreochromis niloticus). Aquac. Int. 2016, 24, 145–156. [Google Scholar] [CrossRef]
- Alhazzaa, R.; Bridle, A.R.; Nichols, P.D.; Carter, C.G. Replacing dietary fish oil with Echium oil enriched barramundi with C18 PUFA rather than long-chain PUFA. Aquaculture 2011, 312, 162–171. [Google Scholar] [CrossRef]
- Butt, R.L.; Volkoff, H. Gut microbiota and energy homeostasis in fish. Front. Endocrinol. 2019, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.-B.; Nie, L.; Chen, J. Current understanding on the roles of gut microbiota in fish disease and immunity. Zool. Res. 2019, 40, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Marzorati, M.; Wittebolle, L.; Boon, N.; Daffonchio, D.; Verstraete, W. How to get more out of molecular fingerprints: Practical tools for microbial ecology. Environ. Microbiol. 2008, 10, 1571–1581. [Google Scholar] [CrossRef] [PubMed]
- Wittebolle, L.; Vervaeren, H.; Verstraete, W.; Boon, N. Quantifying community dynamics of nitrifiers in functionally stable reactors. Appl. Environ. Microbiol. 2008, 74, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xi, L.; Liu, H.; Jin, J.; Yang, Y.; Zhu, X.; Han, D.; Xie, S. High replacement of fishmeal by Chlorella meal affects intestinal microbiota and the potential metabolic function in largemouth bass (Micropterus salmoides). Front. Microbiol. 2022, 13, 3772. [Google Scholar] [CrossRef]
- Cerezuela, R.; Guardiola, F.A.; González, P.; Meseguer, J.; Esteban, M.Á. Effects of dietary Bacillus subtilis, Tetraselmis chuii, and Phaeodactylum tricornutum, singularly or in combination, on the immune response and disease resistance of sea bream (Sparus aurata L.). Fish Shellfish Immunol. 2012, 33, 342–349. [Google Scholar] [CrossRef]
- Lyons, P.P.; Turnbull, J.F.; Dawson, K.A.; Crumlish, M. Effects of low-level dietary microalgae supplementation on the distal intestinal microbiome of farmed rainbow trout Oncorhynchus mykiss (Walbaum). Aquac. Res. 2016, 48, 2438–2452. [Google Scholar] [CrossRef]
- Jorge, S.S.; Enes, P.; Serra, C.R.; Castro, C.; Iglesias, P.; Oliva Teles, A.; Couto, A. Short-term supplementation of gilthead seabream (Sparus aurata) diets with Nannochloropsis gaditana modulates intestinal microbiota without affecting intestinal morphology and function. Aquac. Nutr. 2019, 25, 1388–1398. [Google Scholar] [CrossRef]
- Acerete, L.; Espinosa, E.; Josa, A.; Tort, L. Physiological response of hybrid striped bass subjected to Photobacterium damselae subsp. piscicida. Aquaculture 2009, 298, 16–23. [Google Scholar] [CrossRef]
- Tapia-Paniagua, S.T.; Fumanal, M.; Anguís, V.; Fernández-DÍaz, C.; Alarcón, F.J.; Moriñigo, M.A.; Balebona, M.C. Modulation of intestinal microbiota in Solea senegalensis fed low dietary level of Ulva ohnoi. Front. Microbiol. 2019, 10, 171. [Google Scholar] [CrossRef]
- Mohammadian, T.; Nasirpour, M.; Tabandeh, M.R.; Heidary, A.A.; Ghanei-Motlagh, R.; Hosseini, S.S. Administrations of autochthonous probiotics altered juvenile rainbow trout Oncorhynchus mykiss health status, growth performance and resistance to Lactococcus garvieae, an experimental infection. Fish Shellfish Immunol. 2019, 86, 269–279. [Google Scholar] [CrossRef]
- Liu, W.; Ren, P.; He, S.; Xu, L.; Yang, Y.; Gu, Z.; Zhou, Z. Comparison of adhesive gut bacteria composition, immunity, and disease resistance in juvenile hybrid tilapia fed two different Lactobacillus strains. Fish Shellfish Immunol. 2013, 35, 54–62. [Google Scholar] [CrossRef]
- Vendrell, D.; Balcázar, J.L.; de Blas, I.; Ruiz-Zarzuela, I.; Gironés, O.; Múzquiz, J.L. Protection of rainbow trout (Oncorhynchus mykiss) from lactococcosis by probiotic bacteria. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 337–345. [Google Scholar] [CrossRef]
- Ringø, E.; Løvmo, L.; Kristiansen, M.; Bakken, Y.; Salinas, I.; Myklebust, R.; Olsen, R.E.; Mayhew, T.M. Lactic acid bacteria vs. pathogens in the gastrointestinal tract of fish: A review. Aquac. Res. 2010, 41, 451–467. [Google Scholar] [CrossRef]
- Mohammadian, T.; Alishahi, M.; Tabandeh, M.R.; Ghorbanpoor, M.; Gharibi, D. Changes in immunity, expression of some immune-related genes of shabot fish, Tor grypus, following experimental infection with Aeromonas hydrophila: Effects of autochthonous probiotics. Probiotics Antimicrob. Proteins 2018, 10, 616–628. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-C.; Lai, C.-C.; Huang, H.-L.; Su, Y.-T.; Chiu, Y.-H.; Toh, H.-S.; Chiang, S.-R.; Chuang, Y.-C.; Lu, Y.-C.; Tang, H.-J. Antimicrobial ability and mechanism analysis of Lactobacillus species against carbapenemase-producing Enterobacteriaceae. J. Microbiol. Immunol. Infect. 2021, 54, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Richards, G.P.; Watson, M.A.; Needleman, D.S.; Uknalis, J.; Boyd, E.F.; Fay, J.P. Mechanisms for Pseudoalteromonas piscicida-Induced Killing of Vibrios and Other Bacterial Pathogens. Appl. Environ. Microbiol. 2017, 83, e00175-17. [Google Scholar] [CrossRef]
- Tang, B.L.; Yang, J.; Chen, X.L.; Wang, P.; Zhao, H.L.; Su, H.N.; Li, C.Y.; Yu, Y.; Zhong, S.; Wang, L.; et al. A predator-prey interaction between a marine Pseudoalteromonas sp. and Gram-positive bacteria. Nat. Commun. 2020, 11, 285. [Google Scholar] [CrossRef]
- Peng, L.H.; Liang, X.; Xu, J.K.; Dobretsov, S.; Yang, J.L. Monospecific Biofilms of Pseudoalteromonas Promote Larval Settlement and Metamorphosis of Mytilus coruscus. Sci. Rep. 2020, 10, 2577. [Google Scholar] [CrossRef]
- Paździor, E. Shewanella putrefaciens—A new opportunistic pathogen of freshwater fish. J. Vet. Res. 2016, 60, 429–434. [Google Scholar] [CrossRef]
- Paździor, E.; Pękala-Safińska, A.; Wasyl, D. Phenotypic diversity and potential virulence factors of the Shewanella putrefaciens group isolated from freshwater fish. J. Vet. Res. 2019, 63, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Camara-Ruiz, M.; Balebona, M.C.; Moriñigo, M.A.; Esteban, M.A. Probiotic Shewanella putrefaciens (SpPdp11) as a fish health modulator: A review. Microorganisms 2020, 8, 1990. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Chen, S.; Wu, Y.; Ma, Y.; Qiao, H.; Fan, J.; Wu, H. Dietary supplementation with microalgae enhances the zebrafish growth performance by modulating immune status and gut microbiota. Appl. Microbiol. Biotechnol. 2022, 106, 773–788. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Ingredients (g kg−1 DM) | Control | MB-5 | MB-15 | MB-25 |
|---|---|---|---|---|
| Fishmeal (LT-94) | 650 | 615 | 543 | 471 |
| Microalgae blend a | 0 | 50 | 150 | 250 |
| Fish oil | 32 | 30 | 26 | 22 |
| Soybean lecithin b | 10 | 10 | 10 | 10 |
| Maltodextrin b | 245 | 232 | 208 | 184 |
| Cellulose c | 20 | 20 | 20 | 20 |
| Choline chloride c | 8 | 8 | 8 | 8 |
| Vitamins and minerals d | 25 | 25 | 25 | 25 |
| Sodium alginate d | 10 | 10 | 10 | 10 |
| Proximate composition (% dry matter) | ||||
| Crude protein | 45.2 | 44.8 | 44.9 | 45.7 |
| Crude lipid | 12.4 | 12.0 | 12.1 | 11.9 |
| Ash | 10.8 | 10.9 | 11.1 | 11.2 |
| Moisture | 6.7 | 7.2 | 7.1 | 7.5 |
| Fatty Acids | Experimental Diets | Microalgae blend | |||
|---|---|---|---|---|---|
| Control | MB-5 | MB-15 | MB-25 | ||
| Myristic acid (C14:0) | 7.20 | 3.81 | 3.87 | 3.98 | 2.81 |
| Palmitic acid (C16:0) | 35.63 | 30.21 | 28.92 | 24.45 | 13.22 |
| Palmitoleic acid (C16:1) | 3.94 | 7.31 | 7.41 | 7.32 | 6.32 |
| Stearic acid (C18:0) | 9.07 | 5.33 | 4.28 | 3.89 | 3.78 |
| Oleic acid (C18:1n9) | 6.52 | 15.79 | 16.23 | 16.42 | 15.85 |
| Linoleic acid (C18:2n6) | 5.93 | 7.39 | 6.92 | 7.88 | 10.70 |
| α-linolenic acid (C18:3n3) | 0.81 | 1.72 | 1.91 | 2.14 | 5.18 |
| Eicosenoic acid (C20:1) | 0.83 | 1.27 | 0.89 | 1.12 | 1.12 |
| Eicosatrienoic acid (C20:3n3) | 0.40 | 0.37 | 0.41 | 0.49 | 1.31 |
| Arachidonic acid (ARA; C20:4n6) | 0.25 | 0.38 | 0.41 | 0.39 | 2.44 |
| Eicosapentaenoic acid (EPA; C20:5n3) | 8.17 | 9.10 | 9.26 | 10.48 | 12.88 |
| Eurcic acid (C22:1n9) | 0.64 | 1.02 | 1.11 | 2.07 | 4.17 |
| Docosahexaenoic acid (DHA; C22:6n3) | 12.10 | 12.53 | 13.20 | 15.09 | 16.33 |
| Nervonic acid (C24:1) | 0.72 | 0.42 | 0.38 | 0.31 | 0.92 |
| Saturated fatty acid (SFA) | 59.69 | 42.70 | 41.90 | 36.29 | 22.78 |
| Monounsaturated fatty acid (MUFA) | 12.65 | 25.81 | 26.02 | 27.24 | 28.38 |
| Polyunsaturated fatty acid (PUFA) | 20.66 | 24.49 | 25.11 | 29.47 | 48.84 |
| n3 | 21.48 | 23.72 | 24.78 | 28.20 | 35.70 |
| n6 | 6.18 | 7.77 | 7.33 | 8.27 | 13.14 |
| n9 | 7.16 | 16.81 | 17.34 | 18.49 | 20.02 |
| n3/n6 | 3.47 | 3.05 | 3.38 | 3.40 | 2.71 |
| EPA/DHA | 0.67 | 0.72 | 0.69 | 0.69 | 0.78 |
| Parameters | Control | MB-5 | MB-15 | MB-25 | p |
|---|---|---|---|---|---|
| Growth and nutrient utilization | |||||
| Initial mean weight (g) | 15.9 ± 0.2 | 16.6 ± 0.6 | 15.9 ± 0.7 | 16.0 ± 1.3 | n.s. |
| Final mean weight (g) | 28.7 ± 1.3 | 28.5 ± 2.3 | 27.2 ± 0.7 | 26.9 ± 1.6 | n.s. |
| WGR (%) | 75.8 ± 9.1 | 71.7 ± 8.8 | 71.7 ± 6.5 | 68.8 ± 4.2 | n.s. |
| SGR (% day−1) | 2.0 ± 0.2 | 1.9 ± 0.2 | 1.9 ± 0.1 | 1.8 ± 0.1 | n.s. |
| Total feed intake (g fish−1) | 17.9 ± 1.8 | 15.5 ± 2.9 | 14.7 ± 0.9 | 15.3 ± 2.2 | n.s. |
| FCR | 1.4 ± 0.2 | 1.3 ± 0.3 | 1.3 ± 0.2 | 1.4 ± 0.2 | n.s. |
| PER | 1.1 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.2 | 1.2 ± 0.1 | n.s. |
| CY (%) | 86.3 ± 3.8 | 90.9 ± 8.1 | 87.1 ± 1.6 | 89.2 ± 2.5 | n.s. |
| Somatic indices | |||||
| HSI (%) | 1.9 ± 0.1 | 1.8 ± 0.3 | 2.0 ± 0.4 | 1.6 ± 0.3 | n.s. |
| VI (%) | 13.7 ± 3.8 | 9.1 ± 8.1 | 12.9 ± 1.6 | 10.8 ± 2.5 | n.s. |
| K (%) | 1.8 ± 0.1 | 1.9 ± 0.2 | 1.8 ± 0.1 | 1.9 ± 0.1 | n.s. |
| Parameters | Control | MB-5 | MB-15 | MB-25 | p |
|---|---|---|---|---|---|
| Protein | 60.4 ± 0.7 | 59.3 ± 0.4 | 59.9 ± 0.6 | 59.0 ± 1.6 | n.s. |
| Lipid | 19.0 ± 0.4 a | 19.8 ± 1.6 a | 12.5 ± 1.4 c | 15.4 ± 1.7 b, c | 0.023 |
| Ash | 15.3 ± 0.2 | 15.4 ± 0.7 | 15.6 ± 0.4 | 15.1 ± 0.7 | n.s. |
| Fatty Acids | Control | MB-5 | MB-15 | MB-25 | p |
|---|---|---|---|---|---|
| Myristic acid (C14:0) | 3.65 ± 0.11 | 3.58 ± 0.14 | 3.39 ± 0.21 | 3.08 ± 0.24 | n.s. |
| Palmitic acid (C16:0) | 19.04 ± 0.24 | 19.11 ± 0.29 | 19.23 ± 0.92 | 20.94 ± 0.78 | n.s. |
| Stearic acid (C18:0) | 16.63 ± 0.23 a | 16.03 ± 0.29 a | 12.83 ± 0.35 b | 10.81 ± 0.69 c | 0.016 |
| Oleic acid (C18:1n9) | 19.87 ± 0.46 b | 18.91 ± 0.31 c | 22.03 ± 1.23 a | 22.54 ± 1.41 a | 0.032 |
| Linoleic acid (C18:2n6) | 5.88 ± 0.21 | 5.74 ± 0.19 | 5.79 ± 0.96 | 5.73 ± 0.43 | n.s. |
| α-linolenic acid (C18:3n3) | 1.20 ± 0.03 b | 1.83 ± 0.09 a | 0.85 ± 0.12 c | 0.95 ± 0.10 c | 0.011 |
| Eicosanoic acid (C20:0) | 0.91 ± 0.08 a | 0.95 ± 0.05 a | 0.41 ± 0.09 b | 0.39 ± 0.06 b | <0.001 |
| Eicosatrienoic acid (C20:3n6) | 2.36 ± 0.17 a | 2.21 ± 0.16 a | 1.68 ± 0.11 b | 1.59 ± 0.06 b | 0.024 |
| Arachidonic acid (ARA; C20:4n6) | 7.45 ± 0.11 | 7.67 ± 0.09 | 8.14 ± 1.25 | 8.15 ± 0.99 | n.s. |
| Eicosapentaenoic acid (EPA; C20:5n3) | 6.43 ± 0.28 b | 6.35 ± 0.25 b | 6.53 ± 0.11 b | 7.04 ± 0.16 a | 0.036 |
| Behenic acid (C22:0) | 2.10 ± 0.23 a | 1.95 ± 0.18 b | 1.31 ± 0.02 c | 0.39 ± 0.03 d | <0.001 |
| Erucic acid (C22:1n9) | 0.13 ± 0.06 | 0.16 ± 0.08 | 0.25 ± 0.09 | 0.21 ± 0.03 | n.s. |
| Docosahexaenoic acid (DHA; C22:6n3) | 12.11 ± 0.42 b | 12.98 ± 0.33 b | 15.62 ± 0.41 a | 15.75 ± 0.34 a | 0.039 |
| Lignoceric acid (C24:0) | 2.05 ± 0.11 | 2.30 ± 0.08 | 2.11 ± 0.13 | 2.14 ± 0.21 | n.s. |
| Nervonic acid (C24:1) | 0.19 ± 0.07 b | 0.23 ± 0.06 a | 0.25 ± 0.05 a | 0.29 ± 0.04 a | 0.041 |
| Saturated fatty acid (SFA) | 44.38 ± 0.34 a | 43.92 ± 0.38 a | 39.28 ± 1.03 b | 37.75 ± 0.93 b | 0.018 |
| Monounsaturated fatty acid (MUFA) | 20.19 ± 0.55 b | 19.30 ± 0.55 b | 22.53 ± 1.47 a | 23.04 ± 1.11 a | 0.009 |
| Polyunsaturated fatty acid (PUFA) | 35.43 ± 0.65 c | 36.78 ± 0.65 c | 38.19 ± 0.35 b | 39.21 ± 0.28 a | 0.010 |
| n3 | 19.74 ± 0.75 b | 21.16 ± 0.75 a, b | 22.58 ± 1.18 a | 23.74 ± 0.86 a | 0.016 |
| n6 | 15.69 ± 0.43 | 15.62 ± 0.43 | 15.61 ± 1.23 | 15.47 ± 1.13 | n.s. |
| n9 | 20.00 ± 0.50 b | 19.07 ± 0.35 c | 22.28 ± 1.30 a | 22.75 ± 1.41 a | 0.034 |
| n3/n6 | 1.25 ± 0.17 c | 1.35 ± 0.17 b, c | 1.44 ± 0.03 b | 1.53 ± 0.03 a | 0.041 |
| EPA/DHA | 0.53 ± 0.03 a | 0.48 ± 0.01 b | 0.41 ± 0.06 c | 0.44 ± 0.02 c | 0.044 |
| Fatty Acids | Control | MB-5 | MB-15 | MB-25 | p |
|---|---|---|---|---|---|
| Myristic acid (C14:0) | 4.50 ± 0.06 a | 4.11 ± 0.17 b | 1.98 ± 0.17 c | 1.90 ± 0.14 c | <0.001 |
| Palmitic acid (C16:0) | 16.83 ± 0.40 a | 16.62 ± 0.26 a | 16.41 ± 0.39 a | 15.39 ± 0.31 b | 0.035 |
| Stearic acid (C18:0) | 4.43 ± 0.19 c | 4.63 ± 0.25 c | 6.42 ± 0.10 a | 5.56 ± 0.16 b | 0.008 |
| Oleic acid (C18:1n9) | 19.83 ± 0.14 c | 19.69 ± 0.17 c | 22.58 ± 0.19 b | 23.45 ± 0.18 a | 0.034 |
| Linoleic acid (C18:2n6) | 4.92 ± 0.09 b | 5.30 ± 0.09 a | 3.09 ± 0.22 c | 3.00 ± 0.22 c | 0.016 |
| α-linolenic acid (C18:3n3) | 1.36 ± 0.09 a | 1.37 ± 0.05 a | 0.99 ± 0.13 b | 0.96 ± 0.12 b | 0.021 |
| Eicosanoic acid (C20:0) | 1.28 ± 0.07 b | 1.43 ± 0.08 a | 0.48 ± 0.06 c | 0.46 ± 0.04 c | 0.017 |
| Gondoic acid (C20:1n9) | 3.86 ± 0.66 a | 4.48 ± 0.72 a | 0.82 ± 0.19 b | 0.88 ± 0.19 b | <0.001 |
| Eicosatrienoic acid (C20:3n6) | 0.48 ± 0.06 b | 0.36 ± 0.06 c | 0.61 ± 0.04 a | 0.57 ± 0.05 a, b | 0.031 |
| Arachidonic acid (ARA; C20:4n6) | 8.63 ± 0.11 b | 8.29 ± 0.12 c | 11.95 ± 0.79 a | 11.82 ± 0.82 a | 0.005 |
| Eicosapentaenoic acid (EPA; C20:5n3) | 7.68 ± 0.22 | 7.29 ± 0.16 | 7.01 ± 0.51 | 7.56 ± 0.30 | n.s. |
| Behenic acid (C22:0) | 3.98 ± 0.52 a | 3.54 ± 0.60 a | 2.24 ± 0.63 b | 1.60 ± 0.06 c | 0.004 |
| Erucic acid (C22:1n9) | 0.24 ± 0.09 b | 0.28 ± 0.06 b | 0.63 ± 0.20 a | 0.40 ± 0.08 a | 0.031 |
| Docosahexaenoic acid (DHA; C22:6n3) | 19.19 ± 0.99 b | 19.08 ± 0.89 b | 21.57 ± 0.97 a | 23.01 ± 1.29 a | 0.028 |
| Lignoceric acid (C24:0) | 2.23 ± 0.09 b | 2.25 ± 0.08 b | 2.86 ± 0.12 a | 2.96 ± 0.13 a | 0.045 |
| Nervonic acid (C24:1) | 0.56 ± 0.32 a, b | 0.37 ± 0.11 b | 0.69 ± 0.16 a | 0.48 ± 0.21 b | 0.034 |
| Saturated fatty acid (SFA) | 33.25 ± 1.01 a | 32.58 ± 0.36 a | 30.39 ± 0.74 b | 27.87 ± 0.53 c | 0.022 |
| Monounsaturated fatty acid (MUFA) | 24.49 ± 1.11 | 24.82 ± 0.83 | 24.39 ± 1.11 | 25.21 ± 1.05 | n.s. |
| Polyunsaturated fatty acid (PUFA) | 42.26 ± 1.10 b, c | 41.69 ± 0.97 c | 44.51 ± 1.62 a, b | 46.92 ± 1.44 a | 0.033 |
| n3 | 28.23 ± 1.39 b | 27.74 ± 1.10 b | 28.86 ± 1.06 b | 31.53 ± 1.08 a | 0.039 |
| n6 | 14.03 ± 0.25 b | 13.95 ± 0.08 b | 15.65 ± 0.71 a | 15.39 ± 0.74 a | 0.037 |
| n9 | 23.93 ± 1.02 | 24.45 ± 0.94 | 24.03 ± 0.53 | 24.73 ± 0.42 | n.s. |
| n3/n6 | 2.01 ± 0.21 | 1.98 ± 0.14 | 1.84 ± 0.16 | 2.04 ± 0.08 | n.s. |
| EPA/DHA | 0.40 ± 0.01 | 0.38 ± 0.01 | 0.32 ± 0.09 | 0.32 ± 0.07 | n.s. |
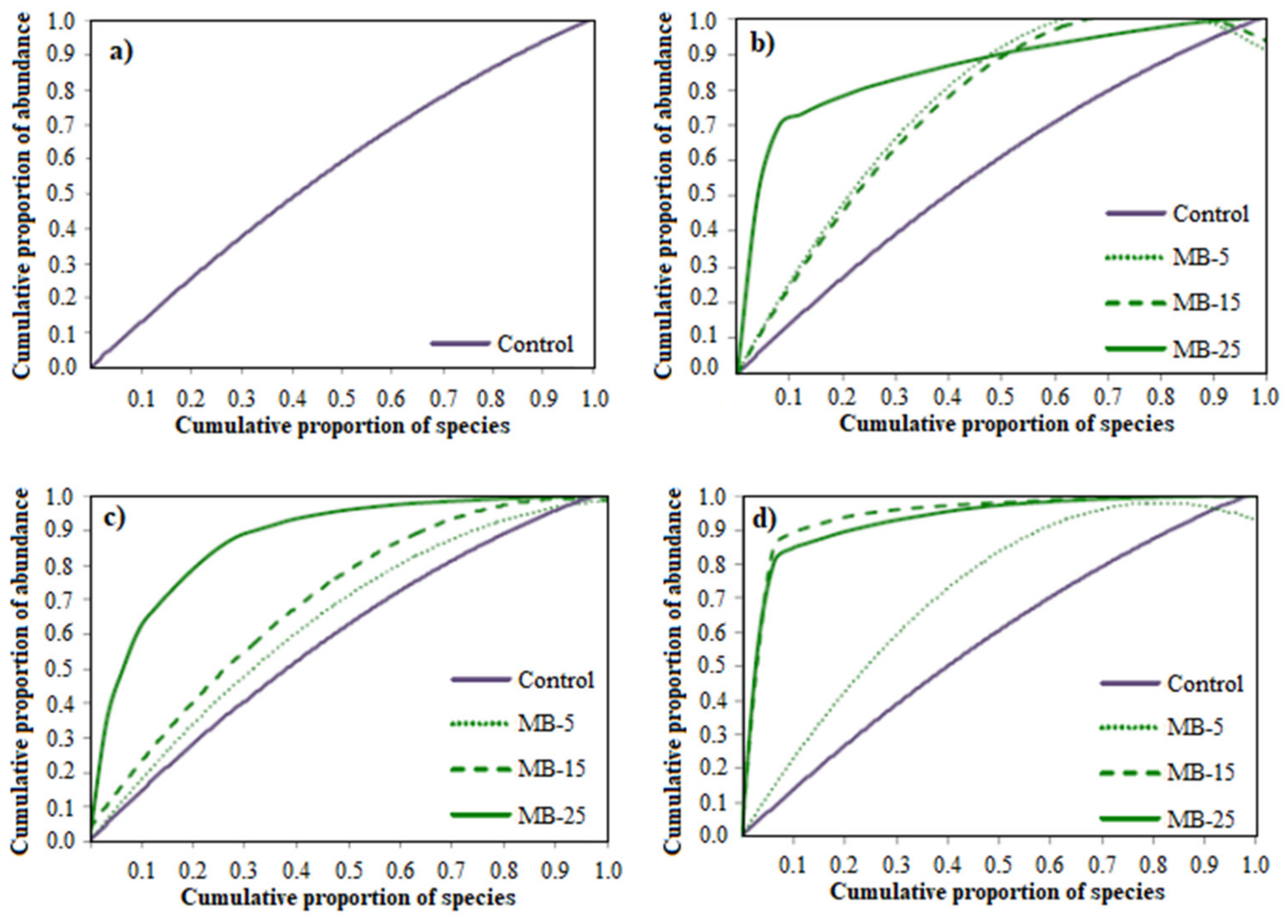
| Day | Diet | R | Gini Coefficient | Rr |
|---|---|---|---|---|
| 0 | Control | 13.25 ± 1.26 b | 0.27 ± 0.01 c | 10.44 ± 3.15 a, b |
| 7 | Control | 13.25 ± 2.63 b | 0.23 ± 0.08 b | 7.97 ± 2.65 a |
| MB-5 | 12.66 ± 3.79 b | 0.21 ± 0.03 b | 20.16 ± 2.84 c | |
| MB-15 | 11.50 ± 4.04 a, b | 0.18 ± 0.03 b | 22.28 ± 10.18 b, c | |
| MB-25 | 23.33 ± 4.04 d | 0.25 ± 0.09 b | 136.29 ± 10.74 f | |
| 15 | Control | 15.50 ± 1.29 b, c | 0.23 ± 0.05 b | 24.72 ± 6.01 c |
| MB-5 | 11.25 ± 3.86 a, b | 0.25 ± 0.02 b, c | 51.22 ± 4.99 d | |
| MB-15 | 7.75 ± 2.22 a | 0.18 ± 0.05 b | 14.96 ± 5.88 b, c | |
| MB-25 | 25.00 ± 5.57 d | 0.25 ± 0.05 b, c | 292.33 ± 15.03 h | |
| 30 | Control | 13.00 ± 1.00 b | 0.23 ± 0.08 b | 14.56 ± 3.83 b, c |
| MB-5 | 17.75 ± 2.63 c | 0.27 ± 0.03 c | 87.79 ± 5.41 e | |
| MB-15 | 27.25 ± 3.59 d | 0.11 ± 0.04 a | 161.67 ± 5.10 g | |
| MB-25 | 36.50 ± 6.40 e | 0.25 ± 0.05 b, c | 374.00 ± 18.47 i | |
| p | <0.001 | <0.001 | <0.001 |
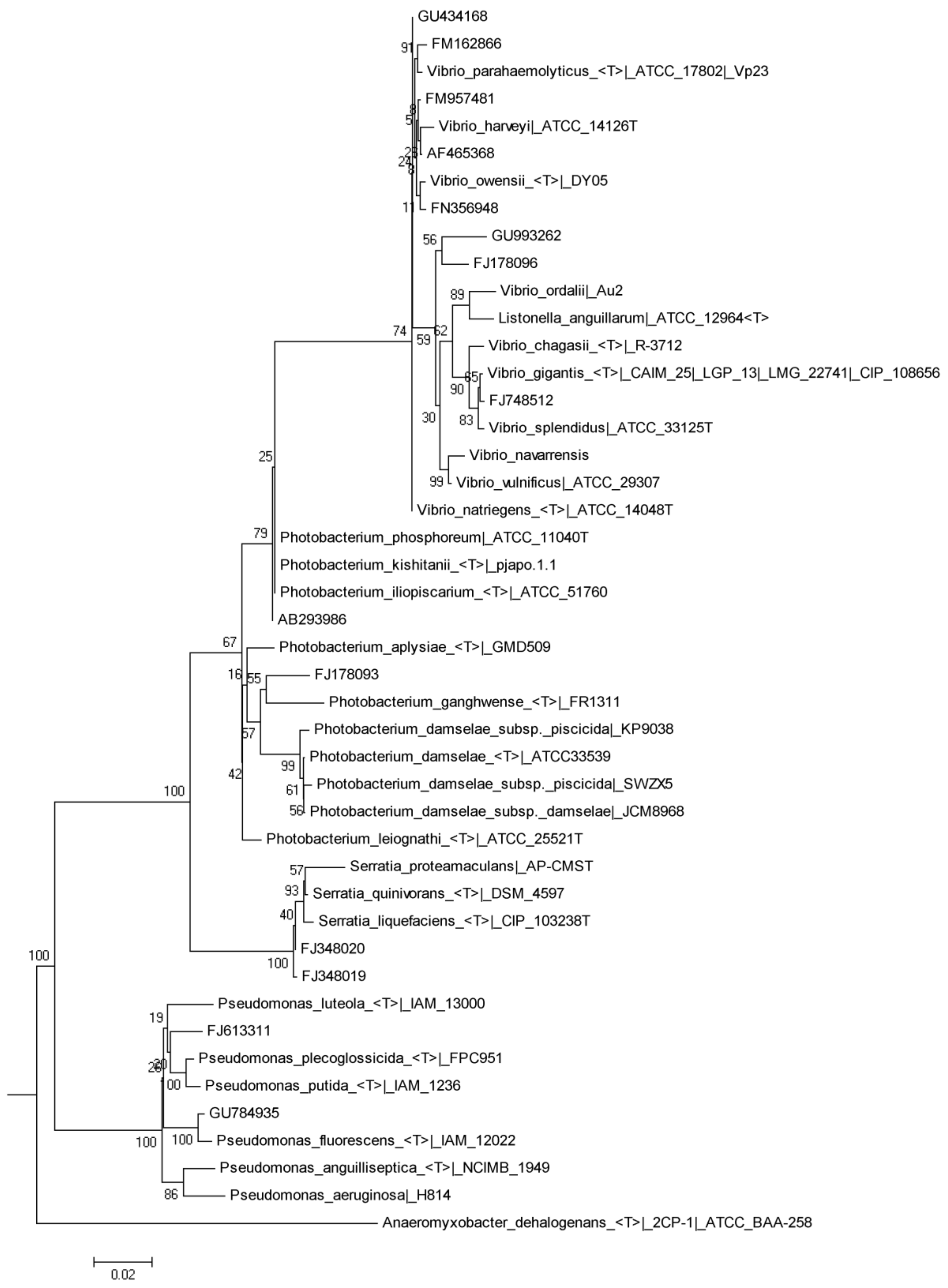
| Control | MB-5 | MB-15 | MB-25 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Closest Relative | Similarity (%) | Genbank Accession Number | Taxon | 0 Days | 7 Days | 15 Days | 30 Days | 7 Days | 15 Days | 30 Days | 7 Days | 15 Days | 30 Days | 7 Days | 15 Days | 30 Days |
| Agarivorans sp. QM34 | 100 | GQ426318 | γ-proteobacteria | X | X | X | X | X | X | X | X | X | X | X | X | |
| Comamonas aquatica strain 530 | 100 | EU841527 | β-proteobacteria | X | X | |||||||||||
| Enterobacteriaceae bacterium | 99 | FJ348019 | γ-Proteobacteria | X | X | X | X | |||||||||
| Lactobacillus delbrueckii subsp. bulgaricus | 100 | FJ915706 | Firmicutes | X | X | X | ||||||||||
| Lactococcus sp. | 98 | FR873792 | Firmicutes | X | X | X | ||||||||||
| Photobacterium sp. | 99 | FJ178093 | γ-proteobacteria | X | X | X | X | X | X | |||||||
| Pseudoalteromonas sp. | 98 | HM475290 | γ-proteobacteria | X | X | |||||||||||
| Pseudoalteromonas sp. DFH4.24 | 98 | FR873779 | γ-proteobacteria | X | ||||||||||||
| Pseudomonas sp. | 98 | GU784935 | γ-proteobacteria | X | X | X | X | |||||||||
| Shewanella sp. | 100 | AY515438 | γ-proteobacteria | X | X | X | X | X | X | X | X | |||||
| Sphingomonas sp. | 98 | GU300600 | α-Proteobacteria | X | ||||||||||||
| Thalassomonas haliotis | 99 | AB369381 | γ-proteobacteria | X | X | |||||||||||
| Uncultured cyanobacterium clone O7 | 99 | FJ178040 | Cyanobacteria | X | X | X | X | X | X | |||||||
| Uncultured Firmicutes | 100 | FM225297 | Firmicutes | X | X | X | X | X | ||||||||
| Vibrio alginolyticus | 100 | DQ005214 | γ-proteobacteria | X | X | X | X | X | X | |||||||
| Vibrio alginolyticus | 99 | EU155510 | γ-proteobacteria | X | X | X | X | X | X | X | ||||||
| Vibrio alginolyticus strain UST981101-031 | 99 | EU833999 | γ-proteobacteria | X | ||||||||||||
| Vibrio communis strain R-40504 | 99 | GU078675 | γ-proteobacteria | X | ||||||||||||
| Vibrio harveyi strain BK2 | 100 | HM355956 | γ-proteobacteria | X | X | X | X | X | X | |||||||
| Vibrio ichthyoenteri | 99 | EF635304 | γ-proteobacteria | X | ||||||||||||
| Vibrio pathogenic sp. | 99 | AB274764 | γ-proteobacteria | X | ||||||||||||
| Vibrio ordalii | 97 | AF493811 | γ-proteobacteria | X | X | |||||||||||
| Vibrio parahaemolyticus | 97 | EU660313 | γ-proteobacteria | X | X | X | ||||||||||
| Vibrio sp. | 100 | GU434168 | γ-proteobacteria | X | X | X | X | X | ||||||||
| Vibrio vulnificus | 100 | AY245180 | γ-proteobacteria | X | ||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Márquez, J.; Rico, R.M.; Acién, F.G.; Mancera, J.M.; Figueroa, F.L.; Vizcaíno, A.J.; Alarcón, F.J.; Moriñigo, M.Á.; Abdala-Díaz, R.T. Dietary Effects of a Short-Term Administration of Microalgae Blend on Growth Performance, Tissue Fatty Acids, and Predominant Intestinal Microbiota in Sparus aurata. Microorganisms 2023, 11, 463. https://doi.org/10.3390/microorganisms11020463
García-Márquez J, Rico RM, Acién FG, Mancera JM, Figueroa FL, Vizcaíno AJ, Alarcón FJ, Moriñigo MÁ, Abdala-Díaz RT. Dietary Effects of a Short-Term Administration of Microalgae Blend on Growth Performance, Tissue Fatty Acids, and Predominant Intestinal Microbiota in Sparus aurata. Microorganisms. 2023; 11(2):463. https://doi.org/10.3390/microorganisms11020463
Chicago/Turabian StyleGarcía-Márquez, Jorge, Rosa María Rico, Francisco Gabriel Acién, Juan Miguel Mancera, Félix L. Figueroa, Antonio Jesús Vizcaíno, Francisco Javier Alarcón, Miguel Ángel Moriñigo, and Roberto Teófilo Abdala-Díaz. 2023. "Dietary Effects of a Short-Term Administration of Microalgae Blend on Growth Performance, Tissue Fatty Acids, and Predominant Intestinal Microbiota in Sparus aurata" Microorganisms 11, no. 2: 463. https://doi.org/10.3390/microorganisms11020463