


Impact of Pipe Material and Temperature on Drinking Water Microbiome and Prevalence of Legionella, Mycobacterium, and Pseudomonas Species
Abstract
:1. Introduction
2. Materials and Methods
2.1. System Design and Sample Collection Procedure
2.2. Chemical Analysis of Water Samples
2.3. Sample Processing and DNA Extraction
2.4. 16S Illumina Sequencing
2.5. Microbiome Statistical Analysis
3. Results
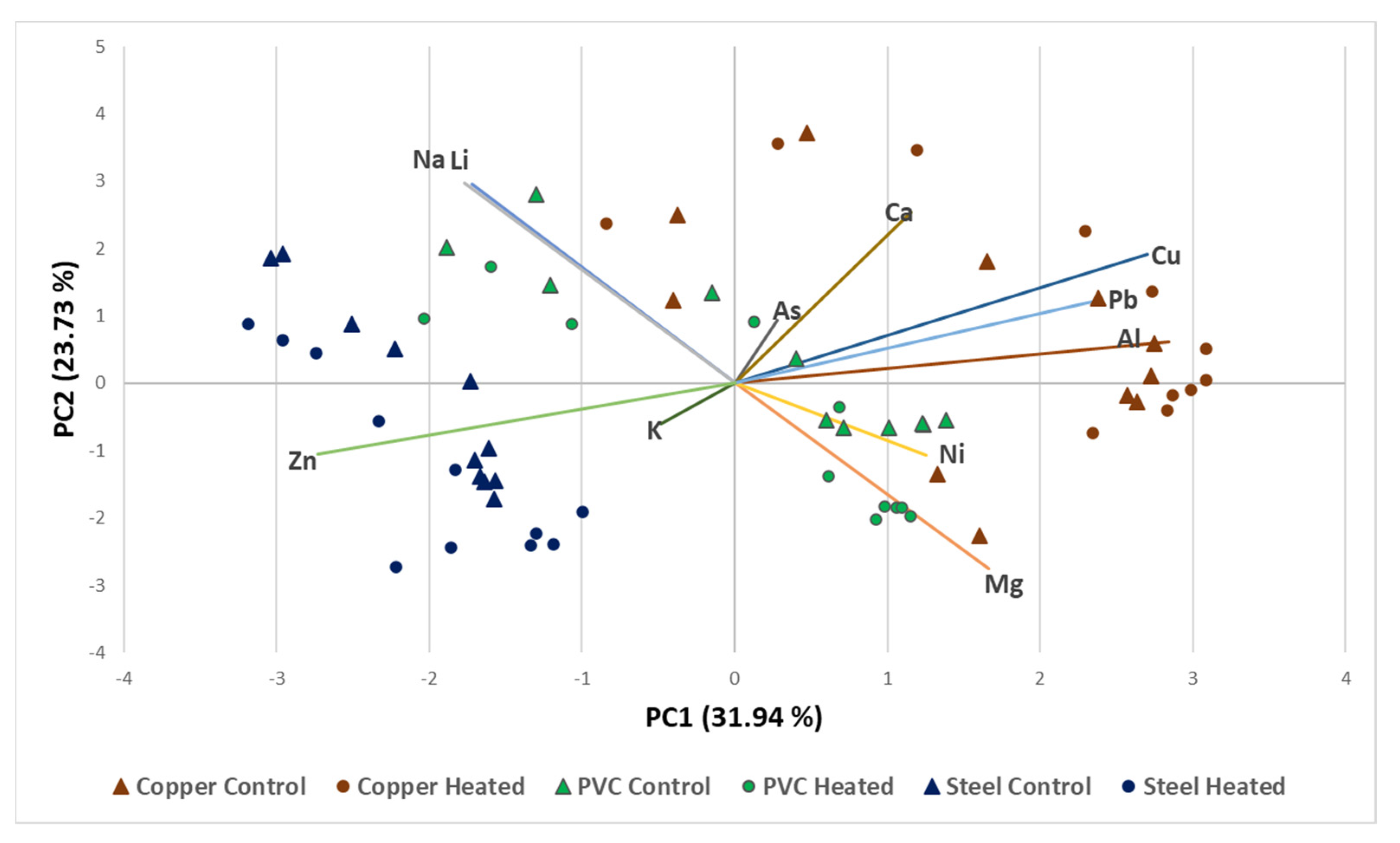
3.1. Characterization of Chemical Variables in the Model Distribution Systems

3.2. Bacterial Diversity of the Water Samples in the Model Water Distribution Systems

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature Condition | Warm | Cold | ||||
|---|---|---|---|---|---|---|
| Material Type | Copper | PEX | Steel | Copper | PEX | Steel |
| Legionellaceae | 100% | 100% | 100% | 100% | 100% | 100% |
| Pseudomonadaceae | 20% | 50% | 90% | 36.4% | 63.6% | 88.9% |
| Mycobacteriaceae | 100% | 100% | 100% | 81.8% | 27.3% | 66.7% |

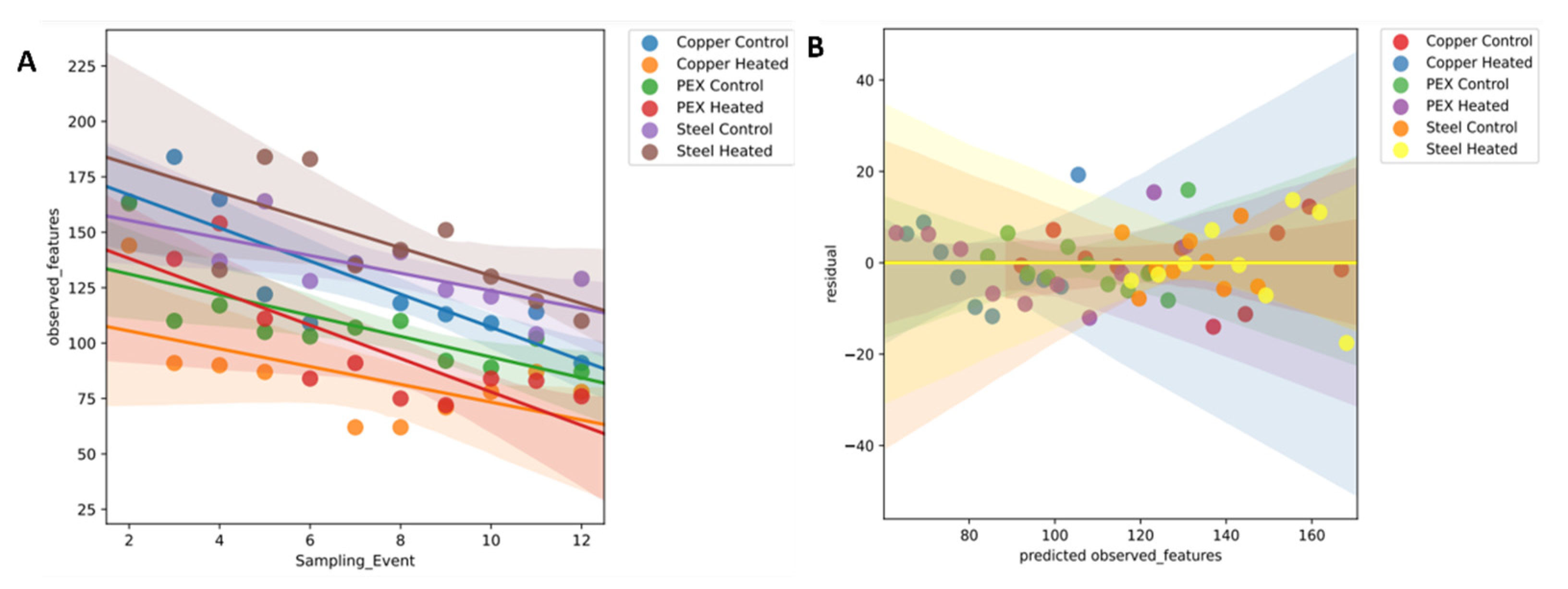
3.3. Species Richness and Evenness for Different Distribution Systems—Alpha Diversity


3.4. Comparison of the Microbial Community Composition-Beta Diversity
| Group 1 | Group 2 | Sample Size | Permutations | Pseudo-F | p-Value | q-Value |
|---|---|---|---|---|---|---|
| Copper 22 °C | Copper 32 °C | 19 | 999 | 9.696 | 0.001 | 0.0012 |
| PEX 22 °C | 20 | 999 | 4.458 | 0.001 | 0.0012 | |
| PEX 32 °C | 19 | 999 | 8.536 | 0.001 | 0.0012 | |
| Steel 22 °C | 20 | 999 | 5.699 | 0.001 | 0.0012 | |
| Steel 32 °C | 18 | 999 | 5.362 | 0.001 | 0.0012 | |
| Copper 32 °C | PEX 22 °C | 21 | 999 | 7.065 | 0.001 | 0.0012 |
| PEX 32 °C | 20 | 999 | 8.201 | 0.001 | 0.0012 | |
| Steel 22 °C | 21 | 999 | 10.648 | 0.001 | 0.0012 | |
| Steel 32 °C | 19 | 999 | 10.865 | 0.001 | 0.0012 | |
| PEX 22 °C | PEX 32 °C | 21 | 999 | 10.928 | 0.001 | 0.0012 |
| Steel 22 °C | 22 | 999 | 6.716 | 0.001 | 0.0012 | |
| Steel 32 °C | 20 | 999 | 7.977 | 0.001 | 0.0012 | |
| PEX 32 °C | Steel 22 °C | 21 | 999 | 10.651 | 0.001 | 0.0012 |
| Steel 32 °C | 19 | 999 | 7.191 | 0.002 | 0.0020 | |
| Steel 22 °C | Steel 32 °C | 20 | 999 | 5.448 | 0.002 | 0.0020 |

3.5. Prediction of Metagenome Functional Profiles
3.6. Differential Abundance of 32 °C and 22 °C Systems Using ALDEx2
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hull, N.M.; Ling, F.; Pinto, A.J.; Albertsen, M.; Jang, H.G.; Hong, P.Y.; Konstantinidis, K.T.; LeChevallier, M.; Colwell, R.R.; Liu, W.T. Drinking Water Microbiome Project: Is it Time? Trends. Microbiol. 2019, 27, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Frederik, H.; Berney, M.; Wang, Y.; Vital, M.; Köster, O.; Egli, T. Flow-Cytometric Total Bacterial Cell Counts as a Descriptive Microbiological Parameter for Drinking Water Treatment Processes. Water Res. 2008, 42, 269–277. [Google Scholar]
- Wingender, J.; Flemming, H.-C. Biofilms in drinking water and their role as reservoir for pathogens. Int. J. Hyg. Environ. Health 2011, 214, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, Z.; Yu, J.; Cao, N.; Liu, R.; Yang, M. Characterization of Bacterial Community Structure in a Drinking Water Distribution System during an Occurrence of Red Water. Appl. Environ. Microbiol. 2010, 76, 7171–7180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, P.; Parks, J.; Edwards, M.A.; Pruden, A. Impact of Water Chemistry, Pipe Material and Stagnation on the Building Plumbing Microbiome. PLoS ONE 2015, 10, e0141087. [Google Scholar] [CrossRef] [Green Version]
- van der Kooij, D.; van der Wielen, P.W. Microbial Growth in Drinking-Water Supplies: Problems, Causes, Control and Research Needs. 2014. Available online: https://books.google.com/books/about/Microbial_Growth_in_Drinking_water_Suppl.html?hl=&id=tDX0jgEACAAJ (accessed on 13 February 2022).
- Wang, H.; Edwards, M.; Falkinham, J.O., 3rd; Pruden, A. Molecular survey of the occurrence of Legionella spp., Mycobacterium spp., Pseudomonas aeruginosa, and amoeba hosts in two chloraminated drinking water distribution systems. Appl. Environ. Microbiol. 2012, 78, 6285–6294. [Google Scholar] [CrossRef] [Green Version]
- Mathys, W.; Stanke, J.; Harmuth, M.; Junge-Mathys, E. Occurrence of Legionella in hot water systems of single-family residences in suburbs of two German cities with special reference to solar and district heating. Int. J. Hyg. Environ. Health 2008, 211, 179–185. [Google Scholar] [CrossRef]
- Bargellini, A.; Marchesi, I.; Righi, E.; Ferrari, A.; Cencetti, S.; Borella, P.; Rovesti, S. Parameters predictive of Legionella contamination in hot water systems: Association with trace elements and heterotrophic plate counts. Water Res. 2011, 45, 2315–2321. [Google Scholar] [CrossRef]
- Falkinham, J.O., 3rd; Norton, C.D.; LeChevallier, M.W. Factors influencing numbers of Mycobacterium avium, Mycobacterium intracellulare, and other Mycobacteria in drinking water distribution systems. Appl. Environ. Microbiol. 2001, 67, 1225–1231. [Google Scholar] [CrossRef] [Green Version]
- Lehtola, M.J.; Miettinen, I.T.; Keinänen, M.M.; Kekki, T.K.; Laine, O.; Hirvonen, A.; Vartiainen, T.; Martikainen, P.J. Microbiology, Chemistry and Biofilm Development in a Pilot Drinking Water Distribution System with Copper and Plastic Pipes. Water Res. 2004, 38, 3769–3779. [Google Scholar] [CrossRef]
- Inkinen, J.; Kaunisto, T.; Pursiainen, A.; Miettinen, I.T.; Kusnetsov, J.; Riihinen, K.; Keinänen-Toivola, M.M. Drinking Water Quality and Formation of Biofilms in an Office Building during Its First Year of Operation, a Full Scale Study. Water Res. 2014, 49, 83–91. [Google Scholar] [CrossRef] [PubMed]
- USEPA. Method 200.7 Trace Elements in Water, Solids, and Biosolids by Inductively Coupled Plasma-Atomic Emission Spectrometry; USEPA: Cincinnati, OH, USA, 2001. [Google Scholar]
- Addinsoft, P. XLSTAT 2016: Data Analysis and Statistical Solution for Microsoft Excel; Addinsoft SARL: Paris, France, 2016. [Google Scholar]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods. 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, T.K.; Ahn, C.W.; Verma, O.P.; Panigrahi, B.K. Soft Computing: Theories and Applications: Proceedings of SoCTA 2020, Volume 2. Springer Nature. 2021. Available online: https://play.google.com/store/books/details?id=jzQ1EAAAQBAJ (accessed on 10 October 2022).
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2--approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). Wiley StatsRef Stat. Ref. Online 2017, 1–15. [Google Scholar] [CrossRef]
- Fernandes, A.D.; Macklaim, J.M.; Linn, T.G.; Reid, G.; Gloor, G.B. ANOVA-like differential expression (ALDEx) analysis for mixed population RNA-Seq. PLoS ONE 2013, 8, e67019. [Google Scholar] [CrossRef]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic. Acids. Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Dillon, M.R.; Zhang, Y.; Rideout, J.R.; Bolyen, E.; Li, H.; Albert, P.S.; Caporaso, J.G. q2-longitudinal: Longitudinal and Paired-Sample Analyses of Microbiome Data. mSystems 2018, 3, e00219-18. [Google Scholar] [CrossRef] [PubMed]
- Seabold, S.; Perktold, J. Statsmodels: Econometric and Statistical Modeling with Python. In Proceedings of the Python in Science Conference, Austin, TX, USA, 28 June–3 July 2010. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martino, C.; Morton, J.T.; Marotz, C.A.; Thompson, L.R.; Tripathi, A.; Knight, R.; Zengler, K. A Novel Sparse Compositional Technique Reveals Microbial Perturbations. mSystems 2019, 4, e00016-19. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Yu, Z.; Chen, X.; Liu, R.; Zhang, H. Molecular characterization of natural biofilms from household taps with different materials: PVC, stainless steel, and cast iron in drinking water distribution system. Appl. Microbiol. Biotechnol. 2013, 97, 8393–8401. [Google Scholar] [CrossRef] [PubMed]
- Henne, K.; Kahlisch, L.; Brettar, I.; Höfle, M.G. Analysis of structure and composition of bacterial core communities in mature drinking water biofilms and bulk water of a citywide network in Germany. Appl. Environ. Microbiol. 2012, 78, 3530–3538. [Google Scholar] [CrossRef] [Green Version]
- Douterelo, I.; Husband, S.; Boxall, J.B. The bacteriological composition of biomass recovered by flushing an operational drinking water distribution system. Water Res. 2014, 54, 100–114. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Edwards, M.A.; Falkinham, J.O., 3rd; Pruden, A. Probiotic approach to pathogen control in premise plumbing systems? A review. Environ. Sci. Technol. 2013, 47, 10117–10128. [Google Scholar] [CrossRef]
- Zeng, Y.; Charkowski, A.O. The Role of ATP-Binding Cassette Transporters in Bacterial Phytopathogenesis. Phytopathology 2021, 111, 600–610. [Google Scholar] [CrossRef]
- Luesken, F.A.; van Alen, T.A.; van der Biezen, E.; Frijters, C.; Toonen, G.; Kampman, C.; Hendrickx, T.L.; Zeeman, G.; Temmink, H.; Strous, M.; et al. Diversity and enrichment of nitrite-dependent anaerobic methane oxidizing bacteria from wastewater sludge. Appl. Microbiol. Biotechnol. 2011, 92, 845–854. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Feng, F.; Medová, H.; Dean, J.; Koblížek, M. Functional type 2 photosynthetic reaction centers found in the rare bacterial phylum Gemmatimonadetes. Proc. Natl. Acad. Sci. USA 2014, 111, 7795–7800. [Google Scholar] [CrossRef]
- Redeker, K.R.; Cai, L.L.; Dumbrell, A.J.; Bardill, A.; Chong, J.P.J.; Helgason, T. Chapter Four—Noninvasive Analysis of the Soil Microbiome: Biomonitoring Strategies Using the Volatilome, Community Analysis, and Environmental Data. In Advances in Ecological Research; Bohan, D.A., Dumbrell, A.J., Woodward, G., Jackson, M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 93–132. [Google Scholar] [CrossRef]
- Lu, J.; Buse, H.Y.; Gomez-Alvarez, V.; Struewing, I.; Santo Domingo, J.; Ashbolt, N.J. Impact of drinking water conditions and copper materials on downstream biofilm microbial communities and Legionella pneumophila colonization. J. Appl. Microbiol. 2014, 117, 905–918. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aloraini, S.; Alum, A.; Abbaszadegan, M. Impact of Pipe Material and Temperature on Drinking Water Microbiome and Prevalence of Legionella, Mycobacterium, and Pseudomonas Species. Microorganisms 2023, 11, 352. https://doi.org/10.3390/microorganisms11020352
Aloraini S, Alum A, Abbaszadegan M. Impact of Pipe Material and Temperature on Drinking Water Microbiome and Prevalence of Legionella, Mycobacterium, and Pseudomonas Species. Microorganisms. 2023; 11(2):352. https://doi.org/10.3390/microorganisms11020352
Chicago/Turabian StyleAloraini, Saleh, Absar Alum, and Morteza Abbaszadegan. 2023. "Impact of Pipe Material and Temperature on Drinking Water Microbiome and Prevalence of Legionella, Mycobacterium, and Pseudomonas Species" Microorganisms 11, no. 2: 352. https://doi.org/10.3390/microorganisms11020352