
Polyphasic Characterization of Geotalea uranireducens NIT-SL11 Newly Isolated from a Complex of Sewage Sludge and Microbially Reduced Graphene Oxide

Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Growth Conditions of NIT-SL11
2.2. Morphological, Physiological, and Biochemical Analyses
2.3. Chemotaxonomic Analysis
2.4. Genetic Characterization
3. Results and Discussion
3.1. Isolation of NIT-SL11
3.2. Phylogenetic Identification Based on 16S rRNA Sequencing
3.3. Physiological and Biochemical Characterization
3.4. Chemotaxonomic Characterization
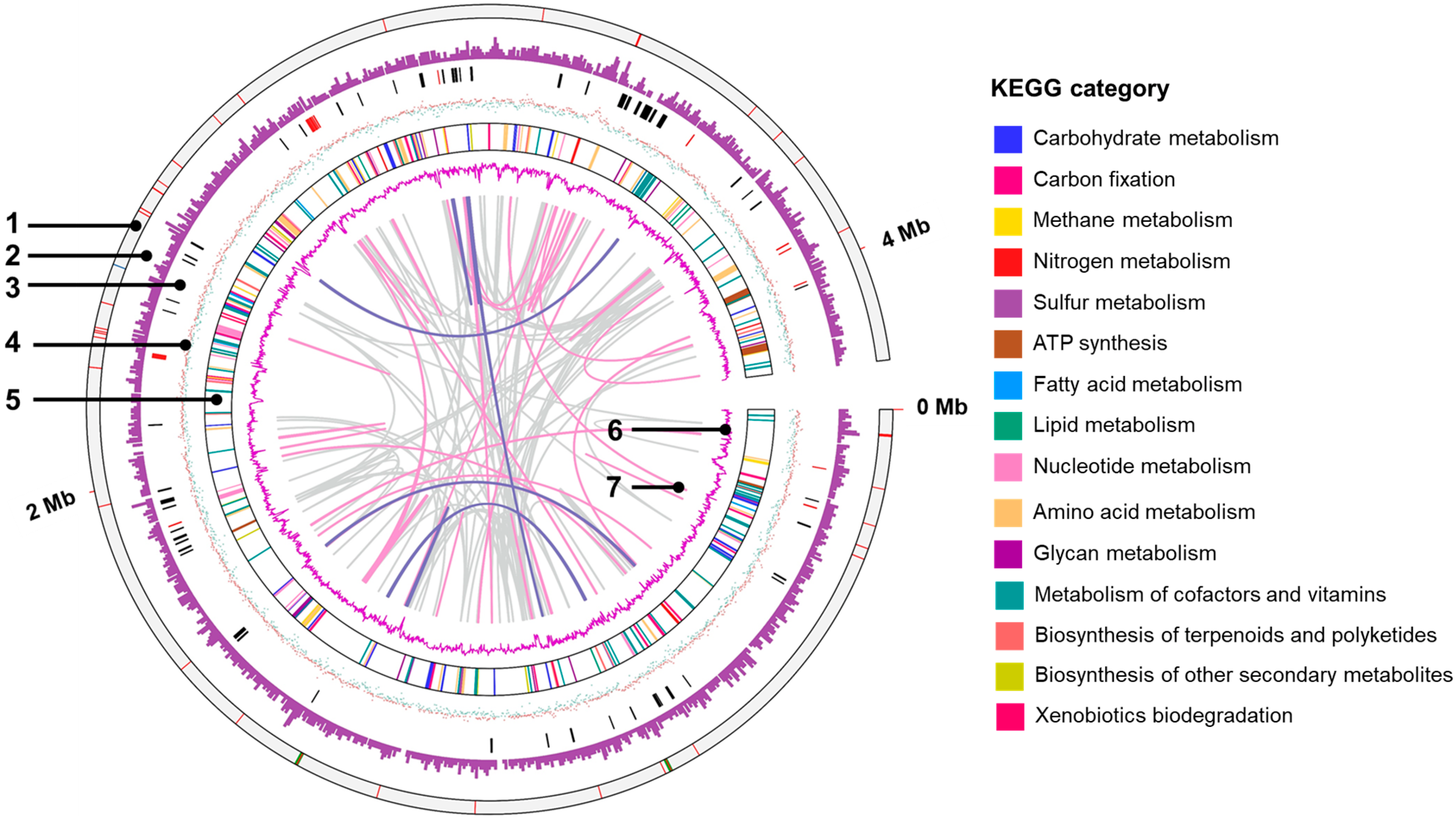
3.5. General Genomic Features
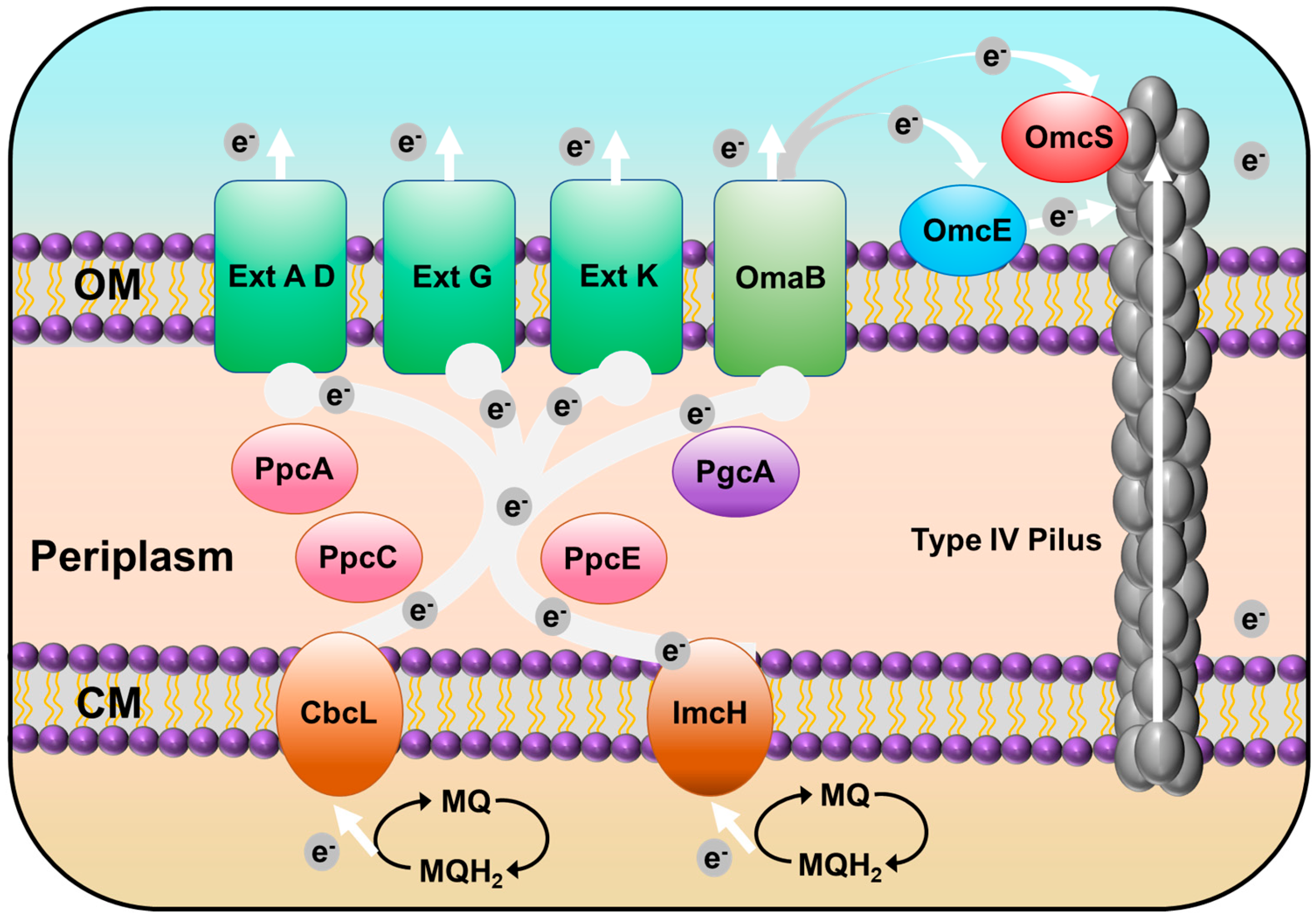
3.6. Putative c-Type Cytochromes
3.7. Type IV Pilus (T4P)-Related Genes
3.8. Exoelectrogens That form the rGO Complex
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Logan, B.E.; Rabaey, K. Conversion of Wastes into Bioelectricity and Chemicals by Using Microbial Electrochemical. Science 2012, 337, 686–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugioka, M.; Yoshida, N.; Yamane, T.; Kakihana, Y.; Higa, M.; Matsumura, T.; Sakoda, M.; Iida, K. Long-Term Evaluation of an Air-Cathode Microbial Fuel Cell with an Anion Exchange Membrane in a 226L Wastewater Treatment Reactor. Environ. Res. 2022, 205, 112416. [Google Scholar] [CrossRef] [PubMed]
- Yamane, T.; Yoshida, N.; Sugioka, M. Estimation of Total Energy Requirement for Sewage Treatment by a Microbial Fuel Cell with a One-Meter Air-Cathode Assuming Michaelis–Menten COD Degradation. RSC Adv. 2021, 11, 20036–20045. [Google Scholar] [CrossRef]
- Chaudhuri, S.K.; Lovley, D.R. Electricity Generation by Direct Oxidation of Glucose in Mediatorless Microbial Fuel Cells. Nat. Biotechnol. 2003, 21, 1229–1232. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Liang, P.; Huang, X. Recent Progress in Electrodes for Microbial Fuel Cells. Bioresour. Technol. 2011, 102, 9335–9344. [Google Scholar] [CrossRef]
- Nagahashi, W.; Yoshida, N. Comparative Evaluation of Fibrous Artificial Carbons and Bamboo Charcoal in Terms of Recovery of Current from Sewage Wastewater. J. Gen. Appl. Microbiol. 2021, 67, 248–255. [Google Scholar] [CrossRef]
- Chen, S.; Liu, Q.; He, G.; Zhou, Y.; Hanif, M.; Peng, X.; Wang, S.; Hou, H. Reticulated Carbon Foam Derived from a Sponge-like Natural Product as a High-Performance Anode in Microbial Fuel Cells. J. Mater. Chem. 2012, 22, 18609–18613. [Google Scholar] [CrossRef]
- Chen, S.; He, G.; Hu, X.; Xie, M.; Wang, S.; Zeng, D.; Hou, H.; Schröder, U. A Three-Dimensionally Ordered Macroporous Carbon Derived from a Natural Resource as Anode for Microbial Bioelectrochemical Systems. ChemSusChem 2012, 5, 1059–1063. [Google Scholar] [CrossRef]
- Feng, Q.; Wang, Y.; Wang, T.; Zheng, H.; Chu, L.; Zhang, C.; Chen, H.; Kong, X.; Xing, X.H. Effects of Packing Rates of Cubic-Shaped Polyurethane Foam Carriers on the Microbial Community and the Removal of Organics and Nitrogen in Moving Bed Biofilm Reactors. Bioresour. Technol. 2012, 117, 201–207. [Google Scholar] [CrossRef]
- Chen, S.; He, G.; Liu, Q.; Harnisch, F.; Zhou, Y.; Chen, Y.; Hanif, M.; Wang, S.; Peng, X.; Hou, H.; et al. Layered Corrugated Electrode Macrostructures Boost Microbial Bioelectrocatalysis. Energy Environ. Sci. 2012, 5, 9769–9772. [Google Scholar] [CrossRef]
- Salas, E.C.; Sun, Z.; Lüttge, A.; Tour, J.M. Reduction of Graphene Oxide via Bacterial Respiration. ACS Nano 2010, 4, 4852–4856. [Google Scholar] [CrossRef]
- Yoshida, N.; Miyata, Y.; Doi, K.; Goto, Y.; Nagao, Y.; Tero, R.; Hiraishi, A. Graphene Oxide-Dependent Growth and Self-Aggregation into a Hydrogel Complex of Exoelectrogenic Bacteria. Sci. Rep. 2016, 6, 21867. [Google Scholar] [CrossRef]
- Yoshida, N.; Goto, Y.; Miyata, Y.; Thakur, V.K. Selective Growth of and Electricity Production by Marine Exoelectrogenic Bacteria in Self-Aggregated Hydrogel of Microbially Reduced Graphene Oxide. C 2016, 2016, 15. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Miyata, Y.; Mugita, A.; Iida, K. Electricity Recovery from Municipal Sewage Wastewater Using a Hydrogel Complex Composed of Microbially Reduced Graphene Oxide and Sludge. Materials 2016, 9, 742. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Miyata, Y.; Iida, K. Current Recovery from Sewage Wastewater Using Electrochemically Oxidized Graphite Felt. RSC Adv. 2019, 9, 39348–39354. [Google Scholar] [CrossRef] [Green Version]
- Goto, Y.; Yoshida, N. Preliminary Evaluation of a Microbial Fuel Cell Treating Artificial Dialysis Wastewater Using Graphene Oxide. AIP Conf. Proc. 2016, 1709, 020007. [Google Scholar] [CrossRef] [Green Version]
- Goto, Y.; Yoshida, N. Microbially Reduced Graphene Oxide Shows Efficient Electricity Ecovery from Artificial Dialysis Wastewater. J. Gen. Appl. Microbiol. 2017, 63, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Goto, Y.; Yoshida, N. Scaling up Microbial Fuel Cells for Treating. Water 2019, 11, 1803. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Xie, L.; Hirose, Y.; Nishiuchi, T.; Yoshida, N. Reduced Graphene Oxide Increases Cells with Enlarged Outer Membrane of Citrifermentans Bremense and Exopolysaccharides Secretion. Biosens. Bioelectron. 2022, 218, 114754. [Google Scholar] [CrossRef]
- Xie, L.; Yoshida, N.; Ishii, S.; Meng, L. Isolation and Polyphasic Characterization of Desulfuromonas Versatilis Sp. Nov., an Electrogenic Bacteria Capable of Versatile Metabolism Isolated from a Graphene Oxide-Reducing Enrichment Culture. Microorganisms 2021, 9, 1953. [Google Scholar] [CrossRef]
- Tanizawa, Y.; Okamoto, Y.; Tsuzuki, K.; Nagao, Y.; Yoshida, N.; Tero, R.; Iwasa, S.; Hiraishi, A.; Suda, Y.; Takikawa, H.; et al. Microorganism Mediated Synthesis of Reduced Graphene Oxide Films. In Journal of Physics: Conference Series, Proceedings of the Asia-Pacific Interdisciplinary Research Conference 2011 (AP-IRC 2011), Aichi, Japan, 17–18 November 2011; Institute of Physics Publishing: London, UK, 2012; Volume 352. [Google Scholar]
- Tomita, R.; Yoshida, N.; Meng, L. Formate: A Promising Electron Donor to Enhance Trichloroethene-to-Ethene Dechlorination in Dehalococcoides-Augmented Groundwater Ecosystems with Minimal Bacterial Growth. Chemosphere 2022, 307, 136080. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, N.; Asahi, K.; Sakakibara, Y.; Miyake, K.; Katayama, A. Isolation and Quantitative Detection of Tetrachloroethene (PCE)-Dechlorinating Bacteria in Unsaturated Subsurface Soils Contaminated with Chloroethenes. J. Biosci. Bioeng. 2007, 104, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Ismaeil, M.; Yoshida, N.; Katayama, A. Bacteroides Sedimenti Sp. Nov., Isolated from a Chloroethenes-Dechlorinating Consortium Enriched from River Sediment. J. Microbiol. 2018, 56, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Tamaoka, J.; Katayama-Fujimura, Y.; Kuraishi, H. Analysis of Bacterial Menaquinone Mixtures by High Performance Liquid Chromatography. J. Appl. Bacteriol. 1983, 54, 31–36. [Google Scholar] [CrossRef]
- Ismaeil, M.; Yoshida, N.; Katayama, A. Identification of Multiple Dehalogenase Genes Involved in Tetrachloroethene-to-Ethene Dechlorination in a Dehalococcoides -Dominated Enrichment Culture. Biomed. Res. Int. 2017, 2017, 9191086. [Google Scholar] [CrossRef] [Green Version]
- He, F.E. Coli Genomic DNA Extraction. Bio. Protoc. 2011, 1, 4–6. [Google Scholar] [CrossRef]
- Ohtsubo, Y.; Maruyama, F.; Mitsui, H.; Nagata, Y.; Tsuda, M. Complete Genome Sequence of Acidovorax Sp. Strain KKS102, a Polychlorinated-Biphenyl Degrader. J. Bacteriol. 2012, 194, 6970–6971. [Google Scholar] [CrossRef] [Green Version]
- Tanizawa, Y.; Fujisawa, T.; Nakamura, Y. DFAST: A Flexible Prokaryotic Genome Annotation Pipeline for Faster Genome Publication. Bioinformatics 2018, 34, 1037–1039. [Google Scholar] [CrossRef] [Green Version]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of Microbial Genomes Using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, 206–214. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M.; Spröer, C.; Klenk, H.P. When Should a DDH Experiment Be Mandatory in Microbial Taxonomy? Arch. Microbiol. 2013, 195, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a Taxonomic Coherence between Average Nucleotide Identity and 16S RRNA Gene Sequence Similarity for Species Demarcation of Prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Lovley, D.R.; Giovannoni, S.J.; White, D.C.; Champine, J.E.; Phillips, E.J.P.; Gorby, Y.A.; Goodwin, S. Geobacter Metallireducens Gen. Nov. Sp. Nov., a Microorganism Capable of Coupling the Complete Oxidation of Organic Compounds to the Reduction of Iron and Other Metals. Arch. Microbiol. 1993, 159, 336–344. [Google Scholar] [CrossRef]
- Waite, D.W.; Chuvochina, M.; Pelikan, C.; Parks, D.H.; Yilmaz, P.; Wagner, M.; Loy, A.; Naganuma, T.; Nakai, R.; Whitman, W.B.; et al. Proposal to Reclassify the Proteobacterial Classes Deltaproteobacteria and Oligoflexia, and the Phylum Thermodesulfobacteria into Four Phyla Reflecting Major Functional Capabilities. Int. J. Syst. Evol. Microbiol. 2020, 70, 5972–6016. [Google Scholar] [CrossRef] [PubMed]
- Shelobolina, E.S.; Nevin, K.P.; Blakeney-Hayward, J.D.; Johnsen, C.V.; Plaia, T.W.; Krader, P.; Woodard, T.; Holmes, D.E.; VanPraagh, C.G.; Lovley, D.R. Geobacter Pickeringii Sp. Nov., Geobacter Argillaceus Sp. Nov. and Pelosinus Fermentans Gen. Nov., Sp. Nov., Isolated from Subsurface Kaolin Lenses. Int. J. Syst. Evol. Microbiol. 2007, 57, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Prakash, O.; Gihring, T.M.; Dalton, D.D.; Chin, K.-J.; Green, S.J.; Akob, D.M.; Wanger, G.; Kostka, J.E. Geobacter Daltonii Sp. Nov., an Fe(III)- and Uranium(VI)-Reducing Bacterium Isolated from a Shallow Subsurface Exposed to Mixed Heavy Metal and Hydrocarbon Contamination. Int. J. Syst. Evol. Microbiol. 2010, 60, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Kunapuli, U.; Jahn, M.K.; Lueders, T.; Geyer, R.; Heipieper, H.J.; Meckenstock, R.U. Desulfitobacterium Aromaticivorans Sp. Nov. and Geobacter Toluenoxydans Sp. Nov., Iron-Reducing Bacteria Capable of Anaerobic Degradation of Monoaromatic Hydrocarbons. Int. J. Syst. Evol. Microbiol. 2010, 60, 686–695. [Google Scholar] [CrossRef]
- Sigrist, C.J.A.; Cerutti, L.; Hulo, N.; Gattiker, A.; Falquet, L.; Pagni, M.; Bairoch, A.; Bucher, P. PROSITE: A Documented Database Using Patterns and Profiles as Motif Descriptors. Brief Bioinform 2002, 3, 265–274. [Google Scholar] [CrossRef]
- Jiang, Y.; Shi, M.; Shi, L. Molecular Underpinnings for Microbial Extracellular Electron Transfer during Biogeochemical Cycling of Earth Elements. Sci. China Life Sci. 2019, 62, 1275–1286. [Google Scholar] [CrossRef]
- Levar, C.E.; Chan, C.H.; Mehta-Kolte, M.G.; Bond, D.R. An Inner Membrane Cytochrome Required Only for Reduction of High Redox Potential Extracellular Electron Acceptors. mBio 2014, 5, e02034-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levar, C.E.; Hoffman, C.L.; Dunshee, A.J.; Toner, B.M.; Bond, D.R. Redox Potential as a Master Variable Controlling Pathways of Metal Reduction by Geobacter Sulfurreducens. ISME J. 2017, 11, 741–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Fredrickson, J.K.; Zachara, J.M.; Shi, L. Direct Involvement of OmbB, OmaB, and OmcB Genes in Extracellular Reduction of Fe(III) by Geobacter Sulfurreducens PCA. Front. Microbiol. 2015, 6, 1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, Z.; Liu, J.; Levar, C.; Edwards, M.J.; Babauta, J.T.; Kennedy, D.W.; Shi, Z.; Beyenal, H.; Bond, D.R.; et al. A Trans-Outer Membrane Porin-Cytochrome Protein Complex for Extracellular Electron Transfer by Geobacter Sulfurreducens PCA. Environ. Microbiol. Rep. 2014, 6, 776–785. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Squier, T.C.; Zachara, J.M.; Fredrickson, J.K. Respiration of Metal (Hydr)Oxides by Shewanella and Geobacter: A Key Role for Multihaem c-Type Cytochromes. Mol. Microbiol. 2007, 65, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Aklujkar, M.; Coppi, M.V.; Leang, C.; Kim, B.C.; Chavan, M.A.; Perpetua, L.A.; Giloteaux, L.; Liu, A.; Holmes, D.E. Proteins Involved in Electron Transfer to Fe(III) and Mn(IV) Oxides by Geobacter Sulfurreducens and Geobacter Uraniireducens. Microbiology 2013, 159, 515–535. [Google Scholar] [CrossRef] [Green Version]
- Butler, J.E.; Young, N.D.; Lovley, D.R. Evolution of Electron Transfer out of the Cell: Comparative Genomics of Six Geobacter Genomes. BMC Genom. 2010, 11, 40. [Google Scholar] [CrossRef] [Green Version]
- Otero, F.J.; Chan, C.H.; Bond, D.R. Identification of Different Putative Outer Membrane Electron Conduits Necessary for Fe(III) Citrate, Fe(III) Oxide, Mn(IV) Oxide, or Electrode Reduction by Geobacter Sulfurreducens. J. Bacteriol. 2018, 200, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Zacharoff, L.A.; Morrone, D.J.; Bond, D.R.; Emerson, D.; Bond, D.R. Geobacter Sulfurreducens Extracellular Multiheme Cytochrome PgcA Facilitates Respiration to Fe (III) Oxides But Not Electrodes. Front. Microbiol. 2017, 8, 2481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campeciño, J.; Lagishetty, S.; Wawrzak, Z.; Alfaro, V.S.; Lehnert, N.; Reguera, G.; Hu, J.; Hegg, E.L. Cytochrome c Nitrite Reductase from the Bacterium Geobacter Lovleyi Represents a New NrfA Subclass. J. Biol. Chem. 2020, 295, 11455–11465. [Google Scholar] [CrossRef] [PubMed]
- Forest, K.T. Type IV Pili: Dynamics, Biophysics and Functional Consequences. Nat. Rev. Microbiol. 2019, 17, 429–440. [Google Scholar] [CrossRef]
- Lovley, D.R.; Walker, D.J.F.; Risgaard-petersen, N.; Lovley, D.R. Geobacter Protein Nanowires. Front. Microbiol. 2019, 10, 2078. [Google Scholar] [CrossRef] [Green Version]
- Walker, D.J.F.; Adhikari, R.Y.; Holmes, D.E.; Ward, J.E.; Woodard, T.L.; Nevin, K.P.; Lovley, D.R. Electrically Conductive Pili from Pilin Genes of Phylogenetically Diverse Microorganisms. ISME J. 2018, 12, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Noori, M.T.; Vu, M.T.; Ali, R.B.; Min, B. Recent advances in cathode materials and configurations for upgrading methane in bioelectrochemical systems integrated with anaerobic digestion. Chem. Eng. J. 2020, 392, 123689. [Google Scholar] [CrossRef]
- Shen, L.; Jin, Z.; Wang, D.; Wang, Y.; Lu, Y. Enhance wastewater biological treatment through the bacteria induced graphene oxide hydrogel. Chemosphere 2018, 190, 201–210. [Google Scholar] [CrossRef]
- Chen, X.; Chen, B. Macroscopic and spectroscopic investigations of the adsorption of nitroaromatic compounds on graphene oxide, reduced graphene oxide, and graphene nanosheets. Environ. Sci. Technol. 2015, 49, 6181–6189. [Google Scholar] [CrossRef]
- Lee, B.M.; Hur, J. Adsorption Behavior of Extracellular Polymeric Substances on Graphene Materials Explored by Fluorescence Spectroscopy and Two-Dimensional Fourier Transform Infrared Correlation Spectroscopy. Environ. Sci. Technol. 2016, 50, 6181–6189. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain NIT-SL11 | G. toluenoxydans TMJ1T | G. daltonii FRC-32T | G. uraniireducens Rf4T | |
|---|---|---|---|---|
| Size (µm) | 0.4 × 1.4 | 0.4 × 2.1–3.8 | 0.3–0.5 × 1.0–1.5 | 0.5–0.6 × 1.2–2.0 |
| Motility | − | − | ND | + |
| Optimum Temp. (°C) | 25–30 | 25–32 | 30 | 32 |
| Optimum pH | 6.0–.7 | 6.6–7.0 | 6.7–7.3 | 6.5–7.0 |
| NaCl (%) | 0–2 | ND | 0–0.7 | ND |
| e− donor | ||||
| Butanol | − | ND | + | − |
| Butyrate | − | + | + | − |
| Caproate | − | ND | ND | ND |
| Ethanol | − | ND | ND | + |
| Fructose | − | ND | ND | ND |
| Glucose | − | ND | ND | ND |
| Glycerol | − | ND | ND | ND |
| Isobutyrate | − | ND | ND | ND |
| Isopropanol | − | ND | ND | ND |
| Malate | − | − | ND | ND |
| Methanol | − | ND | ND | − |
| Phenol | − | + | ND | ND |
| Propionate | − | + | − | − |
| Succinate | − | − | − | − |
| Acetate | + | + | ND | + |
| Benzoate | + | + | + | ND |
| Formate | + | + | + | − |
| Lactate | + | − | − | + |
| Peptone | + | ND | ND | ND |
| Pyruvate | + | + | ND | + |
| YE | + | ND | ND | ND |
| H2 | ND | − | − | − |
| e− acceptors | ||||
| Nitrate | − | − | − | − |
| Sulfate | − | − | ND | − |
| Thiosulfate | − | − | ND | − |
| AQDS | − | ND | ND | + |
| S0 | − | − | + | − |
| Fumarate | + | + | + | + |
| Malate | + | ND | + | + |
| GO | + | ND | ND | ND |
| Electrode | + | ND | ND | ND |
| Fe(III) | ND | + | + | − |
| Mn(Ⅳ) | ND | − | ND | ND |
| Strain NIT-SL11 | G. toluenoxydans TMJ1T | G. daltonii FRC-32T | G. uraniireducens Rf4T | |
|---|---|---|---|---|
| Size (Mb) | 4.2 | 4.2 | 4.3 | 5.1 |
| GC (%) | 60 | 54 | 53 | 54 |
| Total predicted genes | 3923 | − | 3852 | 4591 |
| CDSs | 3860 | − | 3745 | 4457 |
| rRNA | 6 | − | 6 | 6 |
| tRNA | 56 | − | 49 | 49 |
| CytC | 80 | − | 73 | 69 |
| Menaquinone | 8, 9 | 8 | − | − |
| Fatty acids >40% | − | C16:0 | − | − |
| 30–40% | C16:1ω7c , C16:0 | − | C16:1ω7c , C16:0 | C16:1ω7c |
| 20–30% | − | C18:0 | − | C16:0 |
| 10–20% | − | − | ios-C15:0 | ios-C15:0 ,C14:0 |
| Tag | Local | CXnCH | 1 | 2 | 3 | ||
|---|---|---|---|---|---|---|---|
| n = 2 | n = 3 | n = 4 | |||||
| 01590 | − | 3 | 0 | 0 | 0105 | − | 4255 |
| 02110 | CM | 8 | 0 | 0 | cbcL,0274 | 3686 | 0125 |
| 03150 | CM | 6 | 1 | 0 | imcH,3259 | 1014 | 0861 |
| 03230 | − | 3 | 0 | 0 | ppcA,0612 | 1426 | 4121 |
| 03240 | PP | 3 | 0 | 0 | ppcC,0365 | 1426 | 4121 |
| 06040 | − | 3 | 0 | 0 | 0533 | 1401 | 3909 |
| 06440 | CP | 2 | 0 | 1 | 0591 | 1590 | 3656 |
| 06450 | OM | 12 | 0 | 0 | omcQ,0592 | 2170 | 3655 |
| 06470 | PP | 7 | 0 | 0 | cbcA,0594 | 3403 | 2712 |
| 06660 | PP | 3 | 0 | 0 | ppcA,0612 | 1426 | 4121 |
| 06690 | PP | 9 | 2 | 0 | 0615 | 1429 | 3840 |
| 06700 | OM | 8 | 0 | 0 | ctcB,0616 | 1430 | 3839 |
| 06720 | OM | 4 | 0 | 0 | omcE,0618 | 1685 | 3837 |
| 07200 | PP | 12 | 0 | 0 | omcX,0670 | 0830 | 0641 |
| 07560 | − | 5 | 0 | 0 | extCF,2725 | 1685 | 1838 |
| 08460 | − | 8 | 0 | 0 | omaB,2738 | 1681 | 0988 |
| 08470 | − | 9 | 0 | 0 | omcB,2737 | 1682 | 0989 |
| 08960 | PP | 2 | 0 | 0 | imcG,1538 | 1685 | 1316 |
| 09830 | − | 5 | 0 | 0 | extCF,2725 | 1685 | 1838 |
| 09840 | EX | 12 | 1 | 1 | extG,2724 | − | 1837 |
| 13150 | − | 1 | 0 | 0 | 1284 | 2256 | 2997 |
| 15110 | − | 27 | 0 | 0 | 2210 | 2170 | 3135 |
| 15170 | OM | 1 | 0 | 0 | 2204 | − | − |
| 15180 | PP | 10 | 0 | 0 | omcK,2203 | 1685 | 2291 |
| 15200 | − | 8 | 0 | 0 | cytT,2299 | 2175 | 0672 |
| 17180 | CM | 1 | 0 | 0 | coxB,0222 | − | 0414 |
| 17250 | EX | 4 | 0 | 0 | omcE,0618 | 1685 | 3837 |
| 17260 | EX | 8 | 0 | 0 | 2076 | − | − |
| 17360 | − | 6 | 0 | 0 | omcS,2503 | 3166 | 1989 |
| 17430 | PP | 26 | 0 | 0 | extQ,2495 | 3160 | 1995 |
| 17440 | PP | 16 | 0 | 0 | extR,2494 | 3159 | 1996 |
| 17570 | PP | 9 | 1 | 0 | omcI,1228 | 2912 | 2291 |
| 17990 | PP | 2 | 0 | 0 | macA,0466 | 2579 | 1316 |
| 18080 | CM | 8 | 0 | 0 | 2076 | − | − |
| 18110 | − | 13 | 0 | 1 | omcH,2884 | 0831 | 1834 |
| 18290 | CM | 7 | 0 | 0 | cbcA,0594 | 3403 | 2712 |
| 19400 | PP | 12 | 0 | 0 | omcV,1996 | 2379 | 2822 |
| 21440 | PP | 3 | 0 | 0 | ppcE,1760 | 1426 | 3843 |
| 21680 | PP | 1 | 0 | 0 | cycC,1740 | − | 2136 |
| 22490 | − | 5 | 0 | 0 | cbcX,1648 | − | 2381 |
| 22590 | EX | 2 | 0 | 0 | pgcA,1761 | 3176 | 2022 |
| 22840 | − | 3 | 1 | 0 | ctcD,1785 | − | − |
| 22850 | CP | 5 | 1 | 0 | 1786 | 2641 | 1922 |
| 22860 | − | 4 | 0 | 0 | 1787 | 2640 | 1921 |
| 25430 | IM | 1 | 0 | 0 | ccoP,2513 | − | 1913 |
| 25830 | − | 3 | 0 | 0 | 3214 | 1685 | 3748 |
| 26520 | EX | 8 | 0 | 0 | 2076 | − | − |
| 26970 | EX | 6 | 0 | 0 | omcM,2294 | − | − |
| 27550 | OM | 1 | 0 | 0 | omcF,2432 | − | 0331 |
| 28090 | EX | 14 | 0 | 3 | omcN,2898 | 3132 | 2035 |
| 28530 | OM | 8 | 0 | 0 | omcY,2201 | 2175 | 3130 |
| 28650 | EX | 10 | 0 | 0 | omcB,2737 | 1861 | 0989 |
| 28660 | − | 8 | 0 | 0 | omaB,2738 | 1681 | 0988 |
| 28700 | EX | 12 | 0 | 0 | omcB,2737 | 1682 | 0994 |
| 28710 | − | 8 | 0 | 0 | omaB,2738 | 1681 | 0988 |
| 28780 | PP | 1 | 0 | 0 | petJ,2743 | − | 0331 |
| 29020 | OM | 14 | 0 | 3 | omcN,2898 | 3132 | 2035 |
| 30810 | − | 5 | 0 | 0 | extD,2642 | − | − |
| 30820 | − | 4 | 0 | 1 | extCF,2643 | 1681 | 1838 |
| 30840 | PP | 11 | 1 | 0 | extA,2645 | 1685 | 0641 |
| 31410 | OM | 5 | 0 | 0 | ctcC,2801 | 0309 | − |
| 32150 | − | 20 | 0 | 4 | omcG,2882 | 3132 | 3427 |
| 32160 | − | 19 | 0 | 4 | omcH,2883 | 3132 | 3427 |
| 32170 | − | 24 | 0 | 3 | omcH,2884 | 1685 | 3428 |
| 32200 | − | 21 | 0 | 5 | omcG,0702 | 1685 | 2035 |
| 32390 | OM | 23 | 0 | 3 | omcN,2898 | 1513 | 3428 |
| 32470 | EX | 20 | 0 | 3 | omcN,2898 | 1513 | 2035 |
| 32480 | EX | 15 | 0 | 8 | omcM,2899 | 1685 | 0076 |
| 32490 | − | 20 | 0 | 7 | omcG,0702 | 1513 | 3427 |
| 32550 | EX | 23 | 1 | 3 | omcH,2912 | 1685 | 3428 |
| 32560 | EX | 4 | 0 | 0 | omcP,2913 | 1432 | 3837 |
| 32700 | − | 2 | 0 | 0 | 2927 | 1359 | 4833 |
| 32730 | − | 4 | 0 | 0 | cbcR,2930 | 1348 | 0460 |
| 32770 | CP | 10 | 0 | 0 | cbcN,2934 | 1344 | 0456 |
| 32780 | EX | 12 | 0 | 0 | cbcM,2935 | 1343 | − |
| 32800 | PP | 5 | 0 | 0 | extK,2937 | 1340 | 0447 |
| 34580 | PP | 9 | 0 | 0 | 3137 | 1724 | − |
| 34890 | − | 2 | 0 | 0 | 2767 | 1685 | 4149 |
| 35130 | PP | 4 | 0 | 0 | nrfA,3154 | 3111 | 0665 |
| 37130 | − | 2 | 0 | 0 | 3332 | 3818 | 4408 |
 | |||||||
| Tag | Annotation | Aromatic Acid Mole % | 1 | 2 | 3 | |||
|---|---|---|---|---|---|---|---|---|
| 20580 | geopilin domain 1 protein pilA-N | 8.5 | GSU1496 | Geob3369 | Gura2677 | |||
| 20590 | geopilin domain 2 protein pilA-C | 8.8 | GSU1497 | − | − | |||
| 33430 | PilZ domain protein | 11.0 | GSU3028 | Geo0640 | Gura4082 | |||
| 28450 | PilZ domain protein | 9.7 | GSU1051 | Geob0928 | Gura0726 | |||
| 17690 | PilZ domain protein | 10.7 | GSU1240 | Geob0815 | Gura0044 | |||
| 36280 | PilZ domain protein | 8.6 | GSU0137 | − | Gura3986 | |||
| 01060 | PilZ domain protein | 8.2 | GSU0078 | − | − | |||
| 37060 | PilZ domain protein | 10.6 | GSU0312 | Geob1410 | Gura3298 | |||
| 20560 | sensor histidine kinase PilS | 11.0 | − | − | − | |||
| 20570 | sigma-54-dependent transcriptional response pilR | 8.3 | − | − | − | |||
| 36190 | twitching motility pilus retraction protein pilT-1 | 8.6 | − | − | − | |||
| 01730 | twitching motility pilus retraction protein pilT-3 | 8.5 | − | − | − | |||
| 20540 | twitching motility pilus retraction protein pilT-4 | 7.4 | − | − | − | |||
| 26020 | type IV pilus assembly lipoprotein PilP | 5.0 | GSU2029 | Geob3067 | Gura1813 | |||
| 26080 | type IV pilus assembly protein PilY1 | 10.2 | GSU1066 | Geob3067 | − | |||
| 20530 | type IV pilus biogenesis ATPase PilB | 6.7 | − | − | − | |||
| 26050 | type IV pilus biogenesis ATPase PilM | 6.8 | − | − | − | |||
| 26040 | type IV pilus biogenesis ATPase PilM | 4.2 | GSU3069 | Geob3069 | Gura1811 | |||
| 26030 | type IV pilus biogenesis protein PilO | 0.0 | GSU2030 | Geob3068 | Gura1812 | |||
| 20550 | type IV pilus inner membrane protein PilC | 7.9 | − | − | − | |||
| 26130 | type IV pilus minor pilin PilE | 10.2 | GSU3548 | − | − | |||
| 26010 | type IV pilus secretin lipoprotein PilQ | 5.8 | − | − | − | |||
| 26160 | type IV prepilin-like proteins leader peptide pilD | 15.1 | GSU2043 | Geob3081 | Gura1794 | |||
| Identity (%) | 20–30 | 31–40 | 41–50 | 51–60 | 61–70 | 71–80 | 81–90 | 91–100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, L.; Yoshida, N.; Meng, L. Polyphasic Characterization of Geotalea uranireducens NIT-SL11 Newly Isolated from a Complex of Sewage Sludge and Microbially Reduced Graphene Oxide. Microorganisms 2023, 11, 349. https://doi.org/10.3390/microorganisms11020349
Xie L, Yoshida N, Meng L. Polyphasic Characterization of Geotalea uranireducens NIT-SL11 Newly Isolated from a Complex of Sewage Sludge and Microbially Reduced Graphene Oxide. Microorganisms. 2023; 11(2):349. https://doi.org/10.3390/microorganisms11020349
Chicago/Turabian StyleXie, Li, Naoko Yoshida, and Lingyu Meng. 2023. "Polyphasic Characterization of Geotalea uranireducens NIT-SL11 Newly Isolated from a Complex of Sewage Sludge and Microbially Reduced Graphene Oxide" Microorganisms 11, no. 2: 349. https://doi.org/10.3390/microorganisms11020349